
Репродукция вирусов стадии взаимодействия вируса с клеткой хозяина
Этапы взаимодействия вируса с клеткой хозяина:
1. Адсорбция – пусковой механизм, связанный с прикреплением вириона к поверхности клетки. Выделяют две основных фазы адсорбции:
а) неспецифическая обусловлена ионным притяжением между вирусом и клеткой.
б) высокоспецифическая обусловлена гомологией, комплементарностью специфических рецепторов вируса и хозяина (у вируса гриппа – гемагглютинин к сиаловой кислоте гликопротеинов клеток дыхательных путей, у вируса иммунодефицита человека - гликопротеин gp 120 - к СД4 рецепторам Т-хелперов, моноцитов, макрофагов).
Проникновение
а) путем слияния оболочки вириона с мембраной клетки, характерно для некоторых оболочечных вирусов (парамиксовирусов, ретровирусов, герпесвирусов)
б) путем эндоцитоза (пиноцитоза) в результате захватывания и поглощения вириона клеткой: клеточная мембрана с прикрепленным вирионом впячивается с образованием внутриклеточной вакуоли (эндосомы), содержащей вирус.
3. Освобождение нуклеиновых кислот (депротеинизация) – “раздевание” нуклеокапсида и активация нуклеиновой кислоты. В результате депротеинизации удаляются поверхностные структуры вируса и высвобождается его внутренний компонент, способный вызвать инфекционный процесс.
4. Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство вируса. Синтез нуклеиновых кислот и белков вируса разобщен во времени и пространстве, т.к. осуществляется в разных частях клетки. Такой способ размножения вирусов называется дизъюнктивным.
Синтез белков в клетке осуществляется благодаря процессам транскрипции – переписывания генетической информации с нуклеиновой кислоты в нуклеотидную последовательность информационной РНК (иРНК или мРНК) и трансляции – механизмов, при помощи которых последовательность нуклеотидных оснований мРНК переводится в специфическую последовательность аминокислот в синтезируемом полипептиде на рибосомах клетки хозяина.
5. Сборка вирионов – многоступенчатый процесс, включающий в себя соединение всех компонентов вириона.
Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов. У сложно устроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса). У минус-нитевых РНК вирусов в сборку вовлекается матриксный белок (М-белок), который расположен под модифицированной клеточной мембраной.
6. Выход вирусных частиц из клетки взрывным путем или почкованием, экзоцитозом. Полный цикл репродукции вирусов завершается через 5-6 ч (вирус гриппа) или через несколько суток (вирус кори). По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки. Из погибающей клетки одновременно выходит большое количество вирионов. Почкование, экзоцитоз характерен вирусам, имеющим липопротеиновую оболочку, которая является производной клеточных мембран. Сначала образовавшийся нуклеокапсид транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. Клетка способна длительное время сохранять жизнеспособность и продуцировать вирусное потомство.
Исходы взаимодействия вирусов с клеткой хозяина
1. Продуктивный процесс - репликация (продукция) вирусов. Выделяют:
· продуктивный цитолитический процесс с образованием инфекционного потомства и выходом вирионов во внеклеточную среду.
· продуктивной нецитолитический процесс с образованием инфекционных вирусных частиц без лизиса клетки, которая продолжает функционировать.
Цитопатический эффект является результатом интенсивного размножения и формирования большого количества вирусных частиц (характерный результат продуктивного процесса), вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит узнаваемый специфический характер;
2. Интегративный процесс - интеграция вирусного генома с геномом клетки хозяина (вирогения). Это особый вариант продуктивного процесса по типу стабильного взаимодействия. Вирус реплицируется вместе с геномом клетки хозяина и может длительно находиться в латентном состоянии. Встраиваться в ДНК - геном хозяина могут только ДНК- вирусы (принцип “ДНК - в ДНК”). Единственные РНК - вирусы, способные интегрироваться в геном клетки хозяина - ретровирусы, имеют для этого специальный механизм. Синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы (ревертазы) с последующим встраиванием ДНК в геном хозяина.
3. Абортивный процесс - клетки освобождаются от вируса, не вызывая появление дочерней популяции или появление в меньшем количестве. При инфицировании дефектным вирусом, для репликации которого нужен вирус- помощник, самостоятельная репликация этих вирусов невозможна. Например, дельта вирус или вирус гепатита D может реплицироваться в клетке только при наличии вируса гепатита B, его HВs – антигена. При инфицировании вирусом генетически нечувствительных к нему клеток или при заражении чувствительных клеток вирусом в неблагоприятных условиях.
Возможные последствия инфекционного процесса, вызванного вирусами для клетки:
- сохранение жизнеспособности клетки
- деструкция клетки, возникающая при цитолитической инфекции (цитопатогенное действие вируса - ЦПД)
- образование многоядерных клеток в результате их слияния (симпластообразование)
- образование вирусных внутриклеточных включений
· онкогенная трансформация клетки при интеграции вирусного генома с геномом клетки (вирогении, интегративной инфекции)
Дата добавления: 2019-09-13 ; просмотров: 578 ;
Вирусы не способны размножаться на питательных средах – это строгие внутриклеточные паразиты. Более того, в отличие от риккетсий и хламидий, вирусы в клетке хозяина не растут и не размножаются путем деления. Составные части вируса – нуклеиновые кислоты и белковые молекулы синтезируются в клетке хозяина раздельно, в разных частях клетки – в ядре и цитоплазме. При этом клеточные белоксинтезирующие системы подчиняются вирусному геному, его НК.
Репродукция вируса в клетке происходит в несколько фаз:
1)первая фаза – адсорбция вируса на поверхности клетки, чувствительной к данному вирусу.
2)вторая фаза – проникновение вируса в клетку хозяина путем виропексиса.
В зависимости от типа нуклеиновой кислоты этот процесс совершается следующим образом.
ДНК-содержащие (ДНК- и РНК-белок):
1)репродукция происходит в ядре: аденовирусы, герпес, паповавирусы. Используют ДНК-зависимую РНК-полимеразу клетки.
2)репродукция происходит в цитоплазме: вирусы имеют свою ДНК-зависимую РНК-полимеразу.
1)рибовирусы с позитивным геномом (плюс-нитиевые): пикорна-, тога-, коронавирусы. Транскрипции нет.
2)рибовирусы с негативным геномом (минус-нитиевые): грипп, корь, паротит, орто-, парамиксовирусы.
(-)РНК, иРНК-белок (иРНК-комплементарная (-)РНК). Этот процесс идет при участии специального вирусного фермента – вирионная РНК-зависимая РНК-полимераза (в клетке такого фермента быть не может).
3)ретровирусы (-)РНК, ДНК, иРНК-белок (иРНК гомологична РНК). В этом случае процесс образования ДНК на базе (-)РНК возможен при участии фермента – РНК-зависимой ДНК-полимеразы (обратнойтранскриптазы или ревертазы).
4)четвертая фаза – синтез компонентов вириона. Нуклеиновая кислота вируса образуется путем репликации. На рибосомы клетки транслируется информация вирусной иРНК, и в них синтезируется вирус-специфический белок.
5)пятая фаза – сборка вириона. Путем самосборки образуются нуклеокапсиды.
6)шестая фаза – выход вирионов из клетки. Простые вирусы, например, вирус полиомиелита, при выходе из клетки разрушают ее. Сложноорганизованные вирусы, например, вирус гриппа, выходят из клетки путем почкования. Внешняя оболочка вируса (суперкапсид) формируется в процессе выхода вируса из клетки. Клетка при таком процессе на какое-то время остается живой.
Описанные типы взаимодействия вируса с клеткой называются продуктивными, так как приводят к продукции зрелых вирионов.
Кроме обычных вирусов, существуют прионы – белковые инфекционные частицы, не содержащие нуклеиновую кислоту. Они имеют видфибрилл, размером до 200 нм. Вызывают у человека и у животных медленные инфекции с поражением мозга: болезнь Крейтцфельда-Якоба, куру, скрепи и другие.
4)Особенности репродукции вирусов в зависимости от типа нуклеиновой кислоты (+ и – РНК). Виды взаимодействия вирусов и клетки: продуктивный, абортивный, интегративный.
В зависимости от типа нуклеиновой кислоты этот процесс совершается следующим образом.
ДНК-содержащие (ДНК- и РНК-белок):
1)репродукция происходит в ядре: аденовирусы, герпес, паповавирусы. Используют ДНК-зависимую РНК-полимеразу клетки.
2)репродукция происходит в цитоплазме: вирусы имеют свою ДНК-зависимую РНК-полимеразу.
1)рибовирусы с позитивным геномом (плюс-нитиевые): пикорна-, тога-, коронавирусы. Транскрипции нет.
2)рибовирусы с негативным геномом (минус-нитиевые): грипп, корь, паротит, орто-, парамиксовирусы.
(-)РНК, иРНК-белок (иРНК-комплементарная (-)РНК). Этот процесс идет при участии специального вирусного фермента – вирионная РНК-зависимая РНК-полимераза (в клетке такого фермента быть не может).
3)ретровирусы (-)РНК, ДНК, иРНК-белок (иРНК гомологична РНК). В этом случае процесс образования ДНК на базе (-)РНК возможен при участии фермента – РНК-зависимой ДНК-полимеразы (обратнойтранскриптазы или ревертазы).
Типы взаимодействия вируса с клеткой.
Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.Продуктивный тип— завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитолитическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип— не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Адсорбция. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц.
Проникновение в клетку. Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга.
Биосинтез компонентов вируса. Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства.Реализация генетической информации вируса осуществляется в соответствии с процессами транскрипции, трансляции и репликации.
Существуют следующие общие принципы сборки вирусов, имеющих разную структуру:
1. Формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
2. Сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
3. Формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
4. Сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
Вирусы — мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царству Vira. Являясь облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом.
Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
Форма вирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы.
Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид.
Сложные, или оболочечные, вирусы снаружи капсида окружены липопротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок.
Тип симметрии. Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вируса гриппа).
Включения — скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом при специальном окрашивании. Вирус натуральной оспы образует цитоплазмати-ческие включения — тельца Гварниери; вирусы герпеса и аденовирусы — внутриядерные включения.
Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным — натуральной оспы (около 350 нм).
Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Вирусы поражают позвоночных и беспозвоночных животных, а также растения и бактерии. Являясь основными возбудителями инфекционных заболеваний человека, вирусы также участвуют в процессах канцерогенеза, могут передаваться различными путями, в том числе через плаценту (вирус краснухи, цитомегаловирус и др.), поражая плод человека. Они могут приводить к постинфекционным осложнениям — развитию миокардитов, панкреатитов, иммунодефицитов и др.
Кроме обычных вирусов, известны и так называемые неканонические вирусы — прионы — белковые инфекционные частицы, являющиеся агентами белковой природы, имеющие вид фибрилл размером 10—20x100—200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирусной инфекции (болезни Крейтц-фельдта—Якоба, куру и др.).
Другими необычными агентами, близкими к вирусам, являются вироиды — небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белка, вызывающие заболевания у растений.
Типы взаимодействия вируса с клеткой. Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.
Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитолитическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения — характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Адсорбция. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц.
Проникновение в клетку. Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга.
Биосинтез компонентов вируса. Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства.
Реализация генетической информации вируса осуществляется в соответствии с процессами транскрипции, трансляции и репликации.
Существуют следующие общие принципы сборки вирусов, имеющих разную структуру:
1. Формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
2. Сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
3. Формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
4. Сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции.
Структура и химический состав бактериофагов. Взаимодействие фага с бактериальной клеткой. Умеренные и вирулентные бактериофаги. Лизогения. Применение фагов в биотехнологии, микробиологии и медицине.
Бактериофаги — вирусы бактерий, обладающие способностью специфически проникать в бактериальные клетки, репродуцироваться в них и вызывать их растворение (лизис).
В зависимости от формы и структурной организации фаги подразделяют на несколько морфологических типов: нитевидные, мелкие кубические, фаги сперматозоидной формы.
Взаимодействие фага с бактериальной клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги.
После биосинтеза фаговых компонентов и их самосборки в бактериальной клетке накапливается до 200 новых фаговых частиц. Под действием фагового лизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Один литический цикл (от момента адсорбции фагов до их выхода из клетки) продолжается 30—40 мин. Процесс бактериофагии проходит несколько циклов, пока не будут лизированы все чувствительные к данному фагу бактерии.
Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий.
Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геном бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке неограниченному числу потомков.
Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это название отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии.
Лизогенные культуры по своим основным свойствам не отличаются от исходных, но они невосприимчивы к повторному заражению гомологичным или близкородственным фагом и, кроме того, приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии. Последняя имеет место у многих видов микроорганизмов и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов.
По содержанию бактериофагов в объектах окружающей среды (например, в воде) можно судить о присутствии в них соответствующих патогенных бактерий. Подобные исследования проводят при эпидемиологическом анализе вспышек инфекционных болезней.
Фаги применяют также для лечения и профилактики ряда бактериальных инфекций. Производят брюшнотифозный, сальмонеллезный, дизентерийный, синегнойный, стафилококковый, стрептококковый фаги и комбинированные препараты (колипротейный, пиобактериофаги и др). Бактериофаги назначают по показаниям перорально, парентерально или местно в виде жидких, таблети-рованных форм, свечей или аэрозолей.
Бактериофаги широко применяют в генной инженерии и биотехнологии в качестве векторов для получения рекомбинантных ДНК.
Для вирусов характерен дизъюнктивный (от disjuncus — разобщенный) способ репродукции-размножения. Потомство вируса возникает в результате сборки нуклеиновых кислот и белковых субъединиц, которые синтезируются раздельно клеткой хозяина.
Проникновение вируса в клетку и воспроизведение себе подобных проходит в несколько фаз:
1.проникновение в клетку хозяина,
2.синтез ферментов, необходимых для репликации вирусных нуклеиновых кислот,
3.синтез вирусных частей,
4.сборка и композиция зрелых вирионов,
5.выход зрелых вирионов из клетки.
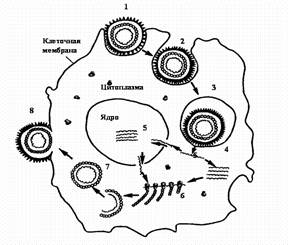
Стадии репродукции вирусов.
1 - адсорбция вириона на клетке; 2 - проникновение вириона в клетку путем виропексиса;
3 - вирус внутри вакуоли клетки; 4 - `раздевание вириона вируса; 5 - репликация вирусной нуклеиновой кислоты; 6 - синтез вирусных белков на рибосомах клетки; 7 - формирование вириона; 8 - выход вириона из клетки путем почкования.
Фаза I — адсорбция вириона на поверхности клетки.
Протекает в две стадии: первая — неспецифическая, когда вирус удерживается на поверхности клетки при помощи электростатических сил, т. е. благодаря возникновению противоположных зарядов между отдельными участками мембраны клеток и вируса. Эта фаза взаимодействия вируса с клеткой обратима, на нее оказывают влияние такие факторы, как рН и солевой состав среды.
Вторая стадия — специфическая, когда взаимодействуют специфические рецепторы вируса и рецепторы клетки, комплементарные друг другу. По химической природе рецепторы клетки могут быть мукопротеидами (или мукополисахаридами) и липопротеидами. Разные вирусы фиксируются на разных рецепторах: вирусы гриппа, парагриппа, аденовирусы — на мукопротеидах, а вирусы клещевого энцефалита, полиомиелита — на липопротеидах.
Фаза II — проникновение вируса в клетку. Электроноскопические наблюдения за процессом проникновения вирусов в чувствительные к ним клетки показали, что оно осуществляется посредством механизма, напоминающего пиноцитоз, или, как чаще называют, виропексис. В месте адсорбции вируса клеточная стенка втягивается внутрь клетки, образуется вакуоль, в которой оказывается вирион. Параллельно клеточные ферменты (липазы и протеазы) вызывают депротеинизацию вириона — растворение белковой оболочки и освобождение нуклеиновой кислоты.
Фаза III — скрытый период (период эклипса — исчезновения). В этот период в клетке невозможно определить наличие инфекционного вируса ни химическими, ни электронно-микроскопическими, ни серологическими методами. О сущности этого явления и его механизмов пока известно мало. Предполагается, что в скрытой фазе нуклеиновая кислота вируса проникает в хромосомы клетки и вступает с ними в сложные генетические взаимоотношения.
Фаза IV — синтез компонентов вириона. В этой фазе вирус и клетка представляют единое целое, вирусная нуклеиновая кислота выполняет генетическую функцию, индуцирует образование ранних белков и изменяет функцию рибосом. Ранние белки подразделяются на:
а) белки-ингибиторы (репрессоры), подавляющие метаболизм клеток
б) белки-ферменты (полимеразы), обеспечивающие синтез вирусных нуклеиновых кислот.
Синтез нуклеиновых кислот и белков протекает неодновременно и в разных структурных частях клетки. У вирусов, содержащих ДНК или РНК, эти процессы имеют некоторые различия и особенности.
Процесс формирования вирионов начинается спустя определенное время после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность этого периода довольно вариабельна и предопределяется природой вируса — для РНК-содержащих обычно короче, чем для ДНК-вирусов. Например, продукция полных вирусных частиц осповакцины начинается приблизительно спустя 5—6 ч после инфицирования клеток и продолжается в течение последующих 7—8 ч, т. е. после того как синтез вирусной ДНК уже завершен.
Между нуклеиновой кислотой и соответствующим белковыми субъединицами образуются очень прочные связи, о чем свидетельствуют трудности отделения белка от вирусной нуклеиновой кислоты. Большую прочность вирусной частице придают входящие в ее состав углеводы и особенно липиды.
Формирование вирионов, так же как и синтез компонентов вируса, происходит в разных местах клетки, при участии различных клеточных структур. После завершения процесса формирования образуется зрелая дочерняя вирусная частица, обладающая всеми свойствами родительского вириона. Но иногда наблюдается образование так называемых неполных вирусов, которые состоят или только из нуклеиновой кислоты, или из белка, или из вирусных частиц, формирование которых остановилось в какой-то промежуточной стадии.
Фаза VI — выход зрелых вирионов из клетки. Существуют два основных механизма выхода зрелых вирионов из клетки:
1) выход вириона с помощью почкования. В этом случае наружная оболочка вириона происходит из клеточной мембраны, она содержит как материал клетки хозяина, так и вирусный материал;
2) выход зрелых вирионов из клетки через бреши в мембране. Эти вирусы не имеют наружной оболочки. При таком механизме выхода вирусов клетка, как правило, погибает и в среде появляется большое количество вирусных частиц.
Причиной гибели зараженной клетки могут быть три механизма:
2.защитная реакция клетки, запускающая генетическую программу ее гибели (апоптоз);
3. иммунная система организма, уничтожающая зараженную клетку.
Кроме продуктивного типа взаимодействия вируса и клетки возможно интегративное сосуществование или вирогения. Вирогения характеризуется интеграцией (встраиванием) нуклеиновой кислоты вируса в геном клетки, а также репликацией и функционированием вирусного генома как составной части генома клетки. Для интеграции с клеточным геномом необходимо возникновение кольцевой формы двунитевой ДНК вируса. Встроенная в состав хромосомы клетки вирусная ДНК называется провирусом. Провирус реплицируется в составе хромосомы и переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. Под влиянием некоторых физических или химических факторов провирус может переходить в автономное состояние с развитием продуктивного типа взаимодействия с клеткой. Дополнительная генетическая информация провируса при вирогении сообщает клетке новые свойства, что может быть причиной развития опухолей, аутоиммунных и хронических заболеваний. На способности вирусов к интеграции с геномом клетки основаны персистенция (от лат. persisto - постоянно пребывать, оставаться) вирусов в организме и развитие персистентных вирусных инфекций. Например, вирус гепатита В способен вызывать персистирующие поражения с развитием хронического гепатита и часто опухолей печени.
Читайте также: