
Транскрипция и трансляция у вирусов
Измененные вирусные моноцистронные мРНК связываются с рибосомами и транслируются в белки в основном с 5'-конца так же, как клеточные мРНК. У некоторых вирусов транскрибируется полицистронная мРНК, которая может транслироваться прямо в несколько генных продуктов. Имеется несколько механизмов перекрывания рамок считывания, что значительно расширяет кодирующий потенциал вирусов.
Это особенно важно для вирусов с небольшим геномом и еще раз указывает на экономичность экспрессии вирусных геномов, кодирующих сложные функции минимальными нуклеотидными последовательностями. Примером сказанному могут служить лентивирусы, содержащие всего 9 генов, но обладающие сложным контролем транскрипции и циклом репликации, а также удовлетворяющими требованиями установления латентной инфекции.
Большинство вирусных белков претерпевают посттрансляционные изменения, такие как: фосфолирование (для связывания нуклеиновой кислоты), ациллирование жирных кислот (для включения в мембрану), гликозилирование, миристалирование или протеолитическое расщепление.
Вирусы используют клеточный метаболизм для синтеза боковых цепей гликопротеинов вирусной оболочки.
Белки, кодируемые вирусными геномами
| 1. Структурные белки вирионов (капсидные, коровые, оболочечные) 2. Ферменты, связанные с вирионами, особенно полимеразы (транскриптазы) и др. 3. Неструктурные белки, главным образом ферменты, необходимые для транскрипции, репликации вирусной нуклеиновой кислоты и расщепления белков 4. Регуляторные белки, контролирующие временную последовательность экспрессии вирусного генома 5. Белки, понижающие экспрессию клеточных генов (транскрипцию, иногда трансляцию) 6. Онкогенные (продукты) белки и инактиваторы клеточных белков, подавляющих трансформацию клеток (герпес-, адено-, папиллома-, полиома- и ретровирусные инфекции) 7. Белки, влияющие на вирулентность вируса, хозяинный спектр, тканевой тропизм и т.д. (наиболее сложные ДНК-вирусы) 8. Вирокины, модифицирующие неинфицированные клетки, тем самым способствуя распространению инфекции в организме |
Посттрансляционное расщепление вирусных белков имеет место у многих вирусов, у которых полицистронная вирусная (+)РНК транслируется непосредственно в один полипротеин, который несет протеазную активность, расщепляющую полипротеин в определенных местах на мелкие белки. Первое расщепление происходит, когда полипротеин еще связан с рибосомой. Некоторые большие фрагменты существуют только быстротечно, тогда как другие функционируют короткий период и в последующем расщепляются дополнительно протеазами, кодируемыми вирусом, на мелкие белки с различными функциями.
Некоторые вирусы кодируют несколько различных протеаз, большинство из которых является трипсиноподобными. Клеточные протеазы присутствуют в таких органеллах как аппарат Гольджи или транспортные вакуоли, которые принимают участие в сборке и созревании многих вирусов. Например, расщепление гемагглютинина ортомиксовирусов или гликопротеина сплавления (F белок) парамиксовирусов имеет важное значение для инфекционности вирионов.
В таблице приведены белки, кодируемые вирусными геномами. В общем, белки, транслируемые с ранних транскриптов ДНК-вирусов, включают ферменты и другие белки, необходимые для репликации вирусных нуклеиновых кислот белков, подавляющих синтез клеточных РНК и белков. Крупные ДНК-вирусы (оспо- и герпесвирусы) также кодируют многочисленные ферменты, связанные с метаболизмом нуклеотидов. Поздние вирусные белки транслируются на поздних мРНК, большинство из которых транскрибируется на молекулах потомства вирусных нуклеиновых кислот. Большинство поздних белков являются вирусными структурными белками и часто образуются в избыточном количестве.
Некоторые вирусные белки обладают другими важными функциями. К ним относятся регуляторные белки, изменяющие транскрипцию или трансляцию клеточных генов или ранних вирусных генов. Крупные ДНК-вирусы также кодируют многие дополнительные белки, называемые вирокинами (подобие клеточных цитокинов), которые не регулируют цикл вирусной репликации, как таковой, но влияют на ответ хозяина на инфекцию.
Лекция 5
ТРАНСКРИПЦИЯ
Транскрипция — это переписывание ДНК на РНК по законам генетического кода. Это означает, что РНК состоит из нуклеотидных последовательностей, комплементарных ДНК. Нити ДНК в участке транскрипции разделяются и функционируют как матрицы, к которым присоединяются комплементарные нуклеотиды благодаря спариванию комплементарных оснований (аденин связывается с тимином, урацил — с аденином, гуанин — с цитозином и цитозин - с гуанином). Транскрипция осуществляется с помощью специального фермента — РНК-полимеразы, который связывает нуклеотиды путем образования 3'-5'-фосфодиэфирных мостиков. Такое связывание происходит лишь в присутствии ДНК-матрицы.
Продуктами транскрипции в клетке являются иРНК. Сама клеточная ДНК, являющаяся носителем генетической информации, не может непосредственно программировать синтез белка. Передачу генетической информации от ДНК к рибосомам осуществляет РНК-посредник. На этом основана центральная догма молекулярной биологии, которая выражается следующей схемой:
ДНК - (транскрипция) –и РНК – (трансляция) - белок
где стрелки показывают направление переноса генетической информации.
Реализация генетической информации у вирусов. Стратегия вирусного генома в отношении синтеза иРНК у разных вирусов различна. У ДНК-содержащих вирусов иРНК синтезируется на матрице одной из нитей ДНК. Формула переноса генетической информации у них такая же, как и в клетке.
ДНК-содержащие вирусы, репродукция которых происходит в ядре, используют для транскрипции клеточную полимеразу. К этим вирусам относятся паповавирусы, аденовирусы, вирусы герпеса. ДНК-содержащие вирусы, репродукция которых происходит в цитоплазме, не могут использовать клеточный фермент, находящийся в ядре. Транскрипция их генома осуществляется вирусспецифическим ферментом — ДНК-полимеразой, которая проникает в клетку в составе вируса. К этим вирусам относятся вирусы оспы и иридовирусы.
К этой группе вирусов относятся пикорнавирусы, тогавирусы, коронавирусы. У них нет необходимости в акте транскрипции для синтеза вируспецифических белков. Поэтому транскрипцию как самостоятельный процесс у этих вирусов не выделяют. Иначе обстоит дело у вирусов, геном которых не может выполнять функцию иРНК. В клетке синтезируется комплементарная геному РНК, которая и является информационной. Передача генетической информации у этих вирусов осуществляется по схеме:
РНК– иPHK - белок
У этих вирусов транскрипция выделена как самостоятельный процесс в инфекционном цикле. К ним относятся две группы вирусов животных.
2. Вирусы, геном которых представлен двунитчатой РНК (диплорнавирусы). Среди вирусов животных к ним относятся реовирусы.
В клетке нет фермента, который может полимеризовать нуклеотиды на матрице РНК. Эту функцию выполняет вирусспецифический фермент — РНК-полимераза, или транскриптаза, которая находится в составе вирусов и вместе с ними проникает в клетку.
Среди РНК-содержащих вирусов животных есть семейство ретровирусов, которые имеют уникальный путь передачи генетической информации. РНК этих вирусов переписывается на ДНК, ДНК интегрирует с клеточным геномом и в его составе переписывается на РНК, которая обладает информационными функциями. Путь передачи генетической информации в-этом случае осуществляется по более сложной схеме:
РНК - ДНК - иРНК - белок
Ферменты, транскрибирующие вирусный геном. Транскрипция ряда ДНК-содержащих вирусов — паповавирусов, аденовирусов, вирусов герпеса, парвовирусов, гепадна-вирусов осуществляется в ядре клетки, и в этом процессе широко используются механизмы клеточной транскрипции — ферменты транскрипции и дальнейшей модификации транскриптов. Транскрипция этих вирусов осуществляется клеточной РНК-полимеразой II — ферментом, который осуществляет транскрипцию клеточного генома. Однако особая группа транскриптов аденовируса синтезируется с помощью другого клеточного фермента — РНК-полимеразы III. У двух других семейств ДНК-содержащих вирусов животных - вирусов оспы и иридовирусов — транскрипция происходит в цитоплазме. Поскольку в цитоплазме нет клеточных полимераз, транскрипция этих вирусов нуждается в специальном вирусном ферменте — вирусной РНК-полимеразе. Этот фермент является структурным вирусным белком.
У РНК-содержащих вирусов транскрипция осуществляется вирусспецифическими транскриптазами, т. е. ферментами, закодированными в вирусном геноме. Вирусспецифические транскриптазы могут быть как структурными белками, входящими в состав вириона (эндогенная транскриптаза), так и неструктурными белками, которые синтезируются в зараженной клетке, но не включаются в вирион.
Транскрипция в зараженной клетке. Синтез комплементарных РНК на родительских матрицах с помощью родительской транскриптазы носит название первичной транскрипции в отличие от вторичной транскрипции, происходящей на более поздних стадиях инфекционного цикла на вновь синтезированных, дочерних матрицах, с помощью вновь синтезированной транскриптазы. Большая часть иРНК в зараженной клетке является продуктом вторичной транскрипции.
Транскриптивные комплексы. У сложно устроенных РНК-содержащих вирусов животных транскрипция происходит не на матрице голой РНК, а в составе вирусных нуклеокапсидов или сердцевин (транскриптивные комплексы). Связанные с геномом капсидные белки не только не препятствуют транскрипции, но и необходимы для нее, обеспечивая правильную конформацию тяжа РНК, защиту его от клеточных протеаз, связь отдельных фрагментов генома друг с другом, а также регуляцию транскрипции.
Регуляция транскрипции. Транскрипция вирусного генома строго регулируется на протяжении инфекционного цикла. Регуляция осуществляется как клеточными, так и вирусспецифическими механизмами. У некоторых вирусов, в основном ДНК-содержащих, существует три периода транскрипций — сверхранняя, ранняя и поздняя. К этим вирусам относятся вирусы оспы, герпеса, паповавирусы, аденовирусы. В результате сверхранней и ранней транскрипции избирательно считываются сверхранние и ранние гены с образованием сверхранних или ранних иРНК. При поздней транскрипции считывается другая часть вирусного генома — поздние гены, с образованием поздних иРНК. Количество поздних генов обычно превышает количество ранних генов. Многие сверхранние гены являются генами для неструктурных белков — ферментов и регуляторов транскрипции и репликации вирусного генома. Напротив, поздние гены обычно являются генами для структурных белков. Обычно при поздней транскрипции считывается весь геном, но с преобладанием транскрипции поздних генов.
Фактором регуляции транскрипции у ядерных вирусов является транспорт транскриптов из ядра в цитоплазму, к месту функционирования иРНК — полисомам.
У РНК-содержащих вирусов синтез транскриптов также строго контролируется в отношении как количества каждого класса транскриптов, так и периода инфекции, когда определенные транскрипты синтезируются с максимальной скоростью. На ранней стадии инфекции преимущественно синтезируются транскрипты двух генов вируса гриппа — NP и NS, на поздней стадии инфекции — транскрипты генов М, НА и NA. Остальные три гена для Р-белков синтезируются примерно с одинаковой скоростью на протяжении всего периода инфекции. У реовирусов на ранней стадии инфекции преимущественно транскрибируется 4 из 10 фрагментов генома и лишь на поздней стадии транскрибируется весь геном. Однако если поместить геном вируса в бесклеточную РНК-синтезирующую систему, будет происходить равномерная транскрипция всех 10 фрагментов генома. Эти факты говорят о жестком контроле транскрипции со стороны клетки-хозяина и возможном наличии специфических клеточных регуляторов.
ТРАНСЛЯЦИЯ
Синтез белка в клетке происходит в результате трансляции иРНК. Трансляцией называется процесс перевода генетической информации, содержащейся в иРНК, на специфическую последовательность аминокислот. Иными словами, в процессе трансляции осуществляется перевод 4-буквенного языка азотистых оснований на 20-буквенный язык аминокислот.
Рибосомы. Синтез белка в клетке осуществляется на рибосоме. Рибосома состоит из двух субъединиц, большой и малой, малая субъединица примерно в два раза меньше большой. Обе субъединицы содержат по одной молекуле рибосомальной РНК и ряд белков. Рибосомальные РНК синтезируются в ядре на матрице ДНК с помощью РНК-полимеразы. В малой рибосомальной субъединице есть канал, в котором находится информационная РНК. В большой рибосомальной субъединице есть две полости, захватывающие также малую рибосомальную субъединицу. Одна из них содержит аминоацильный центр (А-центр), другая — пептидильный центр (П-центр).
Фазы трансляции. Процесс трансляции состоит из трех фаз: 1) инициации, 2) элонгации и 3) терминации.
Вначале с иРНК связывается малая рибосомальная субъединица. К комплексу иРНК с малой рибосомальной субъединицей присоединяются другие компоненты, необходимые для начала трансляции. Их по крайней мере три в прокариотической клетке и более девяти в эукариотической клетке. Инициаторные факторы определяют узнавание рибосомой специфических иРНК и, таким образом, являются определяющим фактором в дискриминации между различными иРНК, присутствующими в клетке, как правило, в избыточном количестве.
В результате формируется комплекс, необходимый для инициации трансляции, который называется инициаторным комплексом. В инициаторный комплекс входят: 1) иРНК; 2) малая рибосомальная субъединица; 3) аминоацил-тРНК, несущая инициаторную аминокислоту; 4) инициаторные факторы; 5) несколько молекул ГТФ.
В рибосоме осуществляется слияние потока информации с потоком аминокислот. Аминоацил-тРНК входит в А-центр большой рибосомальной субъединицы, и ее антикодон взаимодействует с кодоном иРНК, находящейся в малой рибосомальной субъединице. При продвижении иРНК на один кодон тРНК перебрасывается в пептидильный центр, и ее аминокислота присоединяется к инициаторной аминокислоте с образованием первой пептидной связи. Свободная от аминокислоты тРНК выходит из рибосомы и может опять функционировать в транспорте специфических аминокислот.
Терминация трансляции. Терминация трансляции происходит в тот момент, когда рибосома доходит до терминирующего кодона в составе иРНК. Трансляция прекращается, и полипептидная цепь освобождается из полирибосомы. После окончания трансляции полирибосомы распадаются на субьединицы, которые могут войти в состав новых полирибосом.
И транскрипция, и трансляция относятся к матричным биосинтезам. Матричным биосинтезом называется синтез биополимеров (нуклеиновых кислот, белков) на матрице - нуклеиновой кислоте ДНК или РНК. Процессы матричного биосинтеза относятся к пластическому обмену: клетка расходует энергию АТФ.
Матричный синтез можно представить как создание копии исходной информации на несколько другом или новом "генетическом языке". Скоро вы все поймете - мы научимся достраивать по одной цепи ДНК другую, переводить РНК в ДНК и наоборот, синтезировать белок с иРНК на рибосоме. В данной статье вас ждут подробные примеры решения задач, генетический словарик пригодится - перерисуйте его себе :)

Возьмем 3 абстрактных нуклеотида ДНК (триплет) - АТЦ. На иРНК этим нуклеотидам будут соответствовать - УАГ (кодон иРНК). тРНК, комплементарная иРНК, будет иметь запись - АУЦ (антикодон тРНК). Три нуклеотида в зависимости от своего расположения будут называться по-разному: триплет, кодон и антикодон. Обратите на это особое внимание.
Процесс синтеза дочерней молекулы ДНК по матрице родительской ДНК. Нуклеотиды достраивает фермент ДНК-полимераза по принципу комплементарности. Переводя действия данного фермента на наш язык, он следует следующему правилу: А (аденин) переводит в Т (тимин), Г (гуанин) - в Ц (цитозин).
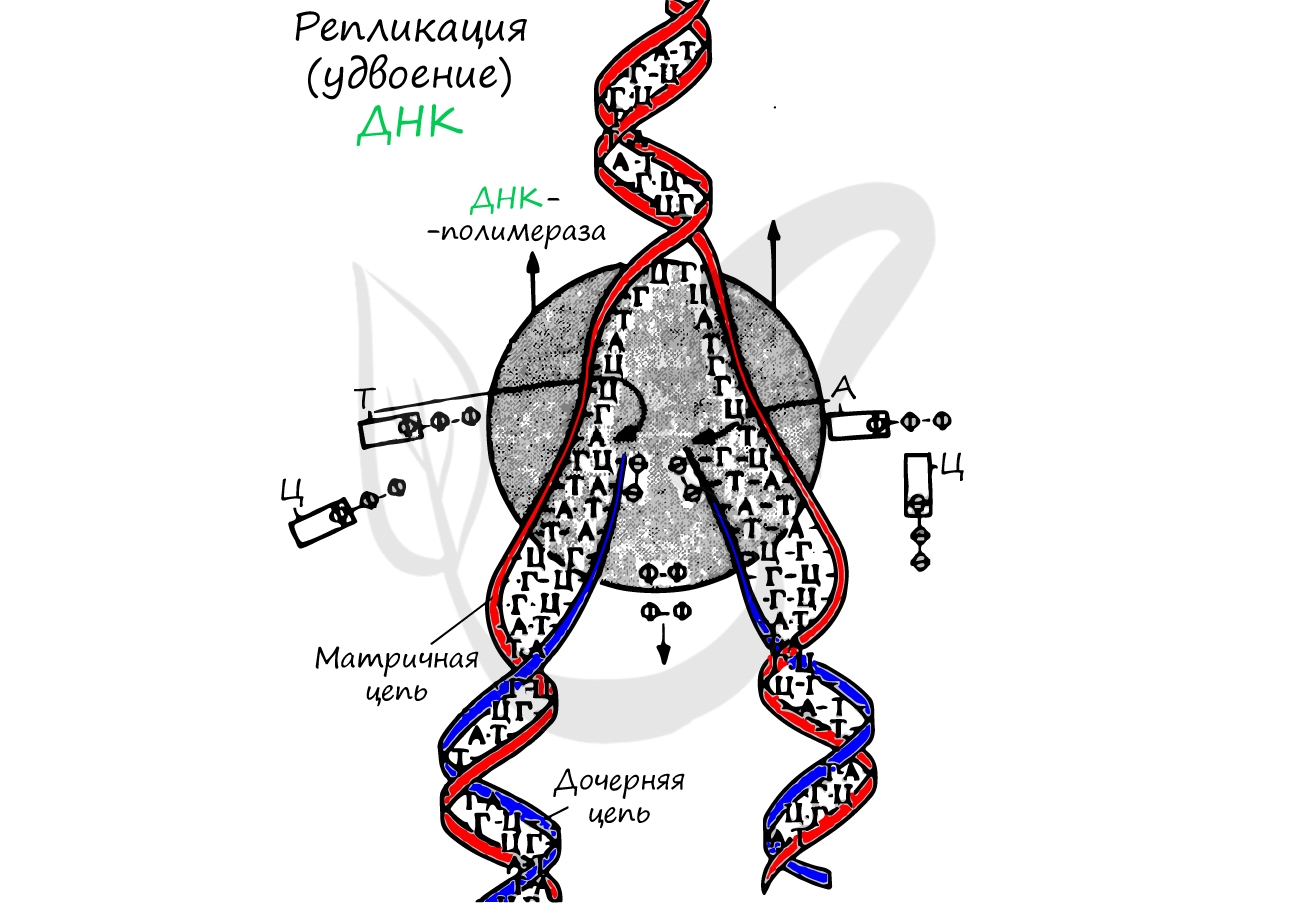
Удвоение ДНК происходит в синтетическом периоде интерфазы. При этом общее число хромосом не меняется, однако каждая из них содержит к началу деления две молекулы ДНК: это необходимо для равномерного распределения генетического материала между дочерними клетками.
Транскрипция представляет собой синтез информационной РНК (иРНК) по матрице ДНК. Несомненно, транскрипция происходит в соответствии с принципом комплементарности азотистых оснований: А - У, Т - А, Г - Ц, Ц - Г (загляните в "генетический словарик" выше).
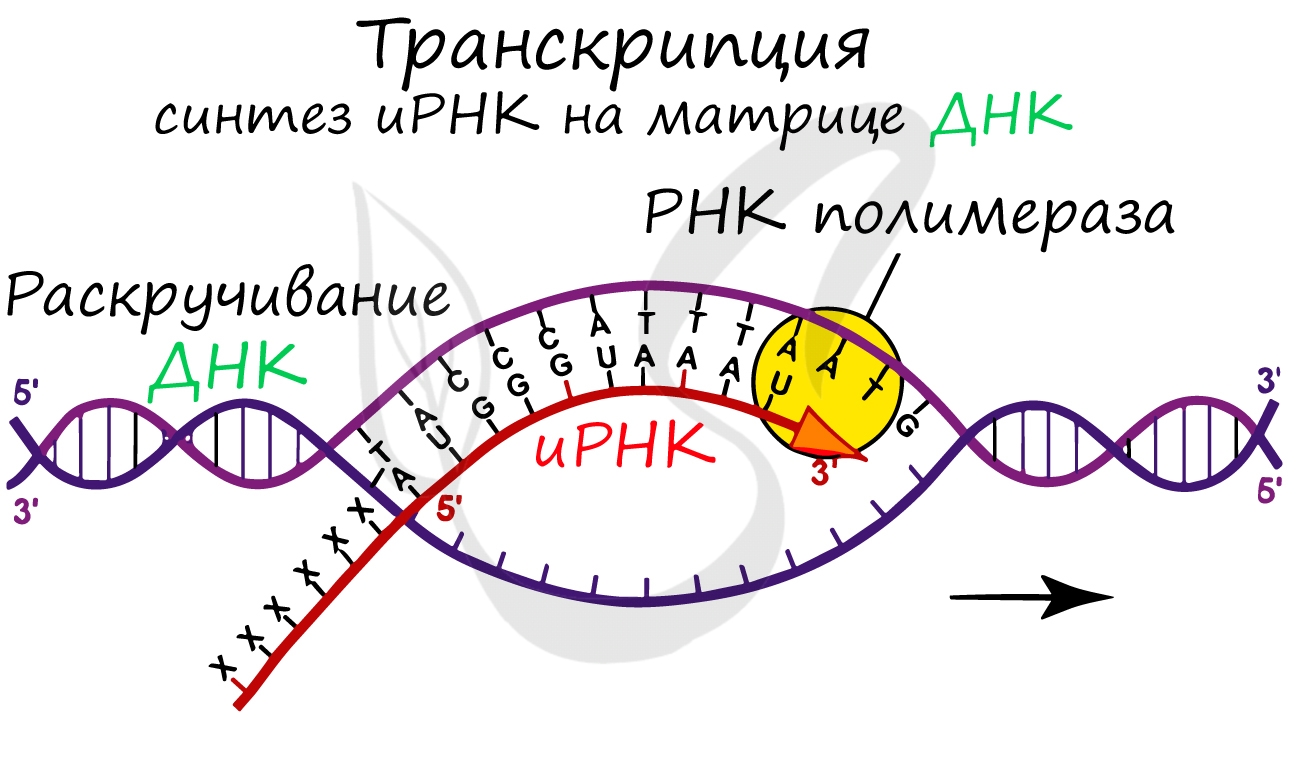
До начала непосредственно транскрипции происходит подготовительный этап: фермент РНК-полимераза узнает особый участок молекулы ДНК - промотор и связывается с ним. После связывания с промотором происходит раскручивание молекулы ДНК, состоящей из двух цепей: транскрибируемой и смысловой. В процессе транскрипции принимает участие только транскрибируемая цепь ДНК.
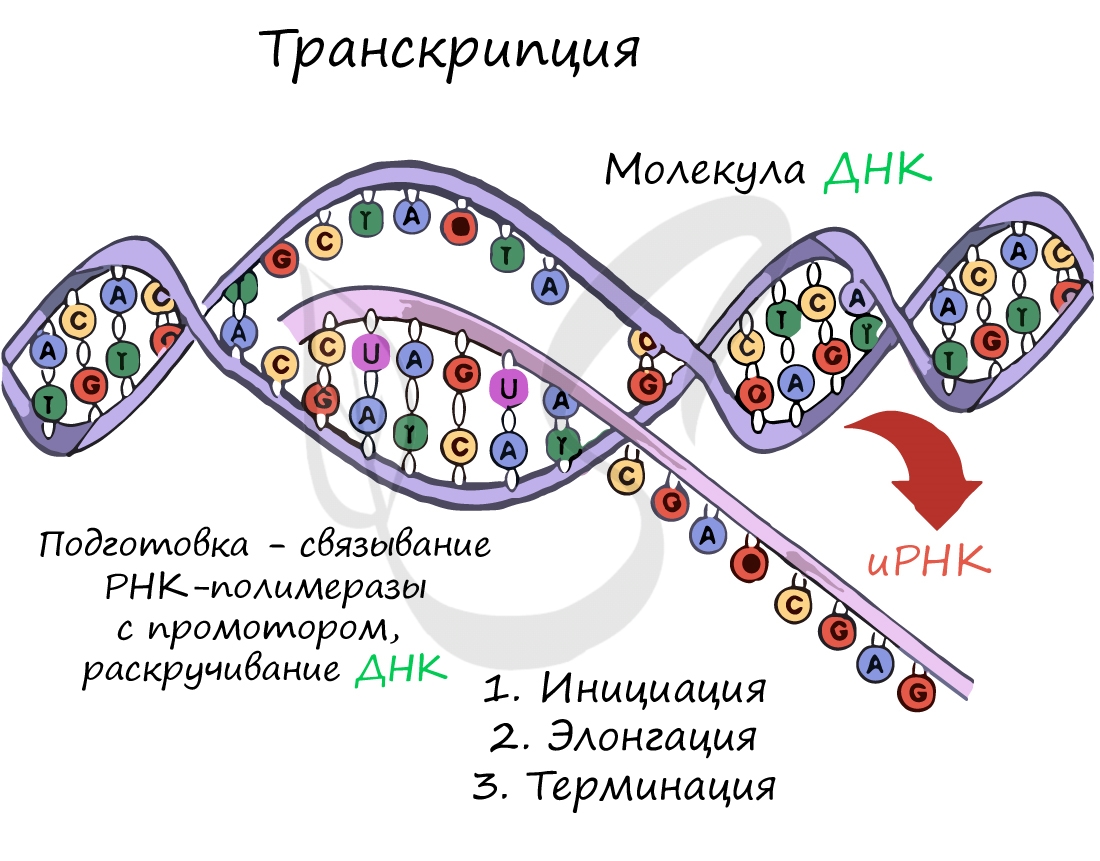
Транскрипция осуществляется в несколько этапов:
-
Инициация (лат. injicere — вызывать)
Образуется несколько начальных кодонов иРНК.
Элонгация (лат. elongare — удлинять)
Нити ДНК последовательно расплетаются, освобождая место для передвигающейся РНК-полимеразы. Молекула иРНК быстро растет.
Терминация (лат. terminalis — заключительный)
Достигая особого участка цепи ДНК - терминатора, РНК-полимераза получает сигнал к прекращению синтеза иРНК. Транскрипция завершается. Синтезированная иРНК направляется из ядра в цитоплазму.
Куда же отправляется новосинтезированная иРНК в процессе транскрипции? На следующую ступень - в процесс трансляции. Он заключается в синтезе белка на рибосоме по матрице иРНК. Последовательность кодонов иРНК переводится в последовательность аминокислот.
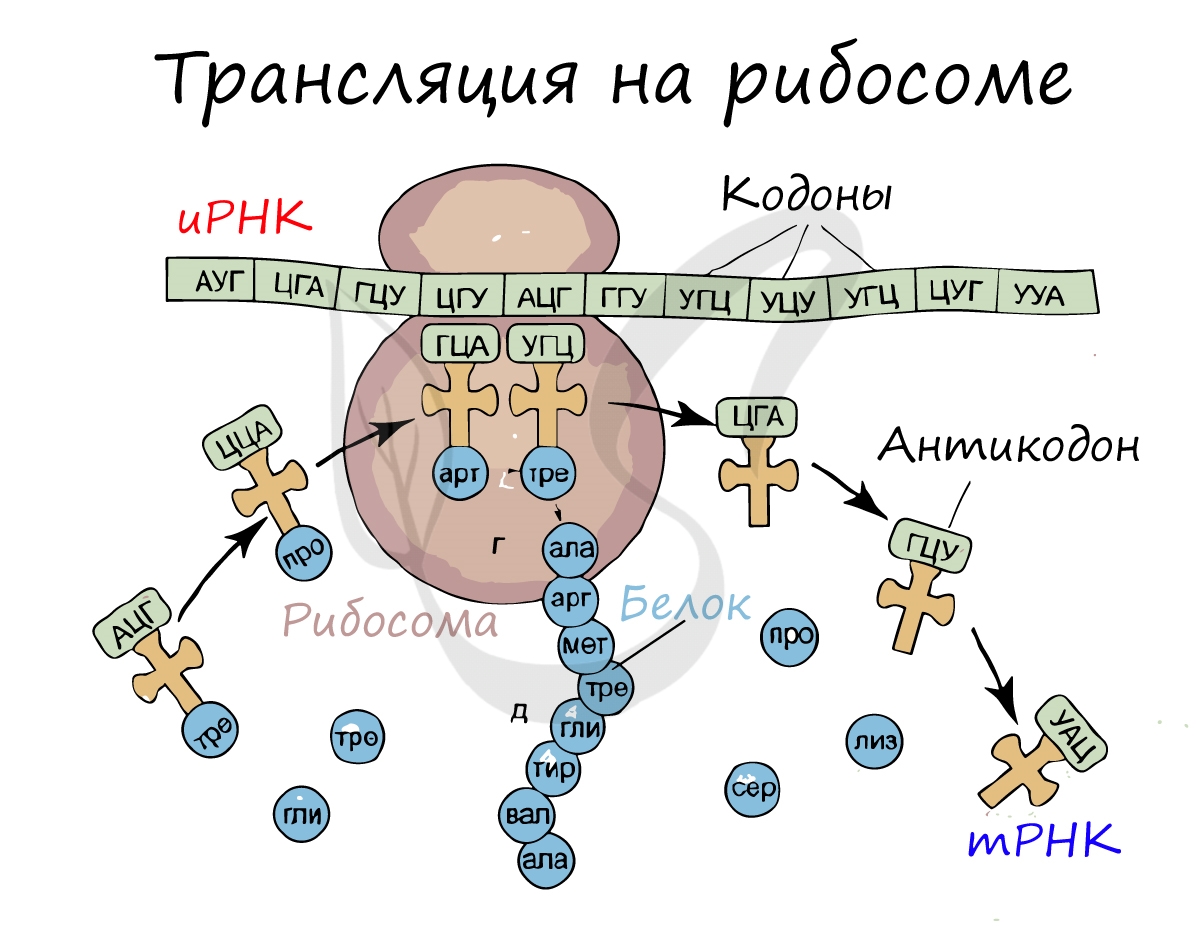
Перед процессом трансляции происходит подготовительный этап, на котором аминокислоты присоединяются к соответствующим молекулам тРНК. Трансляцию можно разделить на несколько стадий:
-
Инициация
Информационная РНК (иРНК, синоним - мРНК (матричная РНК)) присоединяется к рибосоме, состоящей из двух субъединиц. Замечу, что вне процесса трансляции субъединицы рибосом находятся в разобранном состоянии.
Первый кодон иРНК, старт-кодон, АУГ оказывается в центре рибосомы, после чего тРНК приносит аминокислоту, соответствующую кодону АУГ - метионин.
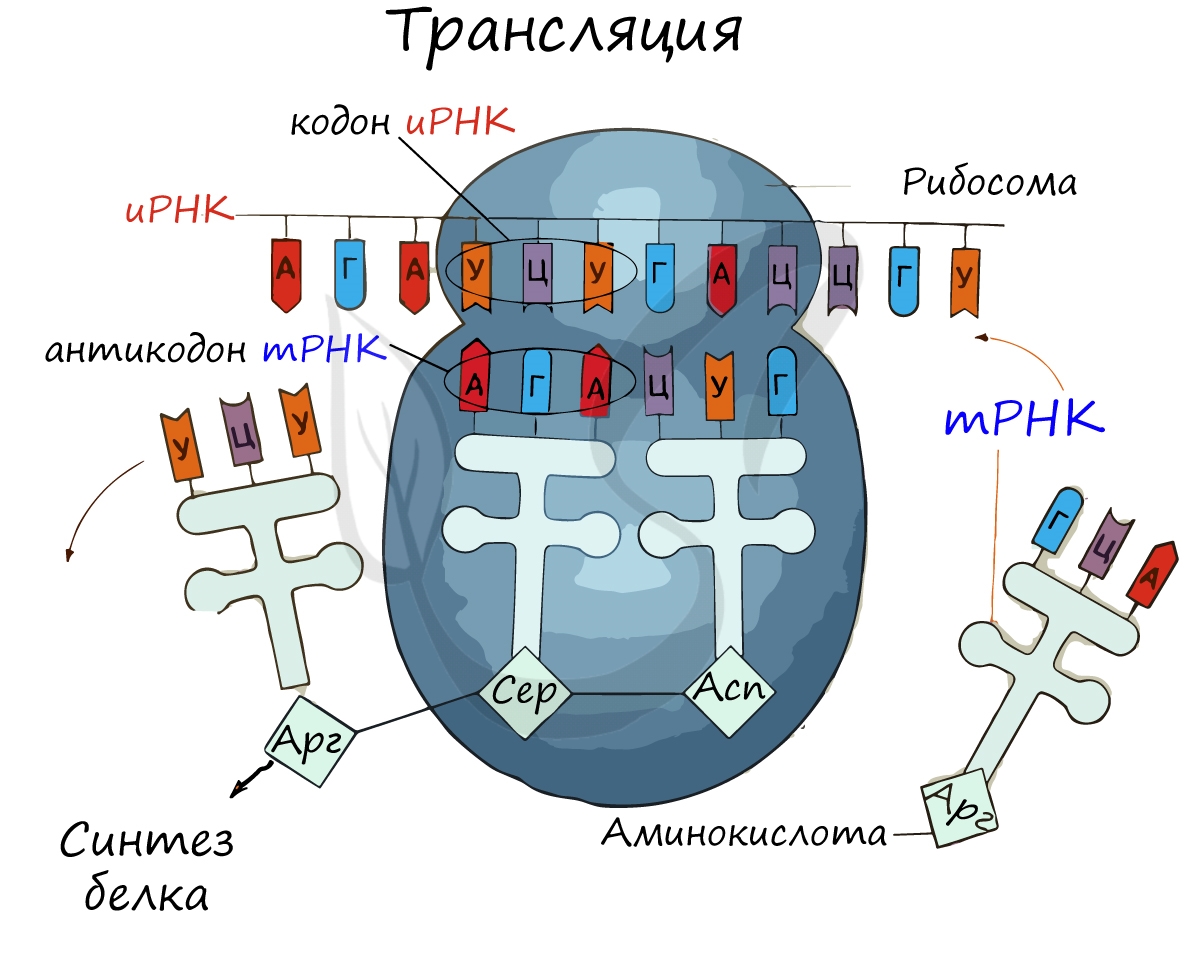
Рибосома делает шаг, и иРНК продвигается на один кодон: такое в фазу элонгации происходит десятки тысяч раз. Молекулы тРНК приносят новые аминокислоты, соответствующие кодонам иРНК. Аминокислоты соединяются друг с другом: между ними образуются пептидные связи, молекула белка растет.
Доставка нужных аминокислот осуществляется благодаря точному соответствию 3 нуклеотидов (кодона) иРНК 3 нуклеотидам (антикодону) тРНК. Язык перевода между иРНК и тРНК выглядит как: А (аденин) - У (урацил), Г (гуанин) - Ц (цитозин). В основе этого также лежит принцип комплементарности.
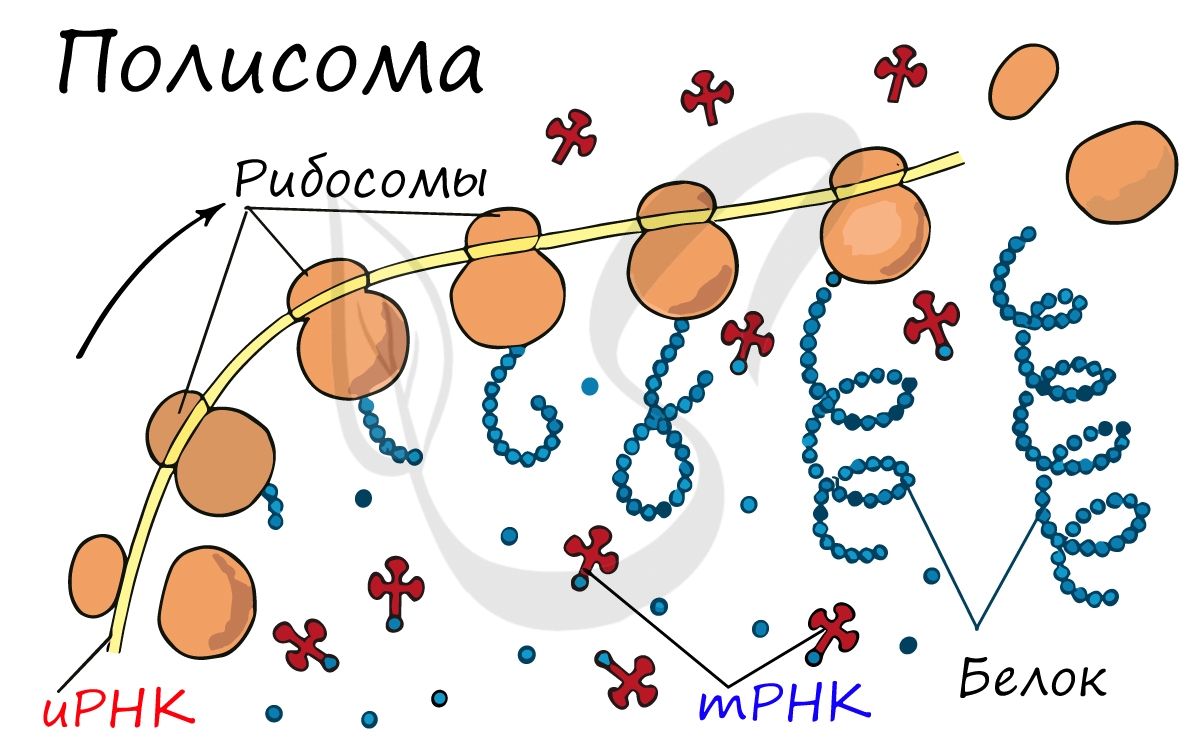
Движение рибосомы вдоль молекулы иРНК называется транслокация. Нередко в клетке множество рибосом садятся на одну молекулу иРНК одновременно - образующаяся при этом структура называется полирибосома (полисома). В результате происходит одновременный синтез множества одинаковых белков.
Синтез белка - полипептидной цепи из аминокислот - в определенный момент завершатся. Сигналом к этому служит попадание в центр рибосомы одного из так называемых стоп-кодонов: УАГ, УГА, УАА. Они относятся к нонсенс-кодонам (бессмысленным), которые не кодируют ни одну аминокислоту. Их функция - завершить синтез белка.
Существует специальная таблица для перевода кодонов иРНК в аминокислоты. Пользоваться ей очень просто, если вы запомните, что кодон состоит из 3 нуклеотидов. Первый нуклеотид берется из левого вертикального столбика, второй - из верхнего горизонтального, третий - из правого вертикального столбика. На пересечении всех линий, идущих от них, и находится нужная вам аминокислота :)
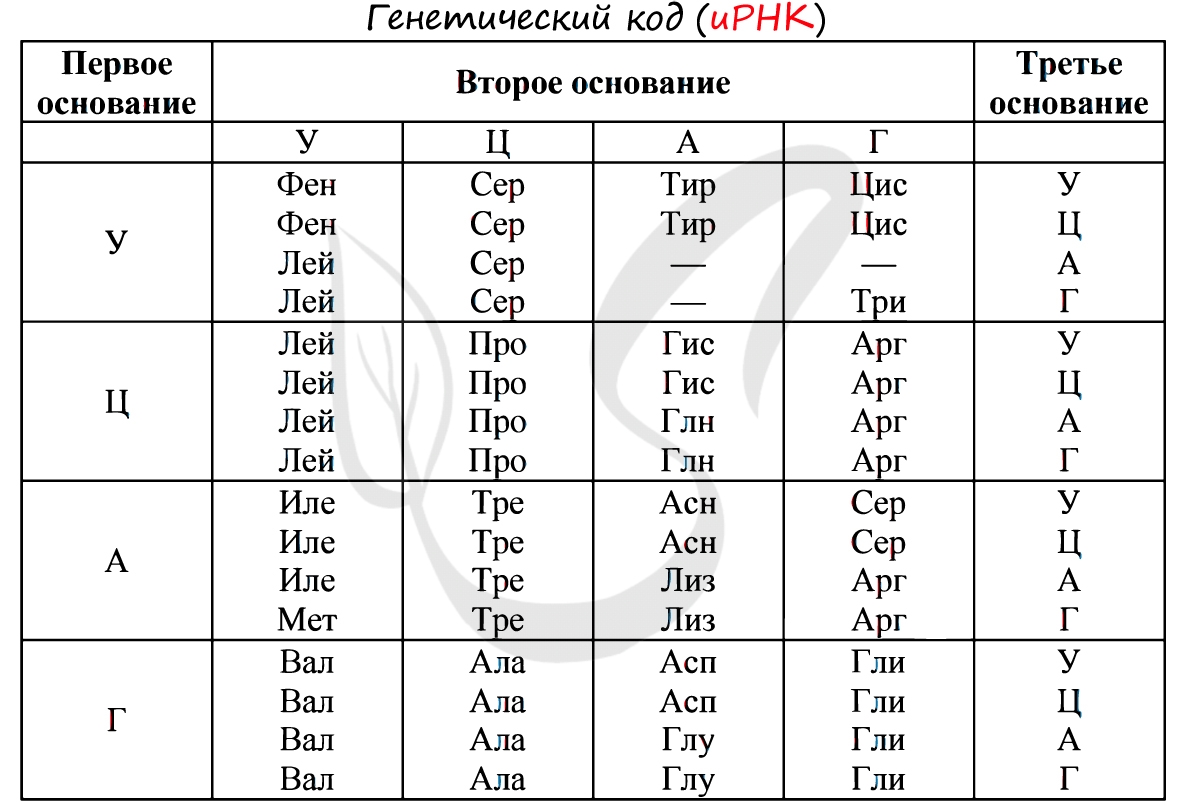
Давайте потренируемся: кодону ЦАЦ соответствует аминокислота Гис, кодону ЦАА - Глн. Попробуйте самостоятельно найти аминокислоты, которые кодируют кодоны ГЦУ, ААА, УАА.
Кодону ГЦУ соответствует аминокислота - Ала, ААА - Лиз. Напротив кодона УАА в таблице вы должны были обнаружить прочерк: это один из трех нонсенс-кодонов, завершающих синтез белка.
Без практики теория мертва, так что скорее решим задачи! В первых двух задачах будем пользоваться таблицей генетического кода (по иРНК), приведенной вверху.
"Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ЦГА-ТГГ-ТЦЦ-ГАЦ. Определите последовательность нуклеотидов во второй цепочке ДНК, последовательность нуклеотидов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка, используя таблицу генетического кода"

По принципу комплементарности мы нашли вторую цепочку ДНК: ГЦТ-АЦЦ-АГГ-ЦТГ. Мы использовали следующие правила при нахождении второй нити ДНК: А-Т, Т-А, Г-Ц, Ц-Г.
Вернемся к первой цепочке, и именно от нее пойдем к иРНК: ГЦУ-АЦЦ-АГГ-ЦУГ. Мы использовали следующие правила при переводе ДНК в иРНК: А-У, Т-А, Г-Ц, Ц-Г.
Зная последовательность нуклеотидов иРНК, легко найдем тРНК: ЦГА, УГГ, УЦЦ, ГАЦ. Мы использовали следующие правила перевода иРНК в тРНК: А-У, У-А, Г-Ц, Ц-Г. Обратите внимание, что антикодоны тРНК мы разделяем запятыми, в отличие кодонов иРНК. Это связано с тем, что тРНК представляют собой отдельные молекулы (в виде клеверного листа), а не линейную структуру (как ДНК, иРНК).
"Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК, имеет следующую последовательность нуклеотидов: ТАГ-ЦАА-АЦГ-ГЦТ-АЦЦ. Установите нуклеотидную последовательность участка тРНК, который синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта тРНК в процессе биосинтеза белка, если третий триплет соответствует антикодону тРНК"

Обратите свое пристальное внимание на слова "Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК ". Эта фраза кардинально меняет ход решения задачи: мы получаем право напрямую и сразу синтезировать с ДНК фрагмент тРНК - другой подход здесь будет считаться ошибкой.
Итак, синтезируем напрямую с ДНК фрагмент молекулы тРНК: АУЦ-ГУУ-УГЦ-ЦГА-УГГ. Это не отдельные молекулы тРНК (как было в предыдущей задаче), поэтому не следует разделять их запятой - мы записываем их линейно через тире.
Третий триплет ДНК - АЦГ соответствует антикодону тРНК - УГЦ. Однако мы пользуемся таблицей генетического кода по иРНК, так что переведем антикодон тРНК - УГЦ в кодон иРНК - АЦГ. Теперь очевидно, что аминокислота кодируемая АЦГ - Тре.
Длина фрагмента молекулы ДНК составляет 150 нуклеотидов. Найдите число триплетов ДНК, кодонов иРНК, антикодонов тРНК и аминокислот, соответствующих данному фрагменту. Известно, что аденин составляет 20% в данном фрагменте (двухцепочечной молекуле ДНК), найдите содержание в процентах остальных нуклеотидов.

Один триплет ДНК состоит из 3 нуклеотидов, следовательно, 150 нуклеотидов составляют 50 триплетов ДНК (150 / 3). Каждый триплет ДНК соответствует одному кодону иРНК, который в свою очередь соответствует одному антикодону тРНК - так что их тоже по 50.
По правилу Чаргаффа: количество аденина = количеству тимина, цитозина = гуанина. Аденина 20%, значит и тимина также 20%. 100% - (20%+20%) = 60% - столько приходится на оставшиеся цитозин и гуанин. Поскольку их процент содержания равен, то на каждый приходится по 30%.
Теперь мы украсили теорию практикой. Что может быть лучше при изучении новой темы? :)
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
После слияния вирусной оболочки с клеточной мембраной нуклеокапсид оказывается в клетке, и вирионная транскриптаза начинает транскрипцию геномной РНК с образованием индивидуальных вирусных мРНК. Для этого не требуется устранения белка NP : в отличие от позитивно-нитевых вирусов , полного "раздевания" вирусной РНК не происходит, что не мешает транскриптазе осуществлять транскрипцию геномной РНК. Транскриптаза следует от З'-конца геномной РНК к 5'-концу, последовательно транскрибируя вирусные гены. Синтез каждой мРНК завершается на терминирующем участке соответствующего гена, и транскриптаза, пройдя межгенный участок, начинает транскрипцию следующего гена. Синтезированные мРНК транслируются , и происходит накопление вирусных белков. Когда их концентрация достигает определенного уровня, белок NP начинает связываться с синтезируемой РНК, что служит для транскриптазы сигналом к изменению характера ее работы. Транскриптаза перестает распознавать межгенные участки, и вместо отдельных коротких мРНК синтезируется полноразмерная плюс-цепь РНК, полностью комплементарная геномной минус-цепи. По ходу синтеза она связывается с белком NP, образуя структуру, сходную с нуклеокапсидом, но содержащую РНК позитивной полярности. Эта РНК, в свою очередь, используется вирусной транскриптазой в качестве матрицы для синтеза новых геномных РНК, которые включаются в состав нуклеокапсидов и либо подвергаются транскрипции, либо используются для новых актов репликации.
Новосинтезированные белки вирусной оболочки попадают в плазматическую мембрану и образуют в ней участки, к которым транспортируются содержащие геномную РНК нуклеокапсиды, после чего происходит почкование вируса и выход вирусных частиц из клетки. Процессы транскрипции и репликации у вирусов семейств Rhabdoviridae , Paramyxoviridae и Filoviridae проходят в цитоплазме, а у Bornaviridae - в ядре. В последнем случае часть молекул мРНК подвергается сплайсингу , что увеличивает разнообразие вирусных белков за счет сдвига рамки трансляции. У Paramyxoviridae и Filoviridae увеличение объема генетической информации достигается за счет использования при трансляции разных инициирующих кодонов , не совпадающих по фазе считывания, а также за счет "проскальзывания" транскриптазы на определенных участках при синтезе мРНК, в результате чего в полинуклеотидную цепь включаются дополнительные нуклеотиды и рамка трансляции мРНК оказывается сдвинутой.
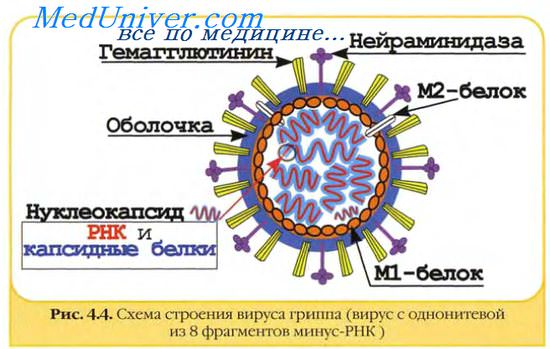
У некоторых семейств негативно-нитевых вирусов РНК представлена не единой молекулой, а несколькими сегментами РНК. Количество сегментов может составлять от 3 (у вирусов семейства Bunyaviridae ) до 8 (у некоторых вирусов семейства Orthomyxoviridae ). Каждый сегмент кодирует либо один вирусный белок, как большинство генов у представителей Orthomyxoviridae, либо два, либо даже несколько белков. У вирусов семейства Orthomyxoviridae транскрипция и репликация осуществляются в клеточном ядре и возможность кодирования двух белков одним и тем же геном обеспечивается сплайсингом. Например, у вируса гриппа A некоторые молекулы мРНК, транскрибированной с гена М , подвергаются сплайсингу, в результате которого трансляция мРНК после первых 9 триплетов идет в иной фазе, нежели трансляция тех мРНК, которые не подверглись сплайсингу. При трансляции мРНК, которая не подверглась сплайсингу, образуется белок M1 , а при трансляции сплайсированной мРНК - белок М2 . У вирусов семейства Bunyaviridae , репликация и транскрипция которых осуществляются в цитоплазме, сплайсинг, естественно, отсутствует, но трансляция в разных рамках считывания, тем не менее, имеет место. В этом случае в мРНК имеется два инициирующих кодона в разных рамках считывания, так что инициация трансляции, начатая с каждого из них, приводит к синтезу разных белков ( белок N и белок NSs у вирусов рода Orthobunyavirus ). Кроме того, у вирусов семейства Bunyaviridae трансляция некоторых мРНК приводит к образованию полипротеина с его последующим разрезанием.
Если у негативно-нитевых вирусов с несегментированным геномом, входящих в порядок Mononegavirales , каждая мРНК комплементарна участку одного гена, а позитивно-нитевая РНК, образующаяся при репликации, комплементарна всему геному, то у вирусов с сегментированным геномом каждый РНК-сегмент не только транскрибируется, но и реплицируется совершенно автономно, без связи с другими сегментами. Поэтому у вирусов с сегментированным геномом не только мРНК, но и те позитивно-нитевые РНК, которые участвуют в репликации, представляют собой комплементарные копии индивидуальных геномных сегментов. Тем не менее между молекулами мРНК и позитивными РНК-транскриптами, участвующими в репликации, имеются различия. У вирусов семейства Orthomyxoviridae мРНК на 5'-конце имеет добавочный кэпированный 5'-концевой участок длиной 10-14 п.о.: вирусная транскриптаза "похищает" этот участок у клеточных пре-мРНК, отщепляя его и используя в качестве праймера при синтезе вирусных мРНК. Кроме того, транскриптаза при синтезе мРНК завершает синтез, не дойдя до конца геномной РНК-матрицы. В результате молекулы мРНК имеют "лишний" участок на 5'-конце, но не содержат участка, соответствующего нескольким последним нуклеотидам геномного РНК-сегмента. Напротив, позитивные РНК-транскрипты, участвующие в репликации, являются полными и точными комплементарными копиями геномных РНК-сегментов.
Читайте также: