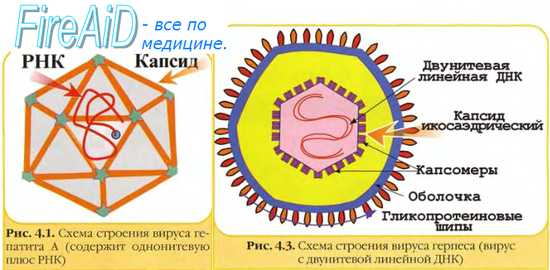
Оглавление темы "Методы обнаружения вирусов. Методы диагностики микозов ( грибковых заболеваний ). Методы обнаружения простейших.": 1. Биологические методы диагностики бактерий. Животные при диагностике инфекций. Какие животные используются для диагностики инфекций?. 2. Методы обнаружения вирусов. Лабораторные методы при диагностике вирусных инфекций. Забор материала для выявления вирусов. Культуры клеток для выявления вирусов. 3. Культуры органов для обнаружения вирусов. Куриные эмбрионы при диагностике вирусных инфекций. Заражение вирусом куриного эмбриона. Методы заражения вирусом куриного эмбриона. 4. Животные модели для обнаружения вирусов. Идентификация вирусов. Качественное определение вирусов. Цитопатические эффекты вирусов. Бляшкообразование вируса. Тельца включений вирусов. 5. Отсутствие цитопатического эффекта вируса. Феномен гемадсорбции вирусов. Цветная реакция. Экспресс-диагностика вирусной инфекции. 6. Количественное определение вирусов. Определение инфекционности вирусов. Выявление вирусных антигенов ( Аг ). Выявление вирусных частиц. Морфология вирусов. 7. Серологические методы диагностики вирусных инфекций. Торможение гемагглютинации. Торможение цитопатического эффекта интерференцией вирусов. Прямая иммунофлюоресценция. Иммуноэлектронная микроскопия. 8. Выявление противовирусных антител ( AT ) в сыворотке крови. РТГА. РСК. РИФ. Иммуносорбционные методы выявления противовирусных антител. 9. Выявление вирусных антигенов ( Аг ). ИФА. Гибридизация ДНК. ПЦР. Методы диагностики микозов ( грибковых заболеваний ). 10. Выделение грибов. Неселективные среды для грибов. Селективные среды для грибов. Выявление противогрибковых антител ( AT ). Выявление грибковых антигенов ( Аг ). 11. Методы обнаружения простейших. Микроскопия простейших. Материал для выявления простейших. Выделение простейших. Серологические исследования при диагностике простейших.
Методы обнаружения вирусов. Лабораторные методы при диагностике вирусных инфекций. Забор материала для выявления вирусов. Культуры клеток для выявления вирусов.
Лабораторные методы при диагностике вирусных инфекций включают: • выделение и идентификацию возбудителя; • обнаружение и определение титров противовирусных AT; • обнаружение Аг вирусов в образцах исследуемого материала; • микроскопическое исследование препаратов исследуемого материала.
При заборе материала для исследований необходимо выполнять следующие условия: • образцы следует отбирать как можно раньше либо с учётом ритма циркуляции возбудителя; • материал следует отбирать в объёме, достаточном для всего комплекса исследований; • образцы следует доставлять в лабораторию незамедлительно (!), при относительно кратковременной транспортировке (не более 5 сут) образцы сохраняют на льду, при более длительной — при температуре -50 С.
Выделение и идентификация возбудителя — золотой стандарт в диагностике вирусных инфекций.
Вирусы размножаются только в живых клетках, и выделение возбудителя в заражённой культуре клеток — один из основных методов диагностики вирусных инфекций. Поскольку большинство патогенных вирусов отличает тканевая и типовая специфичность, то почти к каждому вирусу можно подобрать соответствующие клеточные или тканевые чультуры, а также создать стандартные условия культивирования (наличие клеток одного типа). Размножение вируса обеспечивают чувствительные (пермиссивные) клетки. Поэтому при выделении неизвестного возбудителя проводят одномоментное заражение 3
4 культур клеток, предполагая, что одна из них может оказаться пермиссивной. Культуры клеток получают диспергированием соответствующих органов и тканей, но чаще используют эмбриональные ткани (человека и животных) либо трансформированные опухолевые клетки. При помещении на соответствующую плоскую поверхность клеточные культуры обычно растут в виде монослоя. Первично-трипсинизированные культуры. Суспензии клеток получают гомогенизированием соответствующих тканей, предварительно обработанных трипсином. Культуры часто представлены клетками смешанного типа и не подлежат повторному культивированию. Жизнеспособность таких культур составляет 2-3 нед.
Полуперевиваемые линии клеток представлены диплоидными клетками человека и животных. Культуры ограниченно пригодны к повторному диспергированию и росту (как правило, не более 20-30 пересевов), сохраняя при этом жизнеспособность и не подвергаясь спонтанной трансформации.
Перевиваемые линии клеток (гетероплоидные культуры) представлены клетками, подвергнутыми длительному культивированию и спонтанным трансформациям. Культуры способны к многократному диспергированию и перевиванию. Работа с ними менее трудоёмка по сравнению с приготовлениями первичных культур; перевиваемые клетки относительно одинаковы по своей морфологии и стабильны по свойствам.
2) Постановка РГА (реакции гемадсорбции) – к/к, зараженная вирусами, имеющими гемагглютинины (вирус гриппа и др.), приобретает способность адсорбировать Эр на своей поверхности.
4) Постановка цветной пробы.
(индикация вирусов после культивирования на курином эмбрионе и животныхпроводится с помощью РГА).
Идентификация вирусов в к/к
Проводится РН (реакция нейтрализации) с диагностическими сыворотками. Результат реакции оценивается по наличию ЦПД, методом бляшек, постановкой РТГА, цветной пробы.
(идентификация вирусов после культивирования на курином эмбрионе и животных проводится с помощью РН).
Методы диагностики вирусных инфекций
I. Вирусологический метод диагностики.
Конечной целью метода является идентификация вирусов до вида или серологического варианта. Вирусологический метод включает несколько этапов:
Отбор материала для исследования.
В зависимости от характера заболевания, материалом для исследования могут быть: 1) смывы с носовой части глотки и мазок из глотки; 2) спинномозговая жидкость; 3) кал и ректальные мазки; 4) кровь; 5) моча; 6) жидкость из серозных полостей; 7) мазок с конъюнктивы; 8) содержимое везикул; 8) секционный материал.
• материал берут непосредственно из очага инфекции или исследуют соответствующее отделяемое (гной, мочу, желчь и т.п.);
• количество материала должно быть достаточным для проведения исследования и его повторения в случае необходимости;
• материал берут по возможности в начальном периоде болезни, так как именно в этот период возбудители выделяются чаще, их больше, они имеют более типичную локализацию;
• материал берут до начала антимикробной химиотерапии или через определенный промежуток времени после приема антибактериального препарата, необходимый для его выведения из организма;
• следует предупредить возможность попадания в материал антимикробных препаратов (дезинфектанты, антисептики, антибиотики);
2. Обработка вируссодержащего материала. Проводится с целью освобождения материала от сопутствующей бактериальной микрофлоры. Для этого используются физические и химические методы. Физические методы: 1) фильтрование через различные бактериальные фильтры; 2) центрифугирование. Химические методы: 1) обработка материала эфиром в случаях выделения вирусов, не имеющих суперкапсида; 2) добавление к материалу смеси гептана и фреона; 3) внесение антибиотиков
3. Заражение материалом чувствительных живых систем. Поскольку вирусы являются облигатными внутриклеточными паразитами, для их размножения используют следующие живые системы: 1) лабораторные животные; 2) куриные эмбрионы; 3) культуры органов; 4) культуры тканей.
Индикация вирусов в живых системах.
Индикация вирусов – это обнаружение вирусов в исследуемом материале без установления их принадлежности к семейству, роду, виду или сероварианту.
5. Титрование выделенных вирусов -это обязательный этап вирусологического метода диагностики, целью которого является количественное определение содержания вирусных частиц в единице объема исследуемого материала.
6. Идентификация вирусов в иммунных реакциях. Идентификация, или титрование вирусов – это установление их вариантной, видовой, родовой и семейственной принадлежности. Идентификация вирусов проводится по принципу: определение неизвестного по известному. Известным компонентом при идентификации вирусов являются специфические противовирусные сыворотки (противогриппозные, противокоревые и др.), которые используют в серологических реакциях нейтрализации (РН), торможения гемадсорбции (РТГадс), торможения гемагглютинации (РТГА), РПГА, РСК, а также при ИФА и РИА. Эти сыворотки содержат специфические противовирусные антитела и называются диагностическими.
II. Вирусоскопический метод
1) Электронная микроскопия
Цель: детальное изучение строения вируса.
2) Световая микроскопия
Цель: а) обнаружения вирионов размером от 200 нм (серебрение по Морозову –
выявление телец Пашена – вирус натуральной оспы).
б) обнаружение включений:
- в цитоплазме (тельца Бабеша-Негри при бешенстве (окраска по Муромцеву), тельца Гварниери при натуральной оспе);
Культивирование вирусов в организме лабораторных животных. Выбор экспериментальных животных определяется целью работы и видовой чувствительностью к изучаемому вирусу. Для заражения используют обезьян, кроликов, морских свинок, хомячков, белых крыс и мышей.
Лабораторных животных заражают различными способами в зависимости от тропизма вируса к определенным тканям. Так, например, для культивирования нейротропных вирусов заражение производят преимущественно в мозг (вирусы бешенства, клещевого энцефалита и др.), культивирование респираторных вирусов осуществляется при интраназальном инфицировании животных (вирусы гриппа), дерматотропных (вирус оспы) – путем накожного и внутрикожного заражения. Наиболее часто используются накожное, внутрикожное, внутримышечное, внутрибрюшинное и внутримозговое заражение.
Индикацию, т.е. обнаружение факта размножения вируса, устанавливают на основании развития типичных признаков заболевания, патоморфологических изменений органов и тканей животных или положительной реакции гемагглютинации (РГА). РГА основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов различных видов животных, птиц и человека за счет поверхностного вирусного белка – гемагглютинина.
В настоящее время использование животных для культивирования вирусов ограничено.
Культивирование вирусов в куриных эмбрионах. Большинство известных вирусов обладают способностью размножаться в курином эмбрионе (рис.56). Используют эмбрионы в возрасте от 8 до 14 дней в зависимости от вида вируса, способа заражения и задач исследования. Вирусы гриппа культивируются в 9–10-, осповакцины – в 12-, паротита – в 7-дневных куриных эмбрионах. Размножение вируса в куриных эмбрионах происходит в разных частях зародыша, что связано с особенностями тропизма вируса. Методику выращивания вируса в курином эмбрионе широко используют при промышленном культивировании.
Рис. 56. Строение куриного эмбриона и способы его заражения: 1 – в амнион; 2 – в аллантоисную полость; 3 – в желточный мешок. (Микробиология и иммунология. Под редакцией Воробьева А.А. – М. – 1999).
Существует несколько способов заражения развивающегося куриного эмбриона: на хорионаллантоисную оболочку, в аллантоисную и амниотическую полости, желточный мешок, тело эмбриона.
Заражение на хорионаллантоисную оболочку применяется для выделения и культивирования вирусов, образующих на оболочках бляшки (вирусы вакцины, натуральной оспы, простого герпеса). Перед заражением яйца просвечивают с помощью овоскопа, карандашом очерчивают границу воздушного пространства и хорионаллантоисной оболочки. Поверхность яйца над воздушным пространством и в месте заражения протирают спиртом, прожигают, обрабатывают йодом и делают отверстие в полости воздушного мешка. На месте заражения скорлупу удаляют так, чтобы не повредить подскорлупную оболочку, которую затем прокалывают короткой стерильной иглой, чтобы не повредить хорионаллантоисную оболочку. Воздух из полости воздушного мешка отсасывают. Вирусный материал (0,05–0,2 мл) наносят на хорионаллантоисную оболочку туберкулиновым шприцем с короткой иглой или пастеровской пипеткой. Отверстие в скорлупе закрывают стерильным покровным стеклом или тем же выпиленным кусочком скорлупы и по краям заливают расплавленным парафином. Зараженные эмбрионы располагают на подставке горизонтально и инкубируют в термостате. Вскрытие эмбрионов производится не раньше 48 ч инкубации. На зараженной оболочке обнаруживаются беловатые непрозрачные пятна разной формы (бляшки).
Заражение в аллантоисную полость. Вирус, введенный в аллантоис, размножается в эндодермальных клетках, переходя затем в аллантоисную жидкость. Заражение осуществляют следующим способом: в скорлупе над воздушной камерой острием скальпеля или ножниц производят прокол, после чего через отверстие в вертикальном направлении вводят иглу со шприцем, которая проходит через хорионаллантоисную оболочку и попадает в аллантоисную полость, материал вводится в объеме 0,1 мл и отверстие заливают парафином.
Заражение в желточный мешок. С этой целью используют эмбрионы 5–10-дневного возраста. Наиболее употребительны два метода заражения. По первому материал вводится через воздушное пространство. В центре яйца делают отверстие, помещают его на подставку тупым концом вправо и через отверстие в вертикальном направлении вводят иглу, надетую на шприц, игла проходит через хорионаллантоисную оболочку, аллантоисную полость в желток. В желточный мешок можно ввести от 0,1 до 0,5 мл вируссодержащего материала. После заражения отверстие в скорлупе заливают парафином, и эмбрион помещают в термостат. По второму методу на границе воздушного пространства с той стороны, где лежит желток (стороны, противоположной от эмбриона), делают прокол скорлупы, через который вводят инфекционный материал. Направление иглы должно быть к центру яйца.
Индикацию вирусов в курином эмбрионе осуществляют на основании специфических поражений оболочек и тела эмбриона (оспины, кровоизлияния), а также в РГА.
1. однослойные – клетки, способные прикрепляться и размножаться на поверхности химически нейтрального стекла лабораторной посуды в виде монослоя;
2. суспензионные – клетки, размножающиеся во всем объеме питательной среды при постоянном ее перемешивании;
3. органные – цельные кусочки органов и тканей, сохраняющие исходную структуру вне организма (применяются ограничено).
По числу жизнеспособных генераций культуры клеток подразделяются на:
1. первичные, способные размножаться только на первых генерациях, т.е. в нескольких пассажах после выделения из тканей;
2. перевиваемые, или стабильные, способные размножаться в лабораторных условиях неопределенно длительный срок посредством постоянного пассирования;
3. полуперевиваемые, имеющие ограниченную продолжительность жизни (40-50 пассажей).
Приготовление первичной культуры клеток складывается из нескольких последовательных этапов: измельчение ткани, разъединение клеток путем трипсинизации, отмывание полученной однородной суспензии изолированных клеток от трипсина с последующем суспендированием клеток в питательной среде.
Перевиваемые однослойные культуры клеток приготавливают из злокачественных или нормальных линий клеток, обладающих способностью длительно размножаться in vitro в определенных условиях. К ним относятся злокачественные клетки HeLa, первоначально выделенные из карциномы шейки матки, Hep-3 (из лимфоидной карциномы), а также нормальные клетки амниона человека, почек обезьян и др.
Рис. 57. Индикация репродукции вируса в культуре ткани по цитопатическому действию (ЦПД): а – интактная монослойная культура клеток, б – зараженная культура (ЦПД). (Микробиология и иммунология.-Под ред. А.А. Воробьева.-М, Медицина, 1999.-464 с.)
К полуперевиваемым культурам относятся диплоидные клетки человека. Они представляют собой клеточную систему, сохраняющую в процессе 50 пассажей (до года) диплоидный набор хромосом. Диплоидные клетки человека не претерпевают злокачественного перерождения и этим выгодно отличаются от опухолевых.
Для выращивания вирусов можно использовать культуры тканей любого типа. Доза заражения зависит от цели и назначения опыта. Тканевые культуры используют для выделения новых малоизученных вирусов, когда обычным методом (заражение животных, куриных эмбрионов) невозможно установить вирусную природу возбудителя. Выбор клеточных культур определяется их чувствительностью к отдельным группам вирусов.
Различают острую и хроническую инфекции. Острое течение инфекции характеризуется цитопатическим действием (деструктивными изменениями зараженных клеток, завершающихся их гибелью). Хроническая форма репродукции вируса не вызывает быструю гибель клеток, они долгое время остаются жизнеспособными и внешне могут не отличаться от зараженных.
Индикацию вирусов в культуре клеток проводят на основании следующих феноменов:
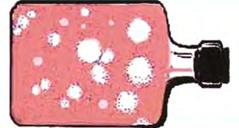
1. Цитопатическое действие (ЦПД) – видимые под микроскопом морфологические изменения клеток, вплоть до их отторжения от стекла, которые возникают в результате внутриклеточной репродукции вирусов (рис. 57). Характер ЦПД при различных вирусных инфекциях неодинаков. При репродукции одних вирусов (парамиксовирусы, герпесвирусы) наблюдается слияние клеток с образованием синцития, других (энтеровирусы, реовирусы) – сморщивание и деструкция клеток, третьих (аденовирусы) – агрегация клеток и т.д.
2. Вирусные включения – скопление вирусных частиц или отдельных компонентов вирусов в цитоплазме или ядре клеток, выявляемые под микроскопом при специальном окрашивании. Включения различаются по величине, форме, численности. Характерные ядерные включения формируются в клетках, зараженных вирусами герпеса, аденовирусами, гриппа, бешенства, оспы и др.
3. Бляшки, или негативные колонии – ограниченные участки, состоящие из дегенеративных клеток, которые вирусы способны образовывать в монослое клеток под агаровым покрытием. Они видны невооруженным глазом как светлые пятна на фоне прижизненно окрашенных нейтральным красным клеток. Одна бляшка соответствует потомству одного вириона. Негативные колонии разных вирусов отличаются по размеру, форме. Бляшкообразование используют для дифференциации, селекции вирусов, а также для определения их концентрации в исследуемом материале. Титр вируса, установленный этим методом, выражают числом бляшкообразующих единиц (БОЕ) в 1 мл.
5. Гемадсорбция – способность культур клеток, инфицированных вирусами, адсорбировать на своей поверхности эритроциты определенных видов животных и птиц. Гемадсорбция проявляется скоплением в виде гроздей эритроцитов, адсорбированных на инфицированных вирусом клетках.
Читайте также:
Cтруктуры внешней памяти, методы организации индексов
GIAI для идентификации инвентарного имущества
II. Методы искусственной детоксикации организма
II. Методы несанкционированного доступа.
III. Методы искусственной физико-химической детоксикации.
III. Методы манипуляции.
IV. Традиционные методы среднего и краткосрочного финансирования.
IX. Методы СТИС
R Терапевтическая доза лазерного излучения и методы ее определения
VΙ.Организация и методы выполнения процесса
При заражении вирусами клеточных культур можно получать различные видимые проявления действия вируса:
· Цитопатическое действие вируса на культуру клеток (ЦПД) – возникновение в ней видимых морфологических дегенеративных изменений (литическая инфекция);
· Приобретение заражённой культурой клеток способностик гемадсорбции – к адсорбции эритроцитов на поверхности клеточного слоя;
· Подавление процессов метаболизма в заражённой вирусом культуре клеток, выявляемое с помощью так называемой цветной пробы.
Окончательная идентификация выделенного вируса проводится с помощью реакции нейтрализации с диагностическими вируснейтрализующими сыворотками. Определяют их родовую и видовую принадлежность.
2.1.1. Цитопатическое действие (ЦПД) вируса в культуре клеток
Основной причиной ЦПД является нарушение метаболизма клетки. Прекращается синтез РНК клетки-хозяина, что ведёт к подавлению синтеза белков, приводит к нарушению структуры клеточных мембран, лизосом, митохондрий. Освобождаются и активируются клеточные ферменты (лизосомальные), которые вызывают деструкцию клеточных компонентов, т.е. развитие ЦПД. При резкой дегенерации клеточный монослой гибнет.
При острой вирусной инфекции может наблюдаться образование гигантских многоядерных клеток – симпластов (или синцитиев). Симпластообразование вызывают более 20 различных вирусов, имеющих ферменты: лецитиназу, нейраминидазу, и богатых липидами (напр., парамиксовирусы).
Рис. 5. Проявление ЦПД в культуре клеток
К проявлению ЦПД вирусов относится образование внутриклеточных включений. Они образуются, если вирус не вызывает гибели клеток, или на стадиях до наступления гибели. Образование включений может быть единственным проявлением реакции клетки на внедрение вируса.
С целью обнаружения вируса в материалах от больных проводят заражение исследуемым материалом однослойных культур клеток. Для заражения отбирают пробирки со сплошным клеточным слоем, просматривая их под малым увеличением микроскопа. Перед заражением из пробирок отсасывают культуральную жидкость, затем вносят по 0,1 мл исследуемого материала и добавляют питательную среду до 1 мл. Каждую пробу материала вносят в 4 пробирки. Для контроля несколько пробирок оставляют незаражёнными, но также сменяют в них питательную среду. Пробирки выдерживают в термостате, обычно при 37 0 С, и ежедневно микроскопируют с целью обнаружения ЦПД (в течение недели и более).
ЦПД различных вирусов на клеточные культуры обладает определённой специфичностью. Родственные вирусы дают цитопатическую реакцию сходного типа, эффект действия отдалённых по свойствам вирусов часто различен, поэтому по типу ЦПД можно судить о семействе или роде, к которым относится исследуемый вирус:
- аденовирусы превращают клеточный слой в скопления мелких, округлых клеток, расположенных в виде гроздьев винограда;
- парагриппозные вирусы, респираторно-синцитиальный, вирусы кори и паротита образуют симпласты.
Количественное определение вирусов в исследуемом материале проводится с помощью титрования. Для этой цели готовят 10-кратные последовательные разведения вируса, которыми заражают клеточные культуры. Титром вируса называют его наибольшее разведение, вызывающее ЦПД в половине заражённых культур. Титр вируса выражают в цитопатических дозах (ЦПД50) в 1 мл. За одну ЦПД50 принимают 0,1 мл вируссодержащего материала, разведённого до титра.
Для идентификации выделенного вируса по нейтрализации ЦПД культуральную жидкость смешивают с равным объёмом диагностической иммунной сыворотки (разведение сыворотки 1:5 или 1:10). После 1-2-часового контакта при комнатной температуре этой смесью (по 0,2 мл) заражают 4 пробирки с культурой клеток, из которой предварительно была удалена питательная среда; после добавления смеси в пробирки вносят по 0,8 мл свежей среды. Опыт сопровождается несколькими контролями:
1 – контроль незаражённой культуры;
2 – контроль дозы вируса – клеточные культуры заражают той же дозой вируса, что и в опыте;
3 – контроль культуры, заражённой смесью культуральной жидкости с нормально сывороткой.
Опыт учитывают через 5-7 дней и более, просматривая пробирки под малым увеличением микроскопа. В первом контроле ЦПД должно отсутствовать, во втором и третьем – обязательное проявление ЦПД. Отсутствие цитопатического эффекта в опытных пробирках указывает, что в данной пробе произошла нейтрализация вируса иммунной сывороткой, сыворотка соответствует типу выделенного вируса.
2.1.2. Реакция гемадсорбции
Гемадсорбирующие свойства имеют многие вирусы: орто- и парамиксовирусы, флавивирусы, поксвирусы. Эти же вирусы обладают спосодностью вызывать гемагглютинацию – склеивание эритроцитов. Приобретение способности к гемадсорбции связано со встраиванием в мембрану заражённых вирусом клеток вирусспецифических белков – гемагглютининов, к которым эритроциты имеют комплементарные рецепторы и поэтому адсорбируются на поверхности заражённых клеток. В реакции применяют эритроциты морской свинки, кур, обезьян, человека 0 (I) группы. По наличию явления гемадсорбции обнаруживают присутствие вирусов в культурах клеток при латентной инфекции, когда цитопатический эффект оказывается слабо выраженным или отсутствует совершенно.
Рис. 6. Реакция гемадсорбции
Техника постановки реакции гемадсорбции
Предварительно клеточные культуры заражают исследуемым материалом. Реакцию гемадсорбции ставят каждые два дня до появления положительной реакции (в течение 8-10 дней). Для этого в пробирку с заражённой культурой вносят по 0,2 мл взвеси эритроцитов (0,4-1%) и выдерживают пробирки в наклонном положении, чтобы эритроциты соприкасались с клеточным слоем. Температура, при которой ставят опыт, и экспозиция зависит от вида вируса. Пробирки встряхивают и оставляют на некоторое время в вертикальном положении для оседания неадсорбировавшихся эритроцитов. При микроскопии под малом увеличением в опытной пробирке регистрируют положительную реакцию гемадсорбции, выражающуюся в том, что эритроциты адсорбируются на клетках культуры, прилипая к клеточному слою; в контрольной пробирке клеточный слой свободен от эритроцитов.
При добавлении специфической иммунной сыворотки в пробирку с клеточной культурой, предварительно заражённой соответствующим вирусом, заражённые клетки теряют способность адсорбировать эритроциты, т.е. происходит явление задержки гемадсорбции. Вследствие специфичности этого явления реакция задержки гемадсорбции может быть использована для идентификации выделенных вирусов, а также с целью обнаружения и титрования вируснейтрализующих антител в сыворотках больных.
2.1.3.Метод бляшек
Для получения бляшек клеточный монослой заражают небольшой концентрацией вируса и фиксируют адсорбировавшиеся на клетках вирионы с помощью агарового покрытия, в которое добавлен витальный краситель – нейтральный красный. ЦПД вирусов имеет очаговый характер, погибшие клетки дегенерируют, теряют способность удерживать нейтральный красный и обесцвечиваются. В результате на непрозрачном равномерно розовом фоне клеточного монослоя появляются бляшки в виде более прозрачных неокрашенных округлых пятен.
Рис. 7. Метод бляшек
Агаровое покрытие готовят из высококачественного агара в концентрации 1-1,5% и других компонентов – солевых буферных растворов, дополнительных питательных веществ, антибиотиков. Раствор нейтрального красного входит в состав среды или его добавляют во флакон или чашку незадолго до учёта опыта. Часто вместо агара используют гель бентионита (5-6%) – это алюмосиликат природного происхождения, который биологически инертен и не токсичен для культур клеток.
Обнаружение и титрование вирусов методом бляшек
Для получения бляшек вирусов во флаконы или чашки наливают взвесь клеток в питательной среде (№ 199 или среда с гидролизатом лактальбумина). Флаконы закрывают резиновыми пробками, чашки заклеивают лейкопластырем и помещают в термостат на 5-6 дней для получения монослоя клеток, который должен быть сплошным, без признаков дегенерации. Перед заражением среду из флаконов или чашек удаляют и однослойную культуру клеток осторожно отмывают раствором Хенкса, который затем тщательно отсасывают. Заражение производят путём внесения разведённого исследуемого материала в объёме 0,1-0,25 мл и равномерного распределения этого материала по поверхности клеточного слоя с помощью покачивания. Через 30-60 минут заражённую культуру клеток заливают покровной средой. Флаконы или чашеи с застывшей средой помещают в термостат (клеточным слоем кверху) и выдерживают до образования бляшек (2-5 суток), после чего производят их подсчёт и изучение.
Разные вирусы образуют бляшки, отличающиеся по величине, форме, характеру краёв, по срокам появления и другим свойствам, что может быть использовано для предварительной идентификации вируса.
Для титрования вирусов готовят серийные разведения вируссодержащего материала и каждое из разведений вносят во флакон или чашку со слоем культуры клеток. Подсчитав образовавшиеся бляшки (с учётом разведения вируса), вычисляют титр вируса – количество вирионов в 1,0 мл исходного материала. Титр вируса, определяемый методом бляшек, принято выражать числом бляшкообразующих единиц (БОЕ) в 1 мл.
Метод бляшек даёт возможность провести точную идентификацию вируса с помощью диагностических специфических сывороток. При соответствии между сывороткой и вирусом происходит нейтрализация вируса и при заражении монослоя клеток такой смесью бляшки не образуются или их количество значительно снижается.
2.1.4. Цветная проба
Цветная проба (или цветная реакция) основана на разнице в цвете среды индикатором фенолрот, в которой растёт нормальная, жизнеспособная культура клеток, и среды, где находится культура, заражённая вирусом. В процессе развития нормальной культуры клеток происходит постепенное накопление кислых продуктов обмена веществ, что приводит к сдвигу рН в кислую сторону. Такое изменение реакции среды говорит о наличии роста и размножения клеток. Среда с индикатором фенолрот, первоначально имевшая красный цвет (рН 7,4 – 7,6), вследствие снижения рН до 7,0 – 6,8 приобретает жёлтый цвет.
Заражение культуры клеток вирусом приводит к развитию в ней дегенеративных процессов: происходит подавление процессов метаболизма, значительно понижается гликолиз, в результате чего кислых продуктов накапливается мало, рН среды остаётся на исходном уровне и среда с индикатором фенолрот остаётся красного цвета.
С помощью цветной реакции можно определить соответствие вируса и вируснейтрализующей сыворотки, если их предварительно смешать и эту смесь после инкубации внести в культуру клеток. Специфическая сыворотка нейтрализует вирус и он не оказывает цитопатическое действие на клетки культуры.
Цветную пробу ставят с различными типами клеточных культур. Чаще берут первичную культуру клеток почки обезьяны или перевиваемые культуры клеток. При постановке цветной реакции необходимо учитывать, что каждая культура клеток, на которой ставят реакцию, имеет определённый уровень метаболизма. Устанавливают дозу клеток для цветной реакции, то есть то оптимальное количество клеток, которое должно быть взято в каждую пробирку.
При работе с культурой почечных клеток обезьяны готовят взвесь клеток густотой 100-200 тыс. в 1 мл. Из неё делают ряд возрастающих разведений в объёме 0,25 мл и определяют минимальное количество клеток, которое при постановке цветной пробы изменяет цвет питательной среды из красного в жёлтый н 5-6-й день выдерживания в термостате; это количество и принимается за дозу клеток. Обычно эта доза – 25 тыс. клеток в объёме 0,25 мл.
Титрование вируса методом цветной пробы
Цветную пробу ставят в пробирках, которые иногда закрывают резиновыми пробками, но чаще разобщение от атмосферного воздуха достигается наслаиванием стерильного вазелинового масла (0,6-0,8 мл) или применение алюминиевой фольги. Однотипную опытную смесь наливают в четыре пробирки.
При оценке результатов реакции учитывают два тона: жёлтый и красный (табл. 3).
При титровании вируса по цветной пробе готовят десятикратные разведения вируссодержащего материала. Эти разведения вируса, взятые в объёме 0,25 мл, смешивают с 1 дозой клеток, содержащейся в таком же объёме, добавляют питательную среду по 0,25 мл и наслаивают вазелиновое масло по 0,6-0,8 мл. В реакции ставят контроли клеточной взвеси: берут 1,1/2 и ¼ дозы клеток. Пробирки помещают на 5-6 дней в термостат при 37 0 С после чего по изменению цвета учитывают реакцию.
Титром вируса по цветной пробе называется то наибольшее разведение вируса, которое вызывает подавление метаболической активности клеток в 50% случаев. В данном примере (табл. 3) титр вируса равен 10 -4 (две пробирки красные, две – жёлтые). Количество вируса, содержащееся в объёме 0,25 мл в разведении, равном титру, называется цитопатической дозой (ЦПД50) вируса.
Схема титрования вируса методом цветной пробы
Ингредиенты опыта (в мл)
№ пробирок
Разведения вируссодержащего материала
Контроль клеток
10 -1
10 -2
10 -3
10 -4
10 -5
10 -6
1 доза
½ доза
¼ доза
Вируссодержащий материал
0,25
0,25
0,25
0,25
0,25
0,25
-
-
-
Взвесь клеток (1 доза)
0,25
0,25
0,25
0,25
0,25
0,25
0,25
0,25
0,25
Питательная среда
0,25
0,25
0,25
0,25
0,25
0,25
0,5
0,5
0,5
Вазелиновое масло
0,6
0,6
0,6
0,6
0,6
0,6
0,6
0,6
0,6
Возможные результаты
Цвет среды в пробирках (красный или жёлтый)
К-4
К-4
К-3 Ж-1
К-2 титр Ж-2
К-1 Ж-3
Ж-4
Ж-4
Ж-2 К-2
К-4
Обозначения:
К-4 – красного цвета 4 пробирки; Ж-3 – жёлтого цвета 3 пробирки
Дата добавления: 2015-04-15 ; просмотров: 67 ; Нарушение авторских прав