
Особенности репликации вирусного генома
Геном вирусов может быть представлен одноцепочечными или двухцепочечными ДНК или РНК. Гены вирусов могут быть заключены в одной хромосоме или разделены на несколько блоков (хромосом) , которые все вместе и составляют геном таких вирусов. Например, у ретвирусов геном представлен двухцепочечной РНК и состоит из десяти сегментов. Геномы вирусов, содержащих одноцепочечную РНК, также могут быть либо цельными (у ретровирусов ), либо сегментированными (у аренавирусов ). Геном РНК-содержащих вирусов представлен только линейными молекулами РНК. Все известные ДНК- содержащие вирусы позвоночных имеют геном, заключенный в одной хромосоме, линейной или кольцевой, одно- или двухцепочечной.Репликация геномов вирусов ДНК-геномы реплицируются клеточными или вирусоспецифическими ДНК-полимеразами. РНК-геномы реплицируются вирусоспецифическими РНК-полимеразами, которые также являются и транскриптазами. Репликация вирусных геномов происходит или одновременно с транскрипцией, или эти два процесса разделены во времени. Механизмы репликации геномов вирусов многообразны и определяются видом генома. Существует три модели репликации – полуконсервативная, консервативная и дисперсная. Консервативная и дисперсная модели репликации нуклеиновых кислот установлены только у вирусов. Полуконсервативная модель предполагает, что после первого раунда репликации одна цепь в каждой из двух дочерних молекул является родительской, другая – синтезируемой заново. По такой схеме реплицируются двунитевыеДНК-геномы вирусов. При реализации консервативной модели репликации одна дочерняя молекула состоит из двух родительских цепей, а другая – из вновь синтезированных цепей. Согласно консервативной модели реплицируются двунитевые РНК ротавирусов .ДНК-содержащие вирусы, реплицирующиеся таким образом, неизвестны. Дисперсная модель репликации приводит к образованию молекулы нуклеиновой кислоты, состоящей из фрагментов, как родительских цепей, так и вновь синтезированных.
Строение геномов эукариот и особенности их репликации
Геном эукариотДля клеток эукариот характерно наличие оформленного ядра. Информационной макромолекулой их генома является ДНК, которая неравномерно распределена по нескольким хромосомам в виде комплексов с многочисленными белками. Жизненно важная генетическая информация заключена и во внехромосомных молекулах ДНК. У эукариот — это ДНК хлоропластов, митохондрий и других пластид. Геном эукариот обладает избыточностью. Эукариотическая клетка содержит во много раз больше генов, чем прокариотическая. Большая часть их геномной ДНК представлена некодирующими последовательностями нуклеотидов. У эукариот около 5% ДНК составляют экзоны (участки гена, кодирующие белок), 25% – интроны (участки гена, которые транскрибируются, но затем удаляются при сплайсинге), а остальные 70% составляют спейсеры – нетранскрибируемые участки ДНК между генами.Особенности репликации у эукариот.Инициация репликации. Репликация начинается на определённом сайте молекулы ДНК, точке начала репликации или ori-сайте. В этой точке двойная спираль ДНК расплетается с образованием двух репликативных вилок, которые движутся по направлению друг к другу .В формировании репликативных вилок участвуют ДНК-топоизоимеразы и ДНК-хеликазы. По окончанию формирования репликативной вилки фермент ликвидирует разрыв и отделяется от ДНК.Разрыв водородных связей в двуцепочечной молекуле ДНК осуществляет фермент ДНК-хеликаза. Для этого фермент использует энергию макроэргических связей АТФ. В результате происходит раскручивание суперпсиральтной структуры ДНК в поддержании такой структуры участвуют SSB-белки. Эти белки не закрывают азотистых оснований и не препятствуют репликации.Элонгация – продолжение репликации. Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами.Субстартами для синтеза новой цепи ДНК являются дезоксинуклеозидтрифосфаты.. Эти же соединения служат источниками энергии. На образование каждой связи расходуются две макроэргические связи. В процессе также участвуют ионы магния, нейтрализуя отрицательный заряд нуклеотидов.У эукариот принимают участие 5 ДНК полимераз (α, β, γ, δ, ε), которые различаются по числу субъединиц, молекулярной массе, способности взаимодействовать с другими белками и функциональному назначению. Полимеразы β, δ и ε не могут начинать образование дочерней ДНК, т.к. не имеют сродства к одиночной цепи ДНК. Начинает репликацию полимераза α, которая синтезирует небольшой фрагмент РНК-праймер и далее примерно 60 нуклеотидных остатков ДНК. Образующийся небольшой двухцепочечный фрагмент позволяет присоединиться полимеразе δ и продолжить синтез новой цепи в направлении от 5/ к 3/. Выбор очередного нуклеотида определяется матрицей ДНК. При созревании отстающей цепи ДНК праймеры удаляют ДНК-полимераза β и присоединяет недостающие нуклеотиды. Восстановленные фрагменты соединяют ДНК-лигазы.

Учебник предназначен для студентов, аспирантов и преподавателей высших медицинских учебных заведений, университетов, микробиологов всех специальностей и практических врачей.
5-е издание, исправленное и дополненное
Книга: Медицинская микробиология, иммунология и вирусология
Репликация вирусных геномов
Репликация вирусных геномов
Тип геномной вирусной ДНК определяет особенности ее репликации.
1. Двунитевая ДНК – репликация происходит по обычному механизму полуконсервативной репликации: нити разделяются, и на каждой из них достраивается комплементарная ей нить.
3. У вирусов, геном которых представлен однонитевой РНК, ее репликация происходит по следующей схеме: вначале на вирионной РНК (вРНК) синтезируются комплементарные ей РНК (кРНК). Этот процесс катализируется специфической РНК-репликазой I. Затем на кРНК синтезируется комплементарная ей, но идентичная исходной вирионная РНК (вРНК), этот процесс также катализируется специфической репликазой II. Таким образом, репликация идет по схеме (рис. 80. II):
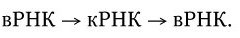
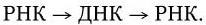
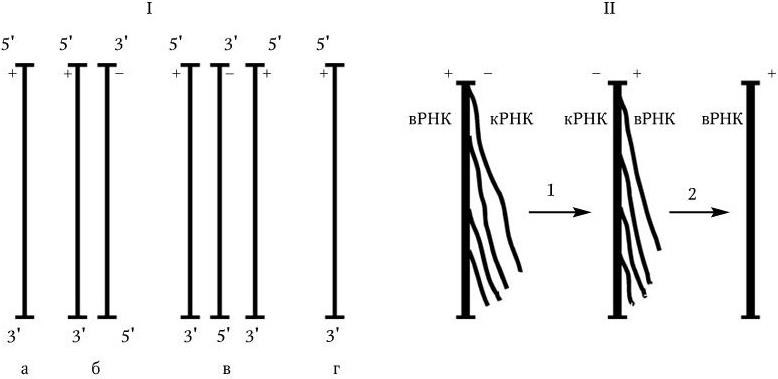
Рис. 80. Механизм репликации однонитевого вирусного ДНК-генома (I) и однонитевого вирусного РНК-генома (II):
а – геномная однонитевая ДНК; б – репликативная форма; в – промежуточная репликативная форма; г – однонитевая дочерняя геномная ДНК; вРНК – вирионная (геномная) РНК; кРНК – комплементарная РНК. Объяснение в тексте
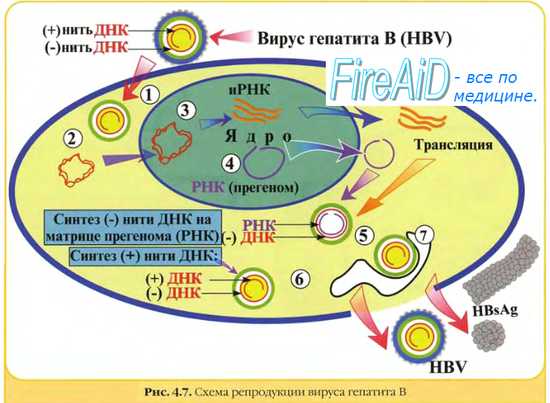
5. Размножение вируса гепатита В также протекает с участием обратной транскриптазы, но вначале клеточная РНК-полимераза синтезирует на вирусной ДНК прегеномную РНК, после чего вирусная ревертаза синтезирует на ней минус-цепь ДНК, которая достраивается плюс-цепью ДНК. В виде двунитевой ДНК вирус интегрируется в хромосому клетки-хозяина, где на ней транскрибируется вирусная РНК.
Существуют некоторые общие закономерности размножения вирусов. Во-первых, все РНК-содержащие вирусы, кроме вирусов гриппа и ретровирусов, размножаются в цитоплазме. Для своего размножения вирусы гриппа А и В и ретровирусы проникают в ядро, что связано с особенностями поведения их генома. Во-вторых, размножение всех ДНК-содержащих вирусов, кроме вирусов оспы, протекает в ядре, где происходит транскрипция и репликация их геномных нуклеиновых кислот, и в цитоплазме, где происходит трансляция вирусных белков, их процессинг и морфогенез вирионов. Лишь размножение вирусов группы оспы происходит в цитоплазме клетки, поскольку они обладают собственными системами транскрипции.
Другая особенность размножения вирусов заключается в том, что их нуклеокапсидные белки синтезируются на свободных полирибосомах (не связанных с мембраной), а суперкапсидные белки – на рибосомах, ассоциированных с мембранами (на шероховатых мембранах). Кроме того, белки некоторых вирусов подвергаются протеолитическому процессингу и гликозилированию. Различают два типа протеолитического процессинга: каскадный и точечный.
При точечном протеолизе разрезанию подвергается один (реже несколько) из вирусных полипептидов. Разрезание происходит, как правило, в определенном участке полипептида. Такой тип протеолиза необходим для того, чтобы определенный белок вируса приобрел свою специфическую активность. Например, суперкапсидный белок вируса гриппа – гемагглютинин – разрезается на две субъединицы: боEльшую и меньшую. В результате меньшая субъединица приобретает способность сливаться с мембранами клетки-мишени и ее лизосомами. Благодаря этому вирус гриппа приобретает способность проникать в клетку. Такой точечный протеолиз наблюдается у ортомиксовирусов, парамиксовирусов, ротавирусов, вирусов группы оспы и др. Точечный протеолиз, как и каскадный, жизненно важен для вируса.
Наконец, еще одна особенность вирусов, обладающих суперкапсидом, заключается в том, что суперкапсидные белки подвергаются в ходе своей транспортировки на наружную поверхность клеточной мембраны гликозилированию.
- 6588
- 5,5
- 0
- 5

Вопрос о происхождении вирусов
Существует три основные теории возникновения вирусов [1]:
Зарождение жизни. Идея последнего универсального общего предка: каким он мог бы быть и что ему предшествовало?

Рисунок 1. Схема трехдоменной классификации, предложенная Вёзе. В основании этой схемы должен находиться последний универсальный общий предок (англ. last universal common ancestor, LUCA). Рисунок из Википедии.
Самый сильный аргумент в пользу существования LUCA — сохранившаяся общая система экспрессии генов (передачи наследственной информации от гена с образованием РНК или белков), одинаковая для всех живущих организмов. Все известные клеточные формы жизни используют один и тот же генетический код из 20 универсальных аминокислот и стоп-сигналов, закодированных в 64 кодонах (единицах генетического кода). Трансляция генетической информации в процессе синтеза белков по заданной матрице выполняется рибосомами, состоящими из трех универсальных молекул РНК и примерно 50 белков, из которых 20 так же одинаковы для всех организмов.
В 2010 году американский биохимик Даглас Теобальд математически проверил вероятность существования LUCA [6]. Он выбрал 23 белка, встречающихся у организмов из всех трех доменов, но имеющих разную структуру у различных видов. И исследовал эти белки у 12 различных видов (по четыре из каждого домена), после чего использовал компьютерное моделирование различных эволюционных сценариев, чтобы понять, при каком из них наблюдаемая картина будет наиболее вероятной. Оказалось, что концепция, включающая существование универсального предка, значительно вероятнее концепций, где его нет. Еще более вероятна модель, основанная на существовании общего предка, но допускающая обмен генами между видами [7].
Предположение о том, что LUCA был прокариотической клеткой, похожей на современные, часто принимается по умолчанию. Однако мембраны архей и бактерий имеют разное строение (рис. 2). Получается, что общий предок должен был обладать комбинаторной мембраной. Новая информация о мембранах LUCA появилась в 2012 году, когда несколько групп ученых подробно проанализировали историю генов всех ферментов биосинтеза компонентов липидов у бактерий, архей и эукариот [8].

Родственными у архей и бактерий оказались ферменты для синтеза терпеновых спиртов и пришивания полярных голов к спиртам. Значит, эти реакции мог проводить и LUCA. Проще всего было предположить, что липиды LUCA состояли из одного остатка терпенового спирта, остатка фосфата и полярной группы (серина или инозитола). Подобные липиды были синтезированы искусственно. Образующиеся из них мембраны обладают высокой подвижностью по сравнению с современными мембранами, хорошо пропускают ионы металлов и малые органические молекулы. Это могло позволять древним протоклеткам поглощать готовую органику из внешней среды даже без транспортных белков.
Реконструкции LUCA методами сравнительной геномики указывают на то, что это должен быть сложный организм без обширного ДНК-генома (геном, состоящий из нескольких сотен РНК-сегментов или ДНК провирусного типа). Но даже если считать возможность существования общего предка доказанной, остается загадкой, в какой среде он мог бы появиться.
Предполагается, что идеальные условия для формирования жизни существовали вблизи термальных геоисточников (морских или наземных) в виде сети неорганических ячеек, обеспечивающих градиенты температуры и рН, способствующих первичным реакциям, и предоставляющих универсальные каталитические поверхности для примитивной биохимии [10].
Эти отсеки могли быть населены разнородной популяцией генетических элементов. Вначале сегментами РНК. Затем более крупными и сложными молекулами РНК (один или несколько белок-кодирующих генов). А позднее и сегментами ДНК, которые постепенно увеличивались (рис. 3).

Такие простейшие генетические системы использовали неорганические соединения из раствора и продукты деятельности других генетических систем. Сначала они должны были подчиняться индивидуальному отбору ввиду большого разнообразия. Но ясно, что важным фактором такого отбора была способность передавать генетическую информацию, то есть, копировать себя. Присутствие одновременно в одной ячейке молекул, способных копировать РНК, кодировать полезные белки и управлять синтезом новых молекул, давало больше шансов выживать в каждой отдельной ячейке. И в такой системе рано или поздно должны были появиться паразитирующие элементы. А если это так, то вирусные элементы стоят у самых истоков эволюции [11].
Возникновение паразитов — неизбежное последствие эволюционного процесса

Рисунок 4. Схематическое представление структуры модели эволюции РНК-подобной системы. На втором этапе цепочки последовательностей начинают соединяться комплементарными связями сами с собой. В результате у двух видов (cat-C и cat-A) возникает вторичная структура молекулы, которая обладает каталитическим свойством. Она ускоряет собственную репликацию (или репликацию несвернувшихся соседей). Два вида при этом приобретают паразитические свойства (par-G и par-U). Пояснения в тексте. Рисунок из [12].
Таким образом, паразитарные репликаторы способствуют эволюции разнообразия, вместо того, чтобы мешать этому разнообразию. Это также делает существующую систему репликатора чрезвычайно стабильной при эволюции паразитов.
Согласно гипотезе Черной Королевы, чтобы поддержать свое существование в постоянно эволюционирующем мире, вид должен реагировать на эти эволюционные изменения и должным образом приспосабливаться к среде. Поэтому, если мы говорим о вирусах как о паразитах, мы обязаны представлять себе взаимоотношения вируса с хозяином. В борьбе с вирусом хозяева развивают новые защитные механизмы, а паразиты отвечают, развивая механизмы для атаки и взлома защиты. Этот процесс может длиться бесконечно либо до вымирания одной из противоборствующих сторон. Так множественные системы защиты составляют существенную часть геномов всех клеточных организмов, а взлом защиты — одна из основных функций генов у вирусов с большими геномами*.
Механизмы клеточной защиты против вирусов
Механизмы защиты от вирусов стандартны, поскольку все вирусы уникальны, и приспособиться к каждому не представляется возможным. Это такие механизмы как:
- Деградация РНК (вирусных и клеточных) — РНК-интерференция;
- Угнетение синтеза белков (вирусных и клеточных);
- Ликвидация зараженных клеток — апоптоз (программируемая клеточная смерть);
- Воспаление.
Получается, что клетка борется с вирусом, нарушая собственные обмен веществ и/или структуру. Защитные реакции клетки — это в основном самоповреждающие механизмы.
Вирус заражает конкретную клетку потому, что его механизмы нападения направлены именно против данного типа клеток. Это такие механизмы как:
- Угнетение синтеза клеточной РНК;
- Угнетение синтеза клеточных белков;
- Нарушение клеточной инфраструктуры и транспорта;
- Подавление/включение апоптоза и других видов клеточной смерти.
Схемы защитных приемов клетки и противозащиты вирусов во многом идентичны. Вирусы и клетки применяют одни и те же приемы. Для подавления синтеза вирусных белков клетка использует интерферон, а чтобы подавить образование интерферона, вирус угнетает синтез белков.
Поскольку узнавание вируса неспецифическое, клетка не может знать намерения конкретного вируса. Она может бороться с вирусом лишь стандартными приемами, поэтому ее оборонные действия часто могут быть чрезмерными.
Понятие о вирусном геноме, типы вирусных генов, концепция генов-сигнатур
В исследовании, проведенном вирусологом Евгением Куниным и его коллегами [16], анализ последовательностей вирусных геномов выявил несколько категорий вирусных генов, принципиально отличающихся по происхождению. Можно обсуждать, какая степень дробности классификации оптимальна, но четко различаются пять классов, укладывающихся в две более крупные категории.
Гены с четко опознаваемыми гомологами у клеточных форм жизни:
- Гены, присутствующие у узких групп вирусов (обычно это гены, гомологичные генам хозяев этих вирусов).
- Гены, консервативные среди большой группы вирусов или даже нескольких групп и имеющие относительно отдаленные клеточные гомологи.
Таким образом, отличительные особенности генов-сигнатур:
- Происхождение из первичного пула генов;
- Наличие лишь очень отдаленных гомологов среди генов клеточных форм жизни, из чего можно сделать вывод, что они никогда не входили в геномы клеточных форм;
- Необходимость для репродукции вирусов.
Из всего вышесказанного следует, что эти гены переходили от вируса к вирусу (или к элементу, подобному вирусу) на протяжении четырех миллиардов лет эволюции жизни, а вирусные геномы появились благодаря перемешиванию и подгонке друг к другу генов в гигантской генетической сети, которую представляет собой мир вирусов. Многочисленные гены клеточных форм жизни также пронизывают эту сеть, прежде всего благодаря геномам крупных вирусов, таких как NCDLV и крупным бактериофагам, которые позаимствовали множество генов от своих хозяев на разных этапах эволюции. Однако большинство заимствованных генов сами по себе не критичны для репликации и экспрессии вирусного генома (исключая некоторые случаи возможного неортологичного замещения генов-сигнатур); обычно эти гены участвуют во взаимодействии между вирусом и хозяином. Таким образом, несмотря на интенсивный взаимообмен генами с хозяевами, вирусы всегда происходят от других вирусов.
Вирусы, встроенные в геном, и горизонтальный перенос генов
В процессе эволюции многие вирусы встроились в геномы клеточных форм жизни путем горизонтального переноса генов (ГПГ). Впервые горизонтальный перенос был описан в 1959 году, когда ученые продемонстрировали передачу резистентности к антибиотикам между разными видами бактерий. В 1999 году Рави Джайн, Мария Ривера и Джеймс Лейк в своей статье писали о произошедшей значительной передаче генов между прокариотами [17]. Этот процесс, по-видимому, оказал некоторое влияние также и на одноклеточные эукариоты. В 2004 году Карл Вёзе опубликовал статью, в которой утверждал, что между древними группами живых организмов происходил массивный перенос генетической информации. В древнейшие времена преобладал процесс, который он называет горизонтальным переносом генов. Причем, чем дальше в прошлое, тем это преобладание сильнее [18].
Горизонтальный перенос генов — процесс, в котором организм передаёт генетический материал другому организму, не являющемуся его потомком. Горизонтальная передача генов реализуется через различные каналы генетической коммуникации — процессы конъюгации, трансдукции, трансформации, переноса генов в составе плазмидных векторов, вирусов, мобильных генетических элементов (МГЭ).
Трансдукция — перенос бактериофагом (агентами переноса генов, АПГ) в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг [19]. Такой бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент). В зависимости от типа трансдукции — неспецифической (общей), специфической или абортивной, геном фага или хозяина-бактерии может быть изменен тем или иным образом:
- При неспецифической трансдукции (рис. 5) ДНК клетки-хозяина включаются в частицу фага (дополнительно к его собственному геному или вместо него);
- При специфической трансдукции гены фага замещаются генами хозяина;
- При абортивной трансдукции внесённый фрагмент ДНК донора не встраивается в ДНК хозяина-реципиента, а остаётся в цитоплазме и не реплицируется. Это приводит к тому, что при клеточном делении он передаётся только одной из дочерних клеток и затем теряется в потомстве.

Рисунок 5. Схема общей трансдукции. Фото с сайта vkjournal.ru.
Наиболее известным примером специфической трансдукции служит трансдукция, осуществляемая фагом λ. Поскольку этот фаг при переходе в состояние профага включается в хромосому бактерий между генами, кодирующими синтез галактозы и биотина, именно эти гены он может переносить при трансдукции.
Вот несколько примеров важных эволюционных событий, связанных с молекулярным одомашниванием:
- Ферменты теломеразы, служащие для восстановления концевых участков хромосом, возможно, ведут свое происхождение от обратных транскриптаз, кодируемых ретровирусами и ретротранспозонами [22];
- Белки RAG, играющие ключевую роль в системе адаптивного иммунитета, по-видимому, происходят от прирученных транспозаз — ферментов, кодируемых транспозонами;
- Ген Peg10, необходимый для развития плаценты, был позаимствован древними млекопитающими у ретротранспозона (рис. 6) [23].

Рисунок 6. Роль гена Peg10 в эмбриональном развитии. Ученые под руководством Рюичи Оно из Токийского медицинского университета Японии показали, что у мышей с выключенным геном Peg10 нарушается развитие плаценты, от чего эмбрион погибает через 10 дней после зачатия [24]. Фото с сайта flickr.com.
В 2008 году в ходе целенаправленного поиска неиспорченных вирусных генов в геноме человека исследователи нашли два очень похожих друг на друга ретровирусных гена (их назвали ENVV1 и ENVV2), которые, по всей видимости, находятся в рабочем состоянии [25]. Это гены белков оболочки ретровируса. Каждый из них входит в состав своего эндогенного ретровируса (ЭРВ), причем все остальные части этих ЭРВ давно не функционируют.
Вирусные гены ENVV1 и ENVV2 у человека и обезьян работают в плаценте и, скорее всего, выполняют следующие функции:
Таким образом, как минимум три полезных применения нашли себе вирусные гены в плаценте приматов. Это показывает, что генетические модификации, которым ретровирусы подвергают организмы, в долгосрочной перспективе могут оказаться полезными или даже определить развитие вида. И с учетом всего вышесказанного древо доменов должно выглядеть как на схеме ниже (рис. 7).

Рисунок 7. Горизонтальный перенос генов в рамках трехдоменного дерева. Рисунок из [26].
Заключение
Возникновение паразитов — обязательная черта эволюционирующих систем репликаторов, а соревнование хозяев и паразитов движет эволюцию тех и других. Любой организм является результатом миллионов лет борьбы клеток с невероятно разнообразным миром вирусов. Их действия и их эволюция пронизывают всю историю клеточной эволюции, и сейчас меняется само наше представление о них. Когда-то вирусы считали деградировавшими клетками, но чем больше мы узнаем о вирусах, тем очевиднее, что их роль в общей эволюции значительна. И невероятно много нам еще предстоит узнать.
Статья написана в соавторстве с Евгенией Щепенок.
Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности.
Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
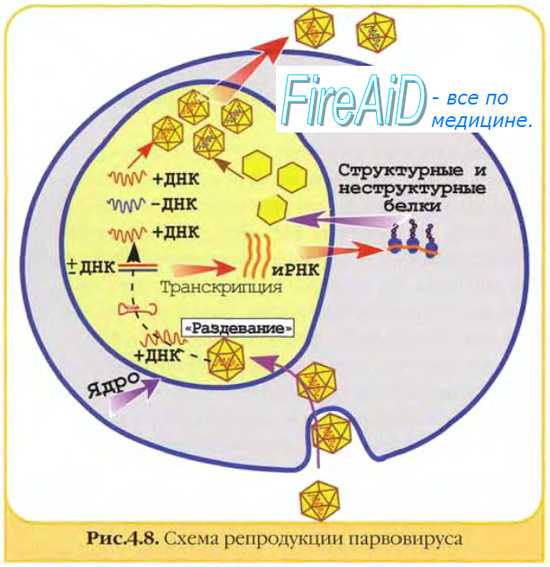
Матричные белкинеобходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе).
Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова- и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
начальный период включает стадии адсорбции вируса на клетке, проникновения в клетку, дезинтеграции (депротеинизации) или "раздевания" вируса. Вирусная нуклеиновая кислота была доставлена в соответствующие клеточные структуры и под действием лизосомальных ферментов клетки освобождается от защитных белковых оболочек. В итоге формируется уникальная биологическая структура: инфицированная клетка содержит 2 генома (собственный и вирусный) и 1 синтетический аппарат (клеточный);
• после этого начинается вторая группа процессов репродукции вируса, включающая средний и заключительный периоды, во время которых происходят репрессия клеточного и экспрессия вирусного генома. Репрессию клеточного генома обеспечивают низкомолекулярные регуляторные белки типа гистонов, синтезируемые в любой клетке. При вирусной инфекции этот процесс усиливается, теперь клетка представляет собой структуру, в которой генетический аппарат представлен вирусным геномом, а синтетический аппарат — синтетическими системами клетки.
2. Дальнейшее течение событий в клетке направлено на репликацию вирусной нуклеиновой кислоты(синтез генетического материала для новых вирионов) и реализацию содержащейся в ней генетической информации (синтез белковых компонентов для новых вирионов). У ДНК-содержащих вирусов, как в прокариотиче-ских, так и в эукариотических клетках, репликация вирусной ДНК происходит при участии клеточной ДНК-зависимой ДНК-полимеразы. При этом у однонитевых ДНК-содержащих вирусов сначала образуется комплементарная нить — так называемая репликативная форма, которая служит матрицей для дочерних молекул ДНК.
3. Реализация генетической информации вируса, содержащейся в ДНК, происходит следующим образом: при участии ДНК-зависимой РНК-полимеразы синтезируются и-РНК, которые поступают на рибосомы клетки, где и синтезируются вирусспе-цифические белки. У двунитевых ДНК-содержащих вирусов, геном которых транскрибируется в цитоплазме клетки хозяина, это собственный геномный белок. Вирусы, геномы которых транскрибируются в ядре клетки, используют содержащуюся там клеточную ДНК-зависимую РНК-полимеразу.
У РНК-содержащих вирусов процессы репликацииих генома, транскрипции и трансляции генетической информации осуществляются иными путями. Репликация вирусных РНК, как минус-, так и плюс-нитей, осуществляется через репликативную форму РНК (комплементарную исходной), синтез которой обеспечивает РНК-зависимая РНК-полимераза — это геномный белок, который есть у всех РНК-содержащих вирусов. Репликативная форма РНК минус-нитевых вирусов (плюс-нить) служит не только матрицей для синтеза дочерних молекул вирусной РНК (минус-нитей), но и выполняет функции и-РНК, т. е. идет на рибосомы и обеспечивает синтез вирусных белков (трансляцию).
У плюс-нитевыхРНК-содержащих вирусов функцию трансляции выполняют ее копии, синтез которых осуществляется через репликативную форму (минус-нить) при участии вирусных РНК-зависимых РНК-полимераз.
У некоторых РНК-содержащих вирусов (реовирусы) имеется совершенно уникальный механизм транскрипции. Он обеспечивается специфическим вирусным ферментом — ревертазой (обратной транскриптазой) и называется обратной транскрипцией. Суть ее состоит в том, что вначале на матрице вирусной РНК при участии обратной транскрипции образуется транскрипт, представляющий собой одну нить ДНК. На нем с помощью клеточной ДНК-зависимой ДНК-полимеразы синтезируется ,вторая нить и формируется двунитевой ДНК-транскрипт. С него обычным путем через образование и-РНК происходит реализация информации вирусного генома.
Результатом описанных процессов репликации, транскрипции и трансляции является образование дочерних молекул вирусной нуклеиновой кислоты и вирусных белков, закодированных в геноме вируса.
После этого наступает третий, заключительный период взаимодействия вируса и клетки. Из структурных компонентов (нуклеиновых кислот и белков) на мембранах цитоплазматического ретикулума клетки собираются новые вирионы. Клетка, геном которой был репрессирован (подавлен), обычно гибнет. Вновь сформировавшиеся вирионы пассивно (в результате гибели клетки) или активно(путем почкования) покидают клетку и оказываются в окружающей ее среде.
Таким образом, синтез вирусных нуклеиновых кислот и белков и сборка новых вирионов происходят в определенной последовательности (разобщены во времени) и в разных структурах клетки (разобщен в пространстве), в связи с чем способ репродукции вирусов и был назван дизъюнктивным (разобщенным). При абортивной вирусной инфекции процесс взаимодействия вируса с клеткой по тем или иным причинам прерывается до того, как произошло подавление клеточного генома. Очевидно, что в этом случае генетическая информация вируса реализована не будет и репродукции вируса не происходит, а клетка сохраняет свои функции неизменными.
При латентной вирусной инфекции в клетке одновременно функционируют оба генома, а при вирус-индуцированных трансформациях вирусный геном становится частью клеточного, функционирует и наследуется вместе с ним.
Дата добавления: 2018-05-09 ; просмотров: 471 ;
Читайте также: