
Вегетативная форма вируса это

Вирусы — это микроскопические патогены, заражающие клетки живых организмов для самовоспроизводства. Они состоят из одного вида нуклеиновой кислоты (или ДНК или РНК, но не обе вместе), которая защищена оболочкой, содержащей белки, липиды, углеводы или их комбинацию. Размер типичного вируса варьируется от 15 до 350 нм, поэтому его можно увидеть только с помощью электронного микроскопа.
В 1892 году русский ученый Д.И. Ивановский впервые доказал существование ранее неизвестного типа возбудителя болезней, это был вирус мозаичной болезни табака. А в 1898 году Фридрих Лоффлер и Пол Фрош нашли доказательства того, что причиной ящура у домашнего скота была инфекционная частица, которая меньше, чем любая бактерия. Это были первые шаги к изучению природы вирусов, генетических образований, которые лежат где-то в серой зоне между живыми и неживыми состояниями материи. На текущий момент описано около 6 тыс. вирусов, но их существует несколько миллионов.
Строение вирусов
Вне клеток-хозяев вирусы существуют в виде белковой оболочки (капсида), иногда заключенного в белково-липидную мембрану. Капсид обволакивает собой либо ДНК, либо РНК, которая кодирует элементы вируса. Находясь в такой форме вне клетки, вирус метаболически инертен и называется вирионом.
Простая структура, отсутствие органелл и собственного метаболизма позволяет некоторым вирусам кристаллизоваться, т.е. они могут вести себя подобно химическим веществам. С появлением электронных микроскопов было установлено, что их кристаллы состоят из тесно прижатых друг к другу нескольких сотен миллиардов частиц. В одном кристалле вируса полиомиелита столько частиц, что ими можно заразить не по одному разу всех жителей Земли.
Формы вирусов
Вирусы встречаются в трех основных формах. Они бывают:
- Сферическими (кубическими или полигидральными). Вирусы герпеса, типулы, полиомы и т.д.
- Спиральными (цилиндрическими или стержнеобразными). Вирусы табачной мозаики, гриппа, эпидемического паротита и др.
- Сложными. Например, бактериофаги.
Проникновение вирусов в клетку-хозяина
Капсид в основном защищает нуклеиновую кислоту от действия клеточного нуклеазного фермента. Но некоторые белки капсида способствуют связыванию вируса с поверхностью клеток-хозяев, и работают, как ключики, вставляемые в нужные замочки. Другие поверхностные белки действуют как ферменты, они растворяют поверхностный слой клетки-хозяина и таким образом помогают проникновению нуклеиновой кислоты вируса в клетку-хозяина.
Жизненный цикл вирусов сильно отличается у разных видов, но существует шесть основных этапов жизненного цикла вирусов:
Присоединение к клетке-хозяину представляет собой специфическое связывание между вирусными капсидными белками и рецепторами на клеточной поверхности. Эта специфика определяет хозяина вируса.
Проникновение следует за прикреплением: вирионы проникают в клетку-хозяина через рецептор-опосредованный эндоцитоз или слияние мембран. Это часто называют вирусной записью.
Проникновение вирусов в клетку достигается за счет:
Размножение вирусов
После того, как вирусный геном освобождается от капсида, начинается его транскрипция или трансляция. Именно эта стадия вирусной репликации сильно различается между ДНК- и РНК-вирусами и вирусами с противоположной полярностью нуклеиновой кислоты. Этот процесс завершается синтезом новых вирусных белков и генома (точных копий внедрённых).
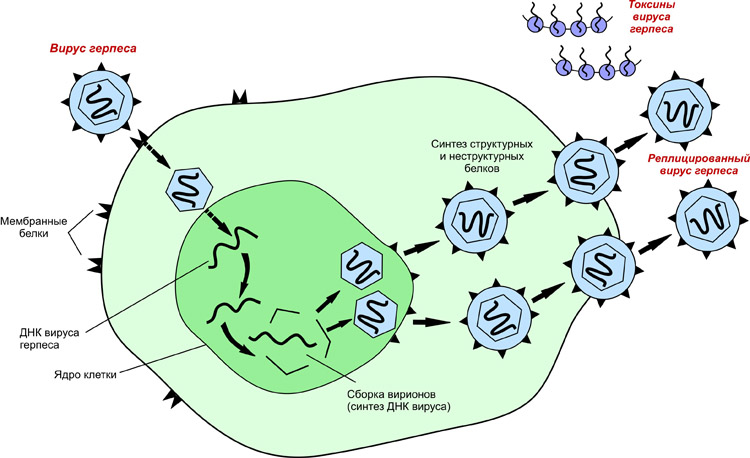
Механизм репликации зависит от вирусного генома.
- ДНК-вирусы обычно используют белки и ферменты клетки-хозяина для получения дополнительной ДНК, она транскрибируется в РНК-мессенджер (мРНК), которая затем используется для управления синтезом белка.
- РНК-вирусы обычно используют ядро РНК в качестве матрицы для синтеза вирусной геномной РНК и мРНК. Вирусная мРНК направляет клетку-хозяина на синтез вирусных ферментов и капсидных белков и сборку новых вирионов. Конечно, есть исключения из этого шаблона. Если клетка-хозяин не обеспечивает ферменты, необходимые для репликации вируса, вирусные гены предоставляют информацию для прямого синтеза отсутствующих белков.
Чтобы преобразовать РНК в ДНК, вирусы должны содержать гены, которые кодируют вирус-специфический фермент обратной транскриптазы. Она транскрибирует матрицу РНК в ДНК. Обратная транскрипция никогда не происходит в неинфицированных клетках. Необходимый фермент, обратная транскриптаза, происходит только от экспрессии вирусных генов в инфицированных клетках.
Вироиды
Вироиды заражают только растения. Одни вызывают экономически важные заболевания сельскохозяйственных культур, в то время как другие являются доброкачественными. Двумя примерами экономически важных вироидов являются кокосный cadang-cadang (он вызывает массовую гибель кокосовых пальм) и вироид рубцовой кожицы яблок, который безнадежно портит товарный вид яблок.
30 известных вироидов были классифицированы в две семьи.
- Члены семейства Pospiviroidae, названные по имени вироида клубневого веретена картофеля, имеют палочковидную вторичную структуру с небольшими одноцепочечными областями, имеет центральную консервативную область, и реплицируются в ядре клетки.
- Avsunviroidae, названный в честь вироида авокадо, имеет как палочковидную, так и разветвленную области, но не имеет центральной консервативной области и реплицируется в хлоропластах растительной клетки.
В отличие от вирусов, которые являются паразитами механизма трансляции хозяина, вироиды являются паразитами клеточных транскрипционных белков.
Бактериофаги

Существуют тысячи разновидностей фагов, каждый из которых может заразить только один тип или несколько близких типов бактерий или архей. Фаги классифицируются по ряду семейств вирусов; например:
Как и все вирусы, фаги являются простыми организмами, которые состоят из ядра генетического материала (нуклеиновой кислоты), окруженного капсидом белка. Нуклеиновая кислота может представлять собой либо ДНК, либо РНК, и может быть двухцепочечной или одноцепочечной.
Существует три основных структурных формы фага:
- Икосаэдрическая (20-сторонняя) головка с хвостом
- Икосаэдрическая головка без хвоста
- Нитевидная форма
Во время заражения фаг прикрепляется к бактерии и вставляет в нее свой генетический материал. После этого фаг обычно следует одному из двух жизненных циклов: литическому (вирулентному) или лизогенному (умеренному).
Литические, или вирулентные, фаги захватывают механизм клетки, чтобы скопировать компоненты фага. Затем они разрушают или лизируют клетку, высвобождая новые частицы фага.
Лизогенные, или умеренные, фаги включают свою нуклеиновую кислоту в хромосому клетки-хозяина и реплицируются с ней как единое целое, не разрушая клетку. При определенных условиях лизогенные фаги могут индуцироваться в соответствии с литическим циклом.
Существуют и другие жизненные циклы, в т.ч. псевдолизогенез и хроническая инфекция. При псевдолизогении бактериофаг проникает в клетку, но не использует механизм репликации клеток и не интегрируется в геном хозяина, просто как бы прячется внутри бактерии, не нанося ей никакого вреда. Псевдолизогенез возникает, когда клетка-хозяин сталкивается с неблагоприятными условиями роста и, по-видимому, играет важную роль в выживании фага, обеспечивая сохранение генома фага до тех пор, пока условия роста хозяина снова не станут благоприятными.
При хронической инфекции новые фаговые частицы образуются непрерывно и длительно, но без явного уничтожения клеток.
Вскоре после открытия фаги начали использовать для лечения бактериальных заболеваний человека, таких как бубонная чума и холера. Но фаговая терапия тогда не была успешной, и после открытия антибиотиков в 1940-х годах она была практически заброшена. Однако с появлением устойчивых к антибиотикам бактерий терапевтическому потенциалу фагов уделяется все больше внимания.
Наше время с антибиотиками заканчивается. В 2016 году женщина в штате Невада умерла от бактериальной инфекции, вызванной Klebsiella pneumoniae, которая была устойчивой ко всем известным антибиотикам. Бактерии, устойчивые к колистину, антибиотику последней инстанции, были обнаружены на свинофермах в Китае. В настоящее время бактерии приспосабливаются к антибиотикам быстрее, чем когда-либо.
Покажите ножницы которыми вирусы разрезают молекулу РНК что бы встроиться для мутации.Может что нибудь придумаете другое.К примеру деление цепочка аминокислот получив энергию из вне как одноименные заряды распадается на две. К каждой соединятся только те какие были ранее (другие проскочат мимо),казалось бы копии,но внутренняя энергия разная(уменьшается увеличивается) поэтому распад и создание. Вся химия углерода на этом построена 1000 орган соединений создает у других хим элементов этого свойства нет. Иммунная система делает накладку(интерференция)с помощью энергии интерферонов пытаясь разрушить цепочку РНК вируса.Надо помочь организму но не вакциной(вирус быстро мутирует)
Вирусы – неклеточная форма жизни, обладает собственным геномом, способностью к самовоиспроизведению (репродукции) в клетках живых организмов или клеточных культурах, адаптационными свойствами и изменчивостью.
Выделены в отдельное царство – Vira.
- нет клеточной организации: не имеют цитоплазмы и ядра, митохондрий, рибосом и других органелл
- содержат только одну из двух нуклеиновых кислот – ДНК или РНК, выполняющих функции генома.
- не имеют собственных белоксинтезирующих и генерирующих энергию систем и являются абсолютными внутриклеточными паразитами на генетическом уровне, полностью зависят от клетки-хозяина
- размножаются не обычным бинарным делением, а репродуцируются в чувствительной клетке, согласно генетической программе в нуклеиновой кислоте вируса, при этом используют биосинтетические системы и ресурсы
Различают две формы существования вирусов – внеклеточную и внутриклточную.
Внеклеточный вирус = вирион. Это покоящаяся (зрелая) форма вируса. Не проявляет жизедеятельности. Функции: сохранение вируса во внешней среде и перенос его из организма в другой организм или из клетки в другую клетку.
Внутриклеточный вирус - вегетативный вирус - репродуцируется в инфицированной клетке, вызывая репродуктивную инфекцию, заканчивающуюся образованием дочернего поколения вирионов и, как правило, гибелью клетки. Процесс репродукции может быть незавершенным, без образования вирионов – возникает абортивная инфекция.
Некоторые вирусы способны встраивать свой генетический материал в хромосомы клетки-хозяина в виде провируса, которые реплицируется вместе с этой хромосомой в процессе деления и переходит в дочерние клетки. Это – интегративная инфекция, она модет существовать длительное время или переходить обратно в продуктивную.
Морфология и строение вирусов (вирионов). Размеры вирусов находятся в диапазоне 20-350 нм.
Могут иметь палочковидную, многогранную, пулевидную, сферическую, нитевидную, булавовидную формы.
Различают: простые (безоболочечные) и сложные (оболочечные) вирусы. У них в центре – молекула нуклеиновой кислоты (ДНК/РНК), окруженная белковой оболочкой – капсидом. Вся структура носит название – нуклокапсид.
Простые вирусы – нуклеиновая кислота, ассоциированная с внутренними белками и капсидом (т.е. представляют собой нуклеокапсид).
Защитная белковая оболочка – капсид – состоит из множества однородных белковых субъединиц. Т.к. на такое строение капсида расходуется мало генетической информации, оно важно для вирусов, обладающих небольшим геномом. Капсиды построены по спиральному или кубическому типу симметрии, в зависимости от расположения белковых субъединиц.
Типы симметрии.
1.Спиральный. Нуклеиновая кислота расположена винтообразно. Вокруг нее по спирали располагаются структурные белковые субъединицы овоидной формы (протомеры), надежно закрывающие нуклеиновую кислоту вируса. Расходуется много белка, но структура – прочная.
Форма нуклеокапсида полочковидная или нитевидная.
Простые вирусы со спиральным типом симметрии - в основном вирусы растений (вирус табачной мозаики – палочковидный, бактериофаги – нитевидные). Среди поражающих человека простых вирусов - таких не встречается.
Сложные вирусы со спиральным типом симметрии (орто-, пара-, миксовирусы) имеют нуклеокапсид, представляющий собой длинный тяж, упакованный в виде клубка или плотной спирали, который покрыт суперкапсидом, благодаря чему вирион имеет сферическую форму.
2. Кубический тип симметрии. Большинство вирусов посмотрены по этому типу, форма – икосаэдра (20-гранник). Капсид построен из морфологических единиц – капсомеров, имеющих шаровидную, иногда призматическую форму. Каждый капсомер состоит из 5 (пентомер) или 6 (секстомер) структурных белковых единиц. В основе кубической симметрии – комбинации равносторонних треугольников, образуемых капсомерами, что ведет к формированию замкнутой сферической поверхности с полостью внутри.
Белка затрачивается меньше, чем при спиральном типе симметрии. Менее прочен и ненадежно закрывает нуклеиновую кислоту.
Капсиды разных вирусов состоят из различного, но строго определенного для данной таксономической группы, для каждого типа, вида вирусов, количества капсомеров (у вируса полиомиелита – 32 капсомера, у вируса гепатита В – 180 капсомеров).
Последнее изменение этой страницы: 2017-01-19; Нарушение авторского права страницы
Многие вирусы, патогенные для человека и животных, хорошо культивируются в развивающемся курином эмбрионе. К ним относятся вирусы вакцины, бешенства, свинки, кори, герпеса и ряд других. Для инокуляции вируса чаще всего используется оболочка, желточный мешок и некоторые другие части эмбриона.
Инокуляции подвергаются оплодотворенные яйца, прошедшие инкубирование при 38° С в течение 3—13 дней. Подходящий возраст эмбриона определяется в зависимости от особенностей данного вируса, а также от того, какой орган избирается для инокуляции. Для нанесения вирусного материала на оболочку она обнажается путем удаления небольшого участка скорлупы и разрыва скорлуповой оболочки. Нужное для операции место определяется при просвечивании яйца. Вскрытый участок скорлупы после заражения закрывается покровным стеклом, края которого заливаются парафином.
После инокуляции яйца инкубируются при 36° С, и в них размножается вирус.
Для накопления вирусов широко применяются также культуры нормальных и опухолевых тканей. Наиболее распространен способ однослойной культуры тканей на стекле. Питательная среда содержит плазму или сыворотку крови, к которым в некоторых случаях прибавляют экстракт куриного эмбриона. Добавляется также сбалансированный солевой раствор.
Размножение фитопатогенных вирусов. Проникнув в результате инокуляции в клетки растения-хозяина, фитопатогенные вирусы размножаются, претерпевая при этом цикл развития, включающий качественно различные стадии, или фазы. Весь цикл развития вирусов пока не исследован. Наиболее изучен в этом отношении вирус табачной мозаики.
В развитии этого вируса отчетливо выделяются две фазы, хотя в действительности их, возможно, больше: вегетативная фаза и фаза покоя.
Вегетативная фаза, во время которой вирус размножается, наступает через 1—2 ч после заражения восприимчивого хозяина в условиях оптимальной температуры (Сухов и Капица, 1956). Размножаясь вначале в отдельных клетках, вегетативный вирус по плазмодесмам проникает в смежные клетки, где также размножается. Попав в ту или иную клетку, вегетативный вирус размножается в ней не беспредельно, а ограниченное время, длящееся приблизительно 9—10 ч. Только после этого в данной клетке вирус переходит из фазы размножения в фазу покоя.
Переход в фазу покоя обусловлен исчерпыванием субстратов, необходимых для размножения вируса, или же накоплением продуктов обмена, тормозящих его дальнейшее размножение.
Вегетативный вирус по ряду свойств отличается от покоящегося. Характерным отличием является его неспособность заражать новые растения при использовании обычных методов инокуляции. Вегетативный вирус прак-тически лишен контагиозности, что объясняется быстрой его инактивацией при разрушении тканей в момент приготовления инокулюма.
Установлено, что при выдерживании листьев с накопленным в них вегетативным вирусом в течение 2—3 ч при 28° С в них появляется значительное количество инфекционного вируса.
Течение инфекционного процесса в клетке сопровождается определенными изменениями в ее органоидах. По данным Виттштейна и Цеха (1962), первая фаза репродукции ВТМ характеризуется усилением синтеза нуклеиновых кислот в ядре и увеличением размеров последнего. Эти изменения становятся заметными уже через 30 мин после заражения и прогрессируют в течение следующих 2—3 ч.
Ядро при этом пронизывается системой цитоплазматических каналов, окаймленных ядерной мембраной. В каналах находятся митохондрии, мембраны Гольджи или эндоплазматического ретикулума. Через 3—7 ч после заражения в цитоплазме, окаймляющей ядро, появляется материал, абсорбирующий ультрафиолетовые лучи в области волн, поглощаемых нуклеиновыми кислотами. К концу этого периода содержание нуклеиновых кислот в ядре падает и абсорбирующий материал из области, окружающей ядро, рассеивается в цитоплазме. Масса цитоплазмы при этом увеличивается. В ней интенсивно накапливаются митохондрии, элементы Гольджи, эндоплазматический ретикулум и другие цитоплазматические органоиды.
Начиная с 9-го ч после заражения, в цитоплазме накапливается вирусный белок, участвующий в образовании спирализованных вирионов ВТМ, и к 25-му ч формируются новые кристаллические включения вируса.
Размножение бактериофагов. Процесс заражения бактерио-фагом бактериальной клетки начинается с адсорбции вирионов бактериофага на ее оболочке.
Бактериофаги на бактериальных клетках адсорбируются при всех градациях температуры от 0 до 37° С. Однако бактериофаг при низкой температуре исторгает генеративный материал не в клетку, а в окружающую среду и заражения бактерии не происходит.
Инъекция ДНК бактериофага в бактериальную клетку происходит замедленно при 37° С и сильно ускоряется при 44° С, что связано с уменьшением вязкости нуклеиновой системы.
При гидролизе участка бактериальной оболочки, вызываемом ферментом прикрепившегося бактериофага, появляются вещества со свободными аминогруппами. Предполагается, что эти вещества вызывают исторжение ДНК (Гарен и Козлов, 1959).
Бактериофаги, имеющие спермиеобразную форму, адсорбируются на бактериальной оболочке концом хвостовидного придатка. В результате действия ферментной системы, заключенной в хвостовидном отростке, целостность оболочки нарушается, образуется отверстие, через которое в цитоплазму клетки исторгается внутреннее содержимое корпускулы, состоящее у бактериофага из ДНК, белка и аргининсодержащего вещества. Основную массу генеративной системы составляет нуклеиновая кислота.
После внедрения генеративного материала в цитоплазму бактерии наступает латентный период инфекции, в течение которого бактериофаг находится в состоянии размножения. Разные формы бактериофага имеют различную продолжительность латентного периода.
В вегетативной фазе развития бактериофаг не обладает приспособлениями, необходимыми для внедрения в новые бактериальные клетки, и поэтому неинфекционен. Если механически разрушить бактериальные клетки, несущие вегетативный бактериофаг, и внести этот материал в культуру восприимчивых бактерий, лизиса не наступает. неспособен заражать новые бактериальные клетки.
После проникновения генеративной системы бактериофага в клетку начавшийся инфекционный процесс проявляется прежде всего в разрушении нуклеоидов бактерии. В это же время начинается синтез ДНК бактериофага. В различных участках клетки образуются вакуоли, заполняющиеся тонким фибриллярным материалом, состоящим из рыхло расположенных нитей вирусной ДНК. Эти области имеют неправильные очертания, но они ограничены и фибриллы не смешиваются с рибосомами цитоплазмы.
В местах скопления фибрилл образуются плотные тельца, по величине близкие к головке бактериофага, но лишенные белковой оболочки. Их образование связано с конденсацией индивидуальных цепей ДНК. В это время в участках скопления указанных частиц сосредоточены значительные массы белковых субъединиц. В краткий период времени, длящийся около 1 мин, происходит формирование белковой оболочки, окружающей каждое нуклеиновое тельце. В следующие 5 мин завершается созревание частиц бактериофага, и они приобретают свой характерный вид.
Размножение зоопатогенных вирусов. Довольно хорошо изучен процесс репродукции малых, содержащих РНК энтеровирусов, к которым относятся вирусы полиомиелита, Коксаки и др.
Первый этап инфекции вируса полиомиелита связан с адсорбцией вирионов на поверхности восприимчивых клеток. Адсорбция специфична и происходит только на клетках тканей приматов и человека. На клетках других млекопитающих вирионы этого вируса не адсорбируются. Вызывающий адсорбцию рецептор в оболочках клеток приматов представлен специфическими липопротеидами. Специфичность адсорбции обусловлена свойствами белковой оболочки вириона. Инфекционная РНК вируса, выделенная из вирионов, лишена этой специфичности и может вызвать заражение клеток в культуре тканей млекопитающих, относящихся к разным отрядам. Однако после прохождения одного цикла развития вновь образовавшиеся вирионы неспособны возобновить инфекционный процесс в культурах тканей этих животных.
После адсорбции вирионов на плазматических мембранах клеток в культуре тканей происходит нарушение целостности капсиды с освобождением вирусной РНК. Этот материал проникает внутрь клетки посредством пиноцитоза, т. е. вовлечения частиц внутрь цитоплазмы в результате образования глубоких впячиваний мембраны, преобразующихся затем в мелкие вакуоли.
Латентный период инфекции вируса полиомиелита длится 2,5—3 ч. В это время в клетках протекают следующие процессы. РНК вируса связывается с рибосомами и при посредстве их аппарата индуцирует синтез специфической вирусной репликазы. Последняя, взаимодействуя с вирусной РНК, катализирует синтез новых цепей РНК. Несколько позже, используя рибосомы, вирусная РНК индуцирует синтез белков, необходимых для образования оболочки вирионов.
Процесс объединения цепей РНК и белковых субъединиц в структуру вирионов происходит в конце латентного периода и определяется, вероятно, физической комплементарностью этих компонентов.
Таким образом, рибосомы являются теми органоидами клетки, где вирусная РНК осуществляет паразитическую функцию и организует синтез вирусных белков.
Латентные вирусные инфекции. Латентное носительство вирусов, когда их присутствие в организме хозяина не проявляется никакими внешними симптомами, широко распространено в природе.
Среди вирусов, патогенных для животных, также известны латентные формы. Например, вирус подчелюстных желез морской свинки часто присутствует у внешне здоровых животных. В клетках желез он образует характерные внутриядерные включения. При подкожном заражении свинок материалом из инфицированных желез не возникает никаких внешних симптомов, но в ядрах подчелюстных желез вскоре появляются включения. Только при интрацеребральном введении такого материала молодым животным возникает тяжелое поражение, и через 5—7 дней они погибают.
Явление латентной инфекции характеризуется наиболее уравновешенными отношениями между хозяином и паразитом и обеспечивает для последнего наилучшие условия для выживания и распространения в природе. Еще более совершенную форму взаимоотношений представляет симбиоз, когда организм хозяина получает какие-либо преимущества от поселившегося в нем симбионта. В случае фито- и зоопатогенных вирусов положительное значение для хозяина может иметь бессимптомное носительство непатогенных штаммов, обеспечивающее инфекционный иммунитет против высоковирулентных штаммов. Такое же значение имеет лизогенность бактерий, сопряженная с иммунитетом против лизирующего бактериофага.
Открытие вирусов
В 1892 году Д.И. Ивановский (см. Рис. 1), изучая мозаичную болезнь табака (см. Рис. 2), установил, что причиной заболевания является некое инфекционное начало, содержащееся в листьях больных растений, которое проходит через фильтр, задерживающий обыкновенные бактерии. Если профильтрованный сок внести в листья здоровых растений, то они также заболевают мозаичной болезнью.

Рис. 1. Д.И. Ивановский

Рис. 2. Мозаичная болезнь табака
В 1898 году независимо от Ивановского аналогичные результаты получил голландский микробиолог М. Бейеринк. Однако он предположил, что мозаичную болезнь табака вызывают не мельчайшие бактерии, а некое жидкое заразное начало, которое он назвал фильтрующим вирусом.
Размеры вирусов определяются нанометрами (20-200 нм), поэтому их изучение началось после открытия электронного микроскопа. В настоящее время описаны вирусы практически всех групп живых организмов.
Строение вирусов
Вирусы – неклеточные формы жизни. Они состоят (см. Рис. 3) из фрагмента генетического материала (РНК или ДНК), составляющего сердцевину вируса, и защитной оболочки, которая называется капсид. У некоторых вирусов (герпес, грипп) есть дополнительная липопротеидная оболочка – суперкапсид, которая возникает из плазматической мембраны клетки-хозяина.

Рис. 3. Строение вируса
Вирусы не способны к самостоятельной жизнедеятельности. Они могут проявлять свойства живого, только попав в клетку-хозяина. Они используют потенциал и энергию этой клетки для создания своих новых вирусных частиц, следовательно, вирусы являются внутриклеточными паразитами.
Размножение вирусов
Обычно вирус связывается с поверхностью клетки-хозяина и проникает внутрь. Каждый вирус ищет своего хозяина, то есть клетки строго определенного вида. Например, вирус – возбудитель гепатита (желтуха) проникает и размножается только в клетках печени, а вирус эпидемического паротита (свинка) – только в клетках околоушных слюнных желез человека.
Проникнув внутрь клетки-хозяина, вирусная ДНК или РНК начинает взаимодействовать с ее генетическим аппаратом таким образом, что клетка начинает синтезировать белки, свойственные вирусу (см. Рис. 4).

Рис. 4. Схема репродукции вируса
При заражении ретровирусом (например, вирус иммунодефицита человека (ВИЧ)), у которого в качестве генетического материала используется молекула РНК, наблюдается другая картина. При попадании ретровируса в клетку-хозяина происходит обратная транскрипция. То есть на основе вирусной РНК синтезируется вирусная ДНК, которая встраивается в ДНК человека. Такой тип взаимодействия вируса с клеткой называется интегративным, а встроенная в состав хромосомы клетки ДНК вируса называется провирусом. Далее провирус реплицируется (удваивается) в составе хромосомы и переходит в геном дочерних клеток. Однако под влиянием некоторых физических и химических факторов провирус может выщепляться из хромосомы клетки и переходить к продуктивному типу взаимодействия, то есть синтезировать новые вирусные частицы.
При заражении ВИЧ человек чувствует себя здоровым, пока вирусный генетический материал встроен в хромосому человека. Однако при выщеплении этого вирусного генетического материала из клетки она начинает образовывать новые вирусные частицы, вследствие чего развивается смертельное заболевание – синдром приобретенного иммунодефицита (СПИД).
Вирусы являются возбудителями большого количества заболеваний человека: корь, грипп, оспа, краснуха, энцефалит, свинка, гепатиты, СПИД. Известен также целый ряд заболеваний растений, вызываемых вирусами, например мозаичная болезнь табака, томатов, огурцов или скручивание листьев картофеля. Всего описано около 500 видов вирусов, поражающих клетки позвоночных животных, и около 300 вирусов растений. Некоторые вирусы участвуют в злокачественном перерождении клеток и тем самым провоцируют онкологические заболевания.
ДНК- и РНК-содержащие вирусы
В зависимости от содержащегося генетического материала вирусы подразделяются на ДНК-содержащие и РНК-содержащие.
Одноцепочные РНК-содержащие вирусы подразделяются на:
1. Плюс-нитевые (положительные). Плюс-нить РНК этих вирусов выполняет наследственную (геномную) функцию и функцию информационной РНК (иРНК).
2. Минус-нитевые (отрицательные). Минус-нить РНК этих вирусов выполняет только наследственную функцию.
К РНК-содержащим вирусам относятся более 
вирусов, вызывающих респираторные заболевания, а также вирус гриппа, кори, краснухи, свинки, ВИЧ. Также существует специфическая группа вирусов – арбовирусы, которые переносятся членистоногими.
Двухцепочные ДНК-содержащие вирусы вызывают такие заболевания, как папиллома человека или герпес, гепатит В (гепатит А и гепатит С вызывается РНК-содержащими вирусами).
ДНК-содержащие вирусы поражают также растения. Они вызывают, например, золотую мозаику бобов или полосатость у кукурузы.
Вирус гепатита С
По своему строению вирус гепатита С – это РНК-содержащий вирус, имеющий сферическую форму, сложно устроенный (см. Рис. 5).
В качестве генетического материала такой вирус содержит линейную однонитчатую молекулу РНК.

Рис. 5. Гепатит С
Вопреки бытующим предрассудкам, подцепить вирус гепатита C невозможно через социальные контакты (поцелуи, объятия), через продукты или воду, через грудное молоко. Вы ничем не рискнете, если разделите с носителем вируса трапезу или напитки. Заразиться гепатитом C можно при контакте с кровью инфицированного человека либо половым путем.
В настоящее время для лечения гепатита С используют два препарата: Интерферон альфа и Рибавирин.
Бактериофаги

Рис. 6. Бактериофаг (Источник)
Особую группу вирусов составляют бактериофаги (или просто фаги), которые заражают бактериальные клетки (см. Рис. 6). Фаг укрепляется на поверхности бактерии при помощи специальных ножек и вводит в ее цитоплазму полый стержень, через который проталкивает внутрь клетки свою ДНК или РНК. Таким образом, генетический материал фага попадает внутрь бактериальной клетки, а капсид остается снаружи. В цитоплазме начинается репликация генетического материала фага, синтез его белков, построение капсида и сборка новых фагов. Уже через 10 мин после заражения в бактерии формируются новые фаги, а через полчаса бактериальная клетка разрушается, и из нее выходят около 200 заново сформированных вирусов – фагов, способных заражать другие бактериальные клетки (см. Рис. 7). Некоторые фаги используются человеком для борьбы с болезнетворными бактериями, вызывающими холеру, дизентерию, брюшной тиф.

Рис. 7. Схема размножения бактериофага (Источник)
Список литературы
- Каменский А.А., Криксунов Е.А., Пасечник В.В. Общая биология 10-11 класс Дрофа, 2005.
- Биология. 10 класс. Общая биология. Базовый уровень / П.В. Ижевский, О.А. Корнилова, Т.Е. Лощилина и др. – 2-е изд., переработанное. – Вентана-Граф, 2010. – 224 стр.
- Беляев Д.К. Биология 10-11 класс. Общая биология. Базовый уровень. – 11-е изд., стереотип. – М.: Просвещение, 2012. – 304 с.
- Агафонова И.Б., Захарова Е.Т., Сивоглазов В.И. Биология 10-11 класс. Общая биология. Базовый уровень. – 6-е изд., доп. – Дрофа, 2010. – 384 с.
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
Домашнее задание
Если вы нашли ошибку или неработающую ссылку, пожалуйста, сообщите нам – сделайте свой вклад в развитие проекта. 

Читайте также: