
Типы генетического материала и механизм его репликации у различных вирусов
Структурная организация и аминокислотный состав белков. Первичная структура нуклеиновых кислот. Типы генетического материала и механизм его репликации у различных вирусов. Взаимодействие вируса с клеткой-хозяином. Последовательности нуклеотидов.
Нажав на кнопку "Скачать архив", вы скачаете нужный вам файл совершенно бесплатно.
Перед скачиванием данного файла вспомните о тех хороших рефератах, контрольных, курсовых, дипломных работах, статьях и других документах, которые лежат невостребованными в вашем компьютере. Это ваш труд, он должен участвовать в развитии общества и приносить пользу людям. Найдите эти работы и отправьте в базу знаний.
Мы и все студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будем вам очень благодарны.
Чтобы скачать архив с документом, в поле, расположенное ниже, впишите пятизначное число и нажмите кнопку "Скачать архив"
| Рубрика | Биология и естествознание |
| Вид | учебное пособие |
| Язык | русский |
| Дата добавления | 10.03.2014 |
| Размер файла | 52,6 M |
Сведения о нуклеиновых кислотах, история их открытия и распространение в природе. Строение нуклеиновых кислот, номенклатура нуклеотидов. Функции нуклеиновых кислот (дезоксирибонуклеиновая - ДНК, рибонуклеиновая - РНК). Первичная и вторичная структура ДНК.
реферат [1,8 M], добавлен 26.11.2014
История изучения нуклеиновых кислот как биополимеров, мономерами которых являются нуклеотиды, функции и значение в жизнедеятельности организма. Правила Чаргаффа. Первичная и вторичная структура ДНК. Особенности репликации у эукариот, ее разновидности.
презентация [533,6 K], добавлен 05.11.2014
Молекулярная организация генетического материала. Транскрипция и трансляция мРНК прокариот. Роль рибонуклеиновых кислот в белковом синтезе. Расположение функциональных центров на субчастицах рибосомы. Свойства генетического кода. Активация аминокислот.
курсовая работа [2,0 M], добавлен 19.11.2013
Свойства вирусов, особенности их строения и классификация. Взаимодействие вируса с клеткой. Процессы, связанные с размножением вируса. Описание основных вирусных заболеваний. Эволюция вирусов на современном этапе. Влияние загрязнения внешней среды.
реферат [466,4 K], добавлен 24.03.2011
Понятие генетического кода как единой системы записи наследственной информации в молекулах нуклеиновых кислот в виде последовательности нуклеотидов. Этапы реализации, свойства и расшифровка хромосомы в клетке. Работа по секвенсированию генома человека.
реферат [89,1 K], добавлен 18.01.2011
Эволюционное происхождение. Свойства вирусов. Природа вирусов. Строение и классификация вирусов. Взаимодействие вируса с клеткой. Значение вирусов. Вирусные заболевания. Особенности эволюции вирусо на соременном этапе.
реферат [299,2 K], добавлен 22.11.2005
Первичная, вторичная и третичная структуры ДНК. Свойства генетического кода. История открытия нуклеиновых кислот, их биохимические и физико-химические свойства. Матричная, рибосомальная, транспортная РНК. Процесс репликации, транскрипции и трансляции.
реферат [4,1 M], добавлен 19.05.2015
Сущность, состав нуклеотидов, их физические характеристики. Механизм редупликации дезоксирибонуклеиновой кислоты (ДНК), транскрипция ее с переносом наследственной информации на РНК и механизм трансляции — синтез белка, направляемый этой информацией.
реферат [461,8 K], добавлен 11.12.2009
Латенция и вирогения как типы взаимодействия вируса с клеткой. Процесс адсорбции вируса и его проникновения в клетку, синтез вирусных белков. Этапы созревания дочерних вирусных частиц, способы их выхода из клетки, общие принципы сборки вирионов.
реферат [18,6 K], добавлен 29.09.2009
Особенности применения метода ядерного магнитного резонанса (ЯМР) для исследования нуклеиновых кислот, полисахаридов и липидов. Исследование методом ЯМР комплексов нуклеиновых кислот с протеинами и биологических мембран. Состав и структура полисахаридов.
курсовая работа [3,5 M], добавлен 26.08.2009

Вирусы — это микроскопические патогены, заражающие клетки живых организмов для самовоспроизводства. Они состоят из одного вида нуклеиновой кислоты (или ДНК или РНК, но не обе вместе), которая защищена оболочкой, содержащей белки, липиды, углеводы или их комбинацию. Размер типичного вируса варьируется от 15 до 350 нм, поэтому его можно увидеть только с помощью электронного микроскопа.
В 1892 году русский ученый Д.И. Ивановский впервые доказал существование ранее неизвестного типа возбудителя болезней, это был вирус мозаичной болезни табака. А в 1898 году Фридрих Лоффлер и Пол Фрош нашли доказательства того, что причиной ящура у домашнего скота была инфекционная частица, которая меньше, чем любая бактерия. Это были первые шаги к изучению природы вирусов, генетических образований, которые лежат где-то в серой зоне между живыми и неживыми состояниями материи. На текущий момент описано около 6 тыс. вирусов, но их существует несколько миллионов.
Строение вирусов
Вне клеток-хозяев вирусы существуют в виде белковой оболочки (капсида), иногда заключенного в белково-липидную мембрану. Капсид обволакивает собой либо ДНК, либо РНК, которая кодирует элементы вируса. Находясь в такой форме вне клетки, вирус метаболически инертен и называется вирионом.
Простая структура, отсутствие органелл и собственного метаболизма позволяет некоторым вирусам кристаллизоваться, т.е. они могут вести себя подобно химическим веществам. С появлением электронных микроскопов было установлено, что их кристаллы состоят из тесно прижатых друг к другу нескольких сотен миллиардов частиц. В одном кристалле вируса полиомиелита столько частиц, что ими можно заразить не по одному разу всех жителей Земли.
Формы вирусов
Вирусы встречаются в трех основных формах. Они бывают:
- Сферическими (кубическими или полигидральными). Вирусы герпеса, типулы, полиомы и т.д.
- Спиральными (цилиндрическими или стержнеобразными). Вирусы табачной мозаики, гриппа, эпидемического паротита и др.
- Сложными. Например, бактериофаги.
Проникновение вирусов в клетку-хозяина
Капсид в основном защищает нуклеиновую кислоту от действия клеточного нуклеазного фермента. Но некоторые белки капсида способствуют связыванию вируса с поверхностью клеток-хозяев, и работают, как ключики, вставляемые в нужные замочки. Другие поверхностные белки действуют как ферменты, они растворяют поверхностный слой клетки-хозяина и таким образом помогают проникновению нуклеиновой кислоты вируса в клетку-хозяина.
Жизненный цикл вирусов сильно отличается у разных видов, но существует шесть основных этапов жизненного цикла вирусов:
Присоединение к клетке-хозяину представляет собой специфическое связывание между вирусными капсидными белками и рецепторами на клеточной поверхности. Эта специфика определяет хозяина вируса.
Проникновение следует за прикреплением: вирионы проникают в клетку-хозяина через рецептор-опосредованный эндоцитоз или слияние мембран. Это часто называют вирусной записью.
Проникновение вирусов в клетку достигается за счет:
Размножение вирусов
После того, как вирусный геном освобождается от капсида, начинается его транскрипция или трансляция. Именно эта стадия вирусной репликации сильно различается между ДНК- и РНК-вирусами и вирусами с противоположной полярностью нуклеиновой кислоты. Этот процесс завершается синтезом новых вирусных белков и генома (точных копий внедрённых).
Механизм репликации зависит от вирусного генома.
- ДНК-вирусы обычно используют белки и ферменты клетки-хозяина для получения дополнительной ДНК, она транскрибируется в РНК-мессенджер (мРНК), которая затем используется для управления синтезом белка.
- РНК-вирусы обычно используют ядро РНК в качестве матрицы для синтеза вирусной геномной РНК и мРНК. Вирусная мРНК направляет клетку-хозяина на синтез вирусных ферментов и капсидных белков и сборку новых вирионов. Конечно, есть исключения из этого шаблона. Если клетка-хозяин не обеспечивает ферменты, необходимые для репликации вируса, вирусные гены предоставляют информацию для прямого синтеза отсутствующих белков.
Чтобы преобразовать РНК в ДНК, вирусы должны содержать гены, которые кодируют вирус-специфический фермент обратной транскриптазы. Она транскрибирует матрицу РНК в ДНК. Обратная транскрипция никогда не происходит в неинфицированных клетках. Необходимый фермент, обратная транскриптаза, происходит только от экспрессии вирусных генов в инфицированных клетках.
Вироиды
Вироиды заражают только растения. Одни вызывают экономически важные заболевания сельскохозяйственных культур, в то время как другие являются доброкачественными. Двумя примерами экономически важных вироидов являются кокосный cadang-cadang (он вызывает массовую гибель кокосовых пальм) и вироид рубцовой кожицы яблок, который безнадежно портит товарный вид яблок.
30 известных вироидов были классифицированы в две семьи.
- Члены семейства Pospiviroidae, названные по имени вироида клубневого веретена картофеля, имеют палочковидную вторичную структуру с небольшими одноцепочечными областями, имеет центральную консервативную область, и реплицируются в ядре клетки.
- Avsunviroidae, названный в честь вироида авокадо, имеет как палочковидную, так и разветвленную области, но не имеет центральной консервативной области и реплицируется в хлоропластах растительной клетки.
В отличие от вирусов, которые являются паразитами механизма трансляции хозяина, вироиды являются паразитами клеточных транскрипционных белков.
Бактериофаги

Существуют тысячи разновидностей фагов, каждый из которых может заразить только один тип или несколько близких типов бактерий или архей. Фаги классифицируются по ряду семейств вирусов; например:
Как и все вирусы, фаги являются простыми организмами, которые состоят из ядра генетического материала (нуклеиновой кислоты), окруженного капсидом белка. Нуклеиновая кислота может представлять собой либо ДНК, либо РНК, и может быть двухцепочечной или одноцепочечной.
Существует три основных структурных формы фага:
- Икосаэдрическая (20-сторонняя) головка с хвостом
- Икосаэдрическая головка без хвоста
- Нитевидная форма
Во время заражения фаг прикрепляется к бактерии и вставляет в нее свой генетический материал. После этого фаг обычно следует одному из двух жизненных циклов: литическому (вирулентному) или лизогенному (умеренному).
Литические, или вирулентные, фаги захватывают механизм клетки, чтобы скопировать компоненты фага. Затем они разрушают или лизируют клетку, высвобождая новые частицы фага.
Лизогенные, или умеренные, фаги включают свою нуклеиновую кислоту в хромосому клетки-хозяина и реплицируются с ней как единое целое, не разрушая клетку. При определенных условиях лизогенные фаги могут индуцироваться в соответствии с литическим циклом.
Существуют и другие жизненные циклы, в т.ч. псевдолизогенез и хроническая инфекция. При псевдолизогении бактериофаг проникает в клетку, но не использует механизм репликации клеток и не интегрируется в геном хозяина, просто как бы прячется внутри бактерии, не нанося ей никакого вреда. Псевдолизогенез возникает, когда клетка-хозяин сталкивается с неблагоприятными условиями роста и, по-видимому, играет важную роль в выживании фага, обеспечивая сохранение генома фага до тех пор, пока условия роста хозяина снова не станут благоприятными.
При хронической инфекции новые фаговые частицы образуются непрерывно и длительно, но без явного уничтожения клеток.
Вскоре после открытия фаги начали использовать для лечения бактериальных заболеваний человека, таких как бубонная чума и холера. Но фаговая терапия тогда не была успешной, и после открытия антибиотиков в 1940-х годах она была практически заброшена. Однако с появлением устойчивых к антибиотикам бактерий терапевтическому потенциалу фагов уделяется все больше внимания.
Наше время с антибиотиками заканчивается. В 2016 году женщина в штате Невада умерла от бактериальной инфекции, вызванной Klebsiella pneumoniae, которая была устойчивой ко всем известным антибиотикам. Бактерии, устойчивые к колистину, антибиотику последней инстанции, были обнаружены на свинофермах в Китае. В настоящее время бактерии приспосабливаются к антибиотикам быстрее, чем когда-либо.
Покажите ножницы которыми вирусы разрезают молекулу РНК что бы встроиться для мутации.Может что нибудь придумаете другое.К примеру деление цепочка аминокислот получив энергию из вне как одноименные заряды распадается на две. К каждой соединятся только те какие были ранее (другие проскочат мимо),казалось бы копии,но внутренняя энергия разная(уменьшается увеличивается) поэтому распад и создание. Вся химия углерода на этом построена 1000 орган соединений создает у других хим элементов этого свойства нет. Иммунная система делает накладку(интерференция)с помощью энергии интерферонов пытаясь разрушить цепочку РНК вируса.Надо помочь организму но не вакциной(вирус быстро мутирует)
Геном вирусов может быть представлен одноцепочечными или двухцепочечными ДНК или РНК. Гены вирусов могут быть заключены в одной хромосоме или разделены на несколько блоков (хромосом) , которые все вместе и составляют геном таких вирусов. Например, у ретвирусов геном представлен двухцепочечной РНК и состоит из десяти сегментов. Геномы вирусов, содержащих одноцепочечную РНК, также могут быть либо цельными (у ретровирусов ), либо сегментированными (у аренавирусов ). Геном РНК-содержащих вирусов представлен только линейными молекулами РНК. Все известные ДНК- содержащие вирусы позвоночных имеют геном, заключенный в одной хромосоме, линейной или кольцевой, одно- или двухцепочечной.Репликация геномов вирусов ДНК-геномы реплицируются клеточными или вирусоспецифическими ДНК-полимеразами. РНК-геномы реплицируются вирусоспецифическими РНК-полимеразами, которые также являются и транскриптазами. Репликация вирусных геномов происходит или одновременно с транскрипцией, или эти два процесса разделены во времени. Механизмы репликации геномов вирусов многообразны и определяются видом генома. Существует три модели репликации – полуконсервативная, консервативная и дисперсная. Консервативная и дисперсная модели репликации нуклеиновых кислот установлены только у вирусов. Полуконсервативная модель предполагает, что после первого раунда репликации одна цепь в каждой из двух дочерних молекул является родительской, другая – синтезируемой заново. По такой схеме реплицируются двунитевыеДНК-геномы вирусов. При реализации консервативной модели репликации одна дочерняя молекула состоит из двух родительских цепей, а другая – из вновь синтезированных цепей. Согласно консервативной модели реплицируются двунитевые РНК ротавирусов .ДНК-содержащие вирусы, реплицирующиеся таким образом, неизвестны. Дисперсная модель репликации приводит к образованию молекулы нуклеиновой кислоты, состоящей из фрагментов, как родительских цепей, так и вновь синтезированных.
Строение геномов эукариот и особенности их репликации
Геном эукариотДля клеток эукариот характерно наличие оформленного ядра. Информационной макромолекулой их генома является ДНК, которая неравномерно распределена по нескольким хромосомам в виде комплексов с многочисленными белками. Жизненно важная генетическая информация заключена и во внехромосомных молекулах ДНК. У эукариот — это ДНК хлоропластов, митохондрий и других пластид. Геном эукариот обладает избыточностью. Эукариотическая клетка содержит во много раз больше генов, чем прокариотическая. Большая часть их геномной ДНК представлена некодирующими последовательностями нуклеотидов. У эукариот около 5% ДНК составляют экзоны (участки гена, кодирующие белок), 25% – интроны (участки гена, которые транскрибируются, но затем удаляются при сплайсинге), а остальные 70% составляют спейсеры – нетранскрибируемые участки ДНК между генами.Особенности репликации у эукариот.Инициация репликации. Репликация начинается на определённом сайте молекулы ДНК, точке начала репликации или ori-сайте. В этой точке двойная спираль ДНК расплетается с образованием двух репликативных вилок, которые движутся по направлению друг к другу .В формировании репликативных вилок участвуют ДНК-топоизоимеразы и ДНК-хеликазы. По окончанию формирования репликативной вилки фермент ликвидирует разрыв и отделяется от ДНК.Разрыв водородных связей в двуцепочечной молекуле ДНК осуществляет фермент ДНК-хеликаза. Для этого фермент использует энергию макроэргических связей АТФ. В результате происходит раскручивание суперпсиральтной структуры ДНК в поддержании такой структуры участвуют SSB-белки. Эти белки не закрывают азотистых оснований и не препятствуют репликации.Элонгация – продолжение репликации. Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами.Субстартами для синтеза новой цепи ДНК являются дезоксинуклеозидтрифосфаты.. Эти же соединения служат источниками энергии. На образование каждой связи расходуются две макроэргические связи. В процессе также участвуют ионы магния, нейтрализуя отрицательный заряд нуклеотидов.У эукариот принимают участие 5 ДНК полимераз (α, β, γ, δ, ε), которые различаются по числу субъединиц, молекулярной массе, способности взаимодействовать с другими белками и функциональному назначению. Полимеразы β, δ и ε не могут начинать образование дочерней ДНК, т.к. не имеют сродства к одиночной цепи ДНК. Начинает репликацию полимераза α, которая синтезирует небольшой фрагмент РНК-праймер и далее примерно 60 нуклеотидных остатков ДНК. Образующийся небольшой двухцепочечный фрагмент позволяет присоединиться полимеразе δ и продолжить синтез новой цепи в направлении от 5/ к 3/. Выбор очередного нуклеотида определяется матрицей ДНК. При созревании отстающей цепи ДНК праймеры удаляют ДНК-полимераза β и присоединяет недостающие нуклеотиды. Восстановленные фрагменты соединяют ДНК-лигазы.
Вирусы - это внеклеточная форма жизни, обладающая собственным геномом и способная к воспроизведению только в клетках живых организмов.
Вирион (или вирусная частица) состоит из одной или нескольких молекул ДНК или РНК, заключенных в белковую оболочку (капсид), иногда содержащую также липидные и углеводные компоненты.
Вирусы размножаются только после инфицирования живых клеток. Различные вирусы проникают в животные и растительные клетки, а также бактерии (вирусы бактерий называются бактериофагами). Вирусы являются внутриклеточными паразитами на генетическом уровне и используют для своего размножения белок-синтезирующий аппарат клетки-хозяина.
По сравнению с геномами других организмов вирусный геном относительно мал и кодирует лишь ограниченное число белков, в основном белки капсида и один или несколько белков, участвующих в репликации и экспрессии вирусного генома. Необходимые метаболиты и энергия поставляются хозяйской клеткой.
Геном вирусов, заключенный внутри вирионов, может быть представлен ДНК или РНК, последние могут быть одно - и двуцепочечными, кольцевыми и линейными. Значения молекулярных масс ДНК вирусов находится в пределах 1∙10 6 - 200∙10 6 ; РНК - 1∙10 6 - 15∙10 6 дальтон.
По сложности строения генома вирусы широко варьируют - от фага Qβ (РНК-содержащий вирус бактерий), имеющего 4 гена, до вируса оспы (ДНК-содержащий вирус), геном которого насчитывает около 250 генов. Кроме того, все гены вируса могут быть заключены в одной молекуле нуклеиновой кислоты или распределены по нескольким молекулам, которые вместе и составляют геном такого вируса.
Типы генетического материала и механизм его репликации
У различных вирусов
3.1 РНК-содержащие вирусы (РНК → РНК)
Геномы почти всех известных РНК-содержащих вирусов, реплицирующихся по данной схеме - это линейные молекулы, их удобно разделить на 2 группы.
Первая группа -это однонитевые геномы положительнойполярности, т.е. с нуклеотидной последовательностью, соответствующей таковой у мРНК.

Такие геномы обозначают как (+) РНК. Цепь РНК такого типа может сразу после проникновения в клетку-хозяина обеспечивать синтез вирусных белков. Однонитевый (+) РНК-геном характерен для фага Qβ, вирусов табачной мозаики, полиомиелита, клещевого энцефалита. Вирусные (+) РНК-геномы кодируют несколько белков, среди которых РНК-зависимая РНК-полимераза (репликаза),способная синтезировать молекулы РНК без участия ДНК. С помощью этого фермента синтезируются сначала (-) нити РНК вируса, а затем репликаза осуществляет синтез (+) нити РНК. На заключительной стадии из накопившихся вирусных белков и (+) РНК формируются новые вирионы.
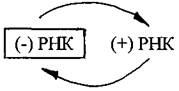
Этот вирусный фермент РНК-зависимая РНК-полимераза,синтезированная в предыдущем цикле размножения,упакован в вирионе в удобной для доставки в клетку форме. Инфекционный процесс начинается с того, что вирусный фермент копирует вирусный геном, образуя (+) РНК, которая выступает в качестве матрицы для синтеза вирусных белков, в том числе РНК-зависимой РНК-полимеразы, которая входит в состав образующихся вирионов.
ДНК-содержащие вирусы
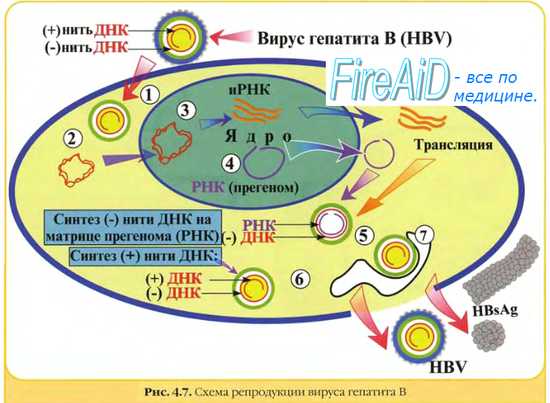
Первая группа- вирусы с двуцепочечной ДНК, репликация которых осуществляется по схеме: ДНК→РНК→ДНК. Они получили название ретроидные вирусы. Представителями этой группы вирусов являются вирус гепатита В и вирус мозайки цветной капусты.
Репликация ДНК-генома этих вирусов осуществляется при посредстве промежуточных молекул РНК:
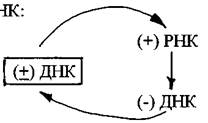
Молекулы (+) РНК образуются в результате транскрипциивирусных ДНК в клеточном ядре хозяйским ферментом ДНК-зависимой РНК-полимеразой.Транскрибируется только одна из нитей вирусной ДНК. Дальнейший синтез ДНК на РНК-матрице происходит в результате реакции, катализируемой обратной транскриптазой;сначала синтезируется (-) нить ДНК, а затем на вновь синтезированной (-) нити ДНК тот же фермент строит (+) нить. (+) РНК также используется для трансляции белков, необходимых для сборки вирусных частиц.
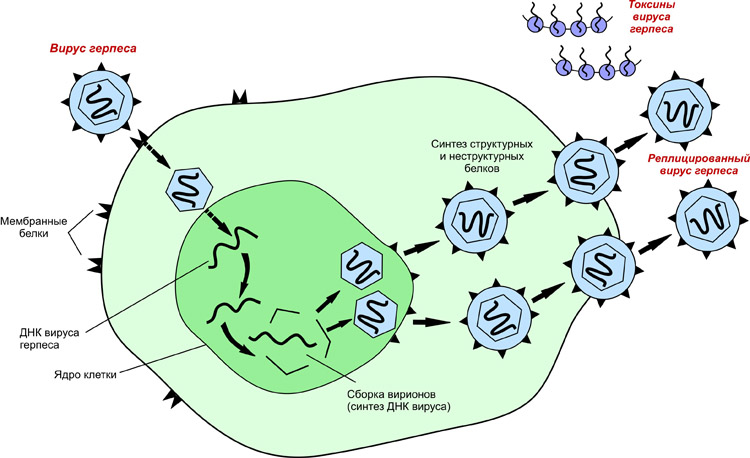
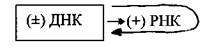
Вторая группа- вирусы с двухцепочечной ДНК, репликация которых осуществляется по схеме ДНК→ДНК. К этой группе относятся вирусы герпеса, оспы и др. В зараженной клетке ДНК-зависимая РНК-полимераза(фермент клетки-хозяина) транскрибирует с генома этих вирусов молекулы мРНК (т.е. (+) РНК), которые принимают участие в синтезе вирусных белков, а размножение вирусного генома осуществляет фермент ДНК-зависимая ДНК-полимераза:
Бактериофаги
Отмеченные особенности характерны для вирусов животных, растений и бактерий. Последние, называемые бактериофагами, наиболее детально изучены с генетической точки зрения. Связано это, прежде всего с легкостью их культивирования в лабораторных условиях.
Разные фаги обнаруживают специфичность действия - они действуют на бактерии лишь определенного вида, редко на близкие виды и часто - на отдельные штаммы данного вида. Фаги со специфическим приспособлением к тому или иному хозяину отличаются от других фагов морфологией, размерами, биохимическими и серологическими свойствами.
Биохимический анализ выявляет в фаговой частице два компонента -белок и нуклеиновую кислоту. Белок образует хвост фага и мембрану головки. При резком изменении осмотической силы среды мембрана может разрываться и ДНК освобождается.
Практически все известные фаги содержат ДНК. Молекулы у них состоят из двух комплементарных нитей. Каждая фаговая частица содержит лишь одну молекулу ДНК.
Поскольку размножение фагов происходит внутри бактериальной клетки, рассмотрим типичную картину взаимодействия фага и чувствительной бактерии.
Фаговые частицы адсорбируются на поверхности бактерии, прикрепляясь к ее оболочке своими хвостами. Зараженная бактерия через 15-45 мин лизируется, освобождая 100 и более зрелых частиц следующего поколения фага. Встречая чувствительные бактериальные клетки, они вновь заражают их, и весь цикл повторяется. Если размножение фага происходит в культуре в жидкой среде, то оно приводит к просветлению (из-за лизиса клеток) суспензии бактерий. Размножение фага на бактериях, растущих на плотной среде, выявляется в виде зон лизиса среди сплошного газона бактерий. Эти пятна обнаженного агара называются негативными колониями фага.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.
Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности.
Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
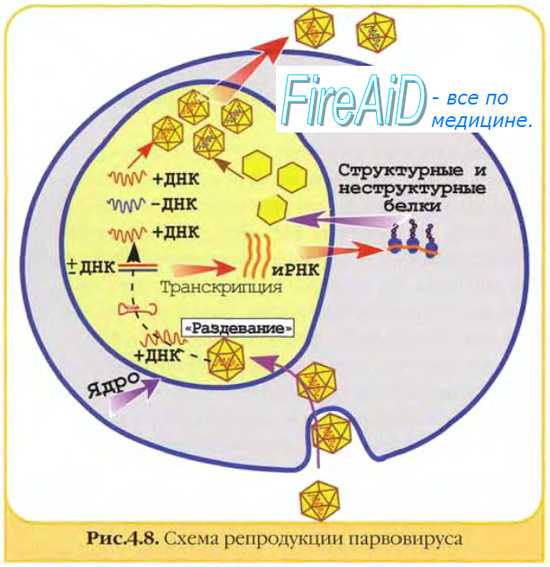
Матричные белкинеобходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе).
Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова- и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
начальный период включает стадии адсорбции вируса на клетке, проникновения в клетку, дезинтеграции (депротеинизации) или "раздевания" вируса. Вирусная нуклеиновая кислота была доставлена в соответствующие клеточные структуры и под действием лизосомальных ферментов клетки освобождается от защитных белковых оболочек. В итоге формируется уникальная биологическая структура: инфицированная клетка содержит 2 генома (собственный и вирусный) и 1 синтетический аппарат (клеточный);
• после этого начинается вторая группа процессов репродукции вируса, включающая средний и заключительный периоды, во время которых происходят репрессия клеточного и экспрессия вирусного генома. Репрессию клеточного генома обеспечивают низкомолекулярные регуляторные белки типа гистонов, синтезируемые в любой клетке. При вирусной инфекции этот процесс усиливается, теперь клетка представляет собой структуру, в которой генетический аппарат представлен вирусным геномом, а синтетический аппарат — синтетическими системами клетки.
2. Дальнейшее течение событий в клетке направлено на репликацию вирусной нуклеиновой кислоты(синтез генетического материала для новых вирионов) и реализацию содержащейся в ней генетической информации (синтез белковых компонентов для новых вирионов). У ДНК-содержащих вирусов, как в прокариотиче-ских, так и в эукариотических клетках, репликация вирусной ДНК происходит при участии клеточной ДНК-зависимой ДНК-полимеразы. При этом у однонитевых ДНК-содержащих вирусов сначала образуется комплементарная нить — так называемая репликативная форма, которая служит матрицей для дочерних молекул ДНК.
3. Реализация генетической информации вируса, содержащейся в ДНК, происходит следующим образом: при участии ДНК-зависимой РНК-полимеразы синтезируются и-РНК, которые поступают на рибосомы клетки, где и синтезируются вирусспе-цифические белки. У двунитевых ДНК-содержащих вирусов, геном которых транскрибируется в цитоплазме клетки хозяина, это собственный геномный белок. Вирусы, геномы которых транскрибируются в ядре клетки, используют содержащуюся там клеточную ДНК-зависимую РНК-полимеразу.
У РНК-содержащих вирусов процессы репликацииих генома, транскрипции и трансляции генетической информации осуществляются иными путями. Репликация вирусных РНК, как минус-, так и плюс-нитей, осуществляется через репликативную форму РНК (комплементарную исходной), синтез которой обеспечивает РНК-зависимая РНК-полимераза — это геномный белок, который есть у всех РНК-содержащих вирусов. Репликативная форма РНК минус-нитевых вирусов (плюс-нить) служит не только матрицей для синтеза дочерних молекул вирусной РНК (минус-нитей), но и выполняет функции и-РНК, т. е. идет на рибосомы и обеспечивает синтез вирусных белков (трансляцию).
У плюс-нитевыхРНК-содержащих вирусов функцию трансляции выполняют ее копии, синтез которых осуществляется через репликативную форму (минус-нить) при участии вирусных РНК-зависимых РНК-полимераз.
У некоторых РНК-содержащих вирусов (реовирусы) имеется совершенно уникальный механизм транскрипции. Он обеспечивается специфическим вирусным ферментом — ревертазой (обратной транскриптазой) и называется обратной транскрипцией. Суть ее состоит в том, что вначале на матрице вирусной РНК при участии обратной транскрипции образуется транскрипт, представляющий собой одну нить ДНК. На нем с помощью клеточной ДНК-зависимой ДНК-полимеразы синтезируется ,вторая нить и формируется двунитевой ДНК-транскрипт. С него обычным путем через образование и-РНК происходит реализация информации вирусного генома.
Результатом описанных процессов репликации, транскрипции и трансляции является образование дочерних молекул вирусной нуклеиновой кислоты и вирусных белков, закодированных в геноме вируса.
После этого наступает третий, заключительный период взаимодействия вируса и клетки. Из структурных компонентов (нуклеиновых кислот и белков) на мембранах цитоплазматического ретикулума клетки собираются новые вирионы. Клетка, геном которой был репрессирован (подавлен), обычно гибнет. Вновь сформировавшиеся вирионы пассивно (в результате гибели клетки) или активно(путем почкования) покидают клетку и оказываются в окружающей ее среде.
Таким образом, синтез вирусных нуклеиновых кислот и белков и сборка новых вирионов происходят в определенной последовательности (разобщены во времени) и в разных структурах клетки (разобщен в пространстве), в связи с чем способ репродукции вирусов и был назван дизъюнктивным (разобщенным). При абортивной вирусной инфекции процесс взаимодействия вируса с клеткой по тем или иным причинам прерывается до того, как произошло подавление клеточного генома. Очевидно, что в этом случае генетическая информация вируса реализована не будет и репродукции вируса не происходит, а клетка сохраняет свои функции неизменными.
При латентной вирусной инфекции в клетке одновременно функционируют оба генома, а при вирус-индуцированных трансформациях вирусный геном становится частью клеточного, функционирует и наследуется вместе с ним.
Дата добавления: 2018-05-09 ; просмотров: 462 ;
Читайте также: