
Метаболизм костной и хрящевой ткани что это
Препараты подгрупп исключены. Включить
Препараты
| Действующее вещество | Торговые названия |
| Информация отсутствует | Адгелон ® Алфлутоп АМБЕНЕ ® Био Гиалгель гель для тела оздоравливающий с Гиалуронатом натрия и Тизолем ГИАЛРИПАЙЕР-02 Хондрорепарант ® Материал гелевый на основе 0,8% модифицированной гиалуроновой кислоты водосодержащий стерильный ГИАЛРИПАЙЕР-02 Хондрорепарант ® Материал гелевый на основе 1,5% модифицированной гиалуроновой кислоты водосодержащий стерильный ГИАЛРИПАЙЕР-10 Хондрорепарант ® Материал гелевый на основе 0,8% модифицированной гиалуроновой кислоты водосодержащий стерильный ГИАЛРИПАЙЕР-10 Хондрорепарант ® Материал гелевый на основе 1,5% модифицированной гиалуроновой кислоты водосодержащий стерильный ДОНА ® Остеогенон Пиаскледин 300 Стекловидное тело Супер Кальций с магнием, цинком, витаминами D и Эстер-C Тридин Хондрактив Хондрамин ® |
| Алендроновая кислота* (Acidum alendronicum) | Алендрокерн Алендронат Алендронат Канон Алендронат КЕРН ФАРМА Алендронат натрия Алендронат натрия тригидрат Алендронат Плива Биносто Осталон ® Остерепар ® Стронгос Теванат ® Фороза ® Фосамакс ® |
| Альфакальцидол* (Alphacalcidolum) | Альфа Д3-Тева ® Альфадол Ван-Альфа ® Оксидевит Оксидевита капсулы Этальфа ® |
| Гиалуроновая кислота* (Acidum hyaluronicum) | Гиалган Фидия |
| Гликозаминогликан-пептидный комплекс (Glicosamineglycane-peptide complex) | Биартрин Румалон ® |
| Глюкозамин + Хондроитина сульфат (Glucosaminum+ Chondroitini sulfas) | Тазан ® Хондроглюксид |
| Глюкозамин* (Glucosaminum) | D-Глюкозамина сульфат натрия хлорид Аминоартрин ® Артракам ® Глюкозамин Глюкозамин сульфат ГЛЮКОЗАМИН ЭВАЛАР Глюкозамина гидрохлорид Глюкозамина сульфат 750 Глюкозамина сульфат натрия хлорид ДОНА ® Сустагард ® Артро Сустилак Фармаскин ТГК Хондроксид ® Максимум Эльбона ® Юниум ® |
| Деносумаб* (Denosumabum) | Пролиа ® Эксджива ™ |
| Дигидротахистерол* (Dihydrotachysterolum) | А.Т.10 ® Дигидротахистерол Тахистин |
| Золедроновая кислота* (Acidum zoledronicum) | Акласта ® Блазтера ® Верокласт Золедронат-Тева Золедроновая кислота Золедроновая кислота моногидрат Золедрэкс Золендроник-Рус 4 Золера Золерикс Зомета ® Метастазол Остеостатикс Резокластин Резокластин ФС Резорба Резоскан, 99mTc Таргезол |
| Ибандроновая кислота* (Acidum ibandronicum) | Бонвива ® Бондронат ® Виванат Ромфарм Резовива |
| Иприфлавон* (Ipriflavonum) | Остеохин |
| Кальцитонин* (Calcitoninum) | Алостин Вепрена ® Кальцитонин Кальцитонин-депо Миакальцик ® Остеовер |
| Кальцитриол* (Calcitriolum) | Остеотриол Рокальтрол ® |
| Клодроновая кислота* (Acidum clodronicum) | Бонефос ® Клобир |
| Колекальциферол* (Colecalciferolum) | Видехол Видехола раствор в масле Компливит ® Аква Д3 |
| Натрия гиалуронат (Natrii hyaluronas) | Stylage Hydro max Имплантат вязко-эластичный для инъекционной контурной пластики Stylage Hydro Имплантат вязко-эластичный для инъекционной контурной пластики Stylage L Имплантат вязко-эластичный для инъекционной контурной пластики Stylage M Lidocaine Имплантат вязко-эластичный для инъекционной контурной пластики Stylage M Имплантат вязко-эластичный для инъекционной контурной пластики Stylage S Имплантат вязко-эластичный для инъекционной контурной пластики Stylage Special Lips Lidocaine Имплантат вязко-эластичный для инъекционной контурной пластики Stylage XL Имплантат вязко-эластичный для инъекционной контурной пластики АДАНТ ® Айстил Гиалуронат натрия Гилан Комфорт Раствор увлажняющий офтальмологический Гилан Ультра комфорт Раствор увлажняющий офтальмологический Интрагель 0,8% Интрагель 1,6% Натрия гиалуронат РИПАРТ ® Средство для замещения синовиальной жидкости РИПАРТ ® Лонг Средство для замещения синовиальной жидкости Синоарт ® Синовиаль Хеалон ® |
| Натрия фторид (Natrii fluoridum) | Натриум флуоратум Натрия фторид Оссин |
| Памидроновая кислота* (Acidum Pamidronicum) | Аредиа Памидронат медак Помегара |
| Парикальцитол* (Paricalcitolum*) | Земплар ® |
| Ризедроновая кислота* (Acidum risedronicum) | Актонель Ризартева Ризендрос |
| Стронция ранелат (Strontii ranelas) | Бивалос ® Остеолат Строметта |
| Терипаратид* (Teriparatidum) | Форстео ® |
| Тилудроновая кислота* (Acidum tiludronicum) | Скелид |
| Хондроитина сульфат (Chondroitini sulfas) | Артогистан АРТРАВИР ® -ИНКАМФАРМ ® АРТРАВИР ® -ТРИВИУМ ® Артрадол ® Артрафик ® АРТРА ® Хондроитин Артрин ® Биоактивный экстракт из мелкой морской рыбы с хондроитином ДРАСТОП ИНЪЕКТРАН ® Мукосат ® Натрия хондроитина сульфат Структум ХОНДРОГАРД ® Хондроитин Хондроитин Велфарм Хондроитин Ди Икс из Акульего Хряща Хондроитин сульфат Хондроитин сульфат натрия Хондроитин сульфат-Тривиум Хондроитин-АКОС Хондроитин-ВЕРТЕКС Хондроитина сульфат Хондроксид ® Хондроксид ® мазь Хондролон ® Хондромед-Лекфарм Хонсат Хонсурид ® |
| Этидроновая кислота* (Acidum etidronicum) | Ксидифон Монокалиевая соль 1-гидроксиэтилидендифосфоновой кислоты дигидрат Фосфотех, 99mTc |
Еще много интересного
Все права защищены.
Не разрешается коммерческое использование материалов.
Информация предназначена для медицинских специалистов.
ОСТЕОГЕНЕЗ (костеобразование) — процесс формирования костей у позвоночных. Осуществляется при помощи специальных клеток (остеобластов), выделяющих костеобразующие минеральные вещества, которые объединяются с сетью коллагеновых волокон, в результате чего образуется основное вещество твердых костей.
РЕЗОРБЦИЯ (от лат. resorbeo — поглощаю) — рассасывание, растворение (разрушение).
Термины:
Костная ткань – вид соединительной ткани, из которой образуются все кости в человеческом организме. Состоит из особых клеток (остеобластов, остеокластов, остеоцитов) и межклеточного вещества.
Межклеточное вещество (костный матрикс) составляет 50% сухого веса костной ткани и состоит из органической (25%), неорганической (50%) частей и воды (25%).
Остеобласты (от др.-греч.: ὀστέον — кость, βλάστη — росток, отпрыск, побег) – молодые клетки костной ткани диаметром 15-20 мкм, располагающиеся в верхних её слоях и вырабатывающие компоненты межклеточного вещества. Проще говоря – клетки-строители костной ткани.
Остеокласты (от др.-греч.: osteon — кость и clao — раздроблять, разбивать) – это крупные многоядерные (5 — 100 ядер) клетки костной ткани размерами до 190 мкм, которые разрушают кость и обызвествлённый хрящ (хрящ с отложениями кальция ).
Остеоциты (от др.-греч.: ὀστέον — кость, κύτος — вместилище, здесь — клетка) – основной тип клеток зрелой костной ткани длиной 20-55 мкм и шириной 5-15 мкм. Образуются из остеобластов в процессе развития костной ткани.
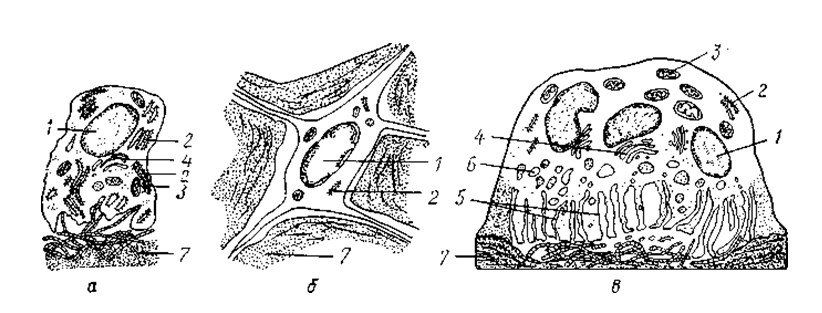
Рис. 1. Клетки костной ткани
а — остеобласт; б — остеоцит; в — остеокласт; 1 — ядро; 2 — гранулярная эндоплазматическая сеть; 3 — митохондрии; 4 — комплекс Гольджи; 5 — гофрированная каемка; 6 — лизосомы; 7 — межклеточное вещество кости (по Е. А. Шубниковой с изменениями)
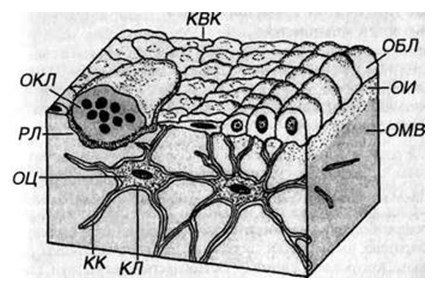
Рис. 2. Схема расположения клеток костной ткани
ОБЛ — остеобласты (активные), КВК — клетки, выстилающие кость (неактивные остеобласты), КЛ — костные лакуны с телами остеоцитов (ОЦ), КК — костные канальцы, содержащие отростки ОЦ, ОКЛ — остеокласт в резорбционной лакуне (РЛ), ОИ — остеоид, ОМВ — обызвествленное межклеточное вещество.
а) коллагеновые белки (90-95%);
б) неколлагеновые белки (их около 200);
в) протеогликаны (белки+небольшое количество углеводов) фиксируют ионы кальция в очагах остеогенеза, образуя комплексы с минералом;
г) гликозаминогликаны связывают большое количество воды, благодаря чему межклеточное вещество приобретает желеобразную консистенцию.
2.Синтез аденозинтрифосфорной кислоты(АТФ) — источника энергии для процесса синтеза органического матрикса и донора фосфата для минерализации.
3. Активное накопление в митохондриях остеобластов положительно заряженных ионов кальция (кальция в митохондриях в 500 раз больше, чем в цитоплазме остеобластов) и отрицательно заряженных ионов фосфатов.
Этапы:
1. Активное отпочковывание в межклеточный матрикс особых пузырьков (мембранных везикул), содержащих:
а) положительно заряженные ионы кальция (концентрация кальция в пузырьках в 25-50 раз выше, чем в остеобластах);
б) сложные жиры (глицерофосфолипиды) – нужны для связывания кальция и фосфора с органическим каркасом;
в) щелочную фосфатазу (ЩФ) – ей принадлежит ведущая роль в образовании центров кристаллизации гидроксиапатитов и ускорении процесса кристаллизации; также ЩФ ликвидирует вещество, тормозящее процесс минерализации (пирофосфат), расщепляя его.
г) другие фосфатазы (пиро-, АТФ-, 5′-АМФ-), необходимые для кальцификации органического матрикса.
2. Разрушение оболочек мембранных везикул и освобождение содержимого пузырьков.
3. Частичный распад протеогликанов, освобождение кальция и фосфатов и формирование поверхности белков, на которой будет происходить образование кристаллической решётки гидроксиапатитов.
4. Формирование ядер кристаллизации гидроксиапатитов. Этот процесс начинается ещё в мембранных везикулах.
5. Формирование кристаллической решётки из минералов.
Рассмотрены подходы к выбору средств для профилактики и лечения потерь костной ткани, восстановления ее структуры и качества. Применяемый препарат должен способствовать синтезу коллагена, формированию костного матрикса, его минерализации и, соответственно
Approaches to selecting the methods of prevention and treatment of bone tissue losses, restoration of its structure and quality, were considered. The used preparation must contribute to collagen synthesis, formation of bone matrix, its mineralization, and, respectively, increase of the bone density and strength.
Остеопороз (ОП) — прогрессирующее системное заболевание скелета, характеризующееся снижением костной массы и нарушением микроархитектоники (качества) костной ткани, что приводит к хрупкости костей и повышению риска переломов. ОП — самое распространенное заболевание костной ткани: остеопоротические переломы отмечается у половины всех женщин, находящихся в периоде постменопаузы, а также у мужчин старших возрастных групп [1]. Очевидно, что рано начатые активные профилактические мероприятия у значительной части населения могут существенно повлиять на распространенность, прогрессирование и исходы заболевания, а также снизить риск переломов. В связи с этим изучение различных лекарственных препаратов и методов, применяемых для профилактики ОП, приобретает особый смысл.
Кость — специализированная разновидность соединительной ткани, состоящая из клеток и межклеточного вещества. В течение всей жизни основные функции костной ткани, такие как жесткость и гибкость, снижаются, поскольку с возрастом наблюдаются повреждение матрикса и потеря минералов. В противовес указанным проявлениям, в кости осуществляется ремоделирование — процесс, направленный на самостоятельное обновление и сохранение скелета как структурного и функционального органа.
Основными клетками костной ткани, функциями которой регулируется гомеостаз кости, являются остеобласты, остеокласты и остеоциты. Основной функцией остеобластов является создание органического межклеточного матрикса кости, остеоида. Остеобласты синтезируют и выделяют в окружающую среду фибриллы коллагена, протеогликаны и гликозаминогликаны. Наряду с этим остеобласты активно синтезируют и выделяют во внеклеточное пространство значительное количество глицерофосфолипидов, способствующих связыванию Ca 2+ и участвующих в процессах минерализации. Клетки сообщаются между собой через десмосомы, которые позволяют проходить Ca 2+ и цАМФ. Они также обеспечивают непрерывный рост кристаллов гидроксиапатитов и выступают в качестве посредников при связывании минеральных кристаллов с белковой матрицей.
В ходе формирования кости некоторые остеобласты оказываются замурованными в толщу матрикса и становятся остеоцитами. Остеоциты контактируют друг с другом через отростки, являются основными компонентами в сформировавшейся костной ткани. Основная функция остеоцитов — поддержание нормального состояния костного матрикса и баланса кальция и фосфора в организме.
Остеокласты — клетки, выполняющие функцию разрушения кости; развиваются из стволовой кроветворной клетки и являются специализированными макрофагами. В процессе ремоделирования кости резорбтивный стимул запускает процесс привлечения остеокластов к участку кости. Прикрепившись к кости, остеокласты продуцируют множество протеолитических ферментов и формируют полость в кальцинированном матриксе. Таким образом, они осуществляют непрерывный процесс резорбции и обновления костной ткани, обеспечивая необходимый рост и развитие скелета, структуру, прочность и упругость.
Важнейшим компонентом костной ткани является межклеточное вещество — уникальный комплекс органических и неорганических компонентов, заполняющих пространство между клетками. Минерализованный матрикс костной ткани поддерживает структуру скелета и под координирующим влиянием остеобластов и остеокластов обеспечивает резервуар как ионов, так и факторов роста, которые высвобождаются в процессе метаболизма.
Органический межклеточный матрикс костной ткани представлен семейством коллагеновых белков. Состав кости необычен тем, что фактически в ней представлен только коллаген I типа (90%), хотя наряду с коллагеном I типа в кости все же присутствуют следы других типов коллагена, таких как V, XI, XII. Скорее всего, что эти типы коллагена принадлежат другим тканям, которые и находятся в костной ткани, но не входят в состав костного матрикса. Например, коллаген V типа обычно обнаруживается в сосудах, которые пронизывают кость. Коллаген XI типа находится в хрящевой ткани и может соответствовать остаткам кальцифицированного хряща. Коллагеновые фибриллы в кости строго ориентированы в соответствии с распределенной функциональной нагрузкой на кость, что обеспечивает упругость и эластичность кости. Веретенообразные и пластинчатые кристаллы гидроксиапатита находятся на коллагеновых волокнах, в их пределах и в окружающем пространстве. Как правило, они ориентированы в том же направлении, что и коллагеновые волокна.
Неколлагеновая часть матрикса (10%) представлена основным веществом (витамин К-зависимыми глютамилпротеинами (остеокальцином), матричными протеинами, остеопонтином, остеонектином, фибронектином, фосфопротеидами, сиалопротеидами, а также протеогликанами).
Минеральные вещества, которыми пропитан органический матрикс, представлены главным образом кристаллами гидроксиапатита Ca10(PO4)6(OH)2. Кроме того, в кости обнаружены ионы Mg 2+ , Na + , K + , SO4 2- , HCO 3- , гидроксильные и другие ионы, которые могут принимать участие в образовании кристаллов.
Важно подчеркнуть, что ОП является результатом уменьшения органического матрикса кости, а вовсе не плохой кальцификацией костной ткани. При ОП существенно снижается скорость образования остеоида, необходимого для формирования кости. Поэтому при планировании профилактических мероприятий чрезвычайно важно учитывать потенциальную возможность препаратов, наряду с адекватной минерализацией, оказывать влияние на синтез органического матрикса.
Разумеется, качественная структура и прочность кости, ее эффективное функционирование и своевременное самообновление возможны лишь при адекватной обеспеченности макро- и микроэлементами, которые, подобно кальцию и витамину D, принимают непосредственное участие в биохимических процессах костной ткани [2–5]. Магний, медь, цинк, марганец, бор, являясь кофакторами ферментов, регулируют синтез костного матрикса, его минерализацию, а также равномерный рост, гибкость и прочность костной ткани. Известно, что дефицит этих веществ замедляет формирование костной массы в детстве и подростковом возрасте, способствует ее ускоренной потере в пожилом возрасте. Соответственно, дефицит любого из известных минеральных веществ в организме препятствует успешной терапии и профилактике нарушений структуры кости [6, 7].
Одним из основных минералов, играющих важную роль в формировании и поддержании структуры костной ткани, является кальций. Поскольку кальций не производится в организме, то для поддержания оптимальной концентрации он должен регулярно поступать извне. Причем желательно, чтобы его поступление в организм обеспечивалось за счет натуральных молочных продуктов, молока и его производных (кефира, простокваши, ряженки, йогурта, творога, сыра). Вместе с тем биодоступность кальция из пищи составляет порядка 30%, причем с высокой индивидуальной вариабельностью. Более того, у лиц пожилого возраста нередко имеет место непереносимость молочных продуктов, связанная со снижением концентрации лактазы в желудочном соке, что приводит к низкому потреблению кальция.
Согласно эпидемиологическим исследованиям, среди женщин в возрасте старше 45 лет, проживающих в мегаполисах, непереносимость молока встречается с частотой 25,0–34,0%. При этом достаточное потребление кальция с продуктами питания имеет место менее чем у 5% женщин [8]. Фактически содержание кальция в пищевом рационе постменопаузальных женщин не соответствует рекомендованным нормам. Очевидно, что обеспечение должного уровня потребления кальция возможно лишь при условии дополнительного регулярного назначения медикаментозных препаратов.
Витамин D — основной регулятор активной абсорбции кальция в организме. Витамин D относят к группе жирорастворимых витаминов. Хотя в отличие от всех других витаминов он биологически не активен. В активную, гормональную, форму он превращается за счет двухступенчатой метаболизации в организме и оказывает многообразные биологические эффекты за счет взаимодействия со специфическими рецепторами, локализованными в ядрах клеток тканей и органов. Другое дело — активный метаболит витамина D. Он действует как истинный гормон, хотя в научной литературе его традиционно называют витамином D [9, 10].
Природная форма витамина D — витамин D2 (эргокальциферол) поступает в организм человека в относительно небольших количествах — не более 20–30% от потребности. В основном из злаковых растений, рыбьего жира, сливочного масла, маргарина, молока, яичного желтка и др. В организме витамин D2 метаболизируется с образованием производных, обладающих сходным с метаболитами витамина D3 действием.
Еще одна природная форма витамина D — витамин D3, или холекальциферол, является ближайшим аналогом витамина D2, но его синтез мало зависит от поступления извне. Холекальциферол образуется в организме позвоночных животных, в том числе амфибий, рептилий, птиц и млекопитающих, в связи с чем играет значительно бóльшую роль в процессах жизнедеятельности человека, чем поступающий в небольших количествах с пищей витамин D2. В организме витамин D3 образуется из находящегося в дермальном слое кожи предшественника (7-дегидрохолестерина) под влиянием коротковолнового ультрафиолетового облучения спектра В (УФ–В/солнечного света, длина волны 290–315 нм) при температуре тела в результате фотохимической реакции раскрытия В-кольца стероидного ядра и термоизомеризации, характерной для секостероидов [9, 10].
В последующем поступивший с пищей и/или образовавшийся в организме в процессе эндогенного синтеза витамин D подвергается реакции 25-гидроксилирования в печени. Важно, что гидроксилирование витамина D3 в печени представляет собой полностью субстратзависимый процесс, который протекает весьма быстро и ведет к повышению уровня 25(ОН)D в сыворотке крови. Уровень этого вещества отражает как образование витамина D в коже, так и его поступление с пищей, в связи с чем может использоваться как маркер статуса витамина D [9, 10].
Вторая реакция гидроксилирования 25(ОН)D, с образованием наиболее важной, качественно и количественно значимой активной гормональной формы — 1a,25-дигидроксивитамина D3 (1α,25(ОН)2D3), называемой также D-гормоном, кальцитриолом, протекает уже в основном в почках, в клетках проксимальных отделов канальцев коры почек при участии фермента 1α-гидроксилазы (CYP27В1). Этот процесс строго регулируется рядом эндогенных и экзогенных факторов. Во-первых, регуляция синтеза 1a,25(ОН)2D3 в почках является непосредственной функцией паратиреоидного гормона (ПТГ), на концентрацию которого в крови, в свою очередь, по механизму обратной связи оказывают влияние как уровень самого активного метаболита витамина D3, так и концентрация кальция и фосфора в плазме крови. Во-вторых, активация синтеза 1a-гидроксилазы и реакции 1a-гидроксилирования зависит от половых гормонов (эстрогенов и андрогенов), кальцитонина, пролактина, гормона роста (через ИПФР-1) и др. В-третьих, ингибирующее влияние на активность 1a-гидроксилазы оказывают глюкокортикостероидные гормоны, 1α,25(ОН)2D3 и ряд его синтетических аналогов. Фактор роста из фибробластов (FGF23), секретируемый в клетках кости, вызывает образование натрий-фосфат-котранспортера, который действует в клетках почек и тонкого кишечника, оказывает тормозящее влияние на синтез 1,25-дигидроксивитамина D3. На метаболизм витамина D оказывают влияние и некоторые лекарственные средства, например, противоэпилептические препараты.
Основными реакциями, в которых участвует D-гормон, являются абсорбция кальция в желудочно-кишечном тракте и его реабсорбция в почках. D-гормон усиливает кишечную абсорбцию кальция в тонком кишечнике за счет взаимодействия со специфическими РВD. Об эффективности данного механизма свидетельствует тот факт, что без участия витамина D лишь 10–15% пищевого кальция и 60% фосфора абсорбируются в кишечнике. Взаимодействие между 1a,25-дигидроксивитамином D3 и РВD повышает эффективность кишечной абсорбции Са 2+ до 30–40%, т. е. в 2–4 раза, а фосфора — до 80%. Сходные механизмы действия D-гормона лежат в основе осуществляемой под его влиянием реабсорбции Са 2+ в почках.
В костях 1α,25(ОН)2D3 связывается с рецепторами на кость-формирующих клетках — остеобластах, вызывая повышение экспрессии ими лиганда рецептора активатора ядерного фактора кВ (RANKL). Рецептор-активатор ядерного фактора кВ (RANK), являющийся рецептором для RANKL, локализованным на преостеокластах, связывает RANKL, что вызывает быстрое созревание преостеокластов и их превращение в зрелые остеокласты. В процессах костного ремоделирования зрелые остеокласты резорбируют кость, что сопровождается выделением кальция и фосфора из минерального компонента (гидроксиапатита) и обеспечивает поддержание уровня кальция и фосфора в крови. В свою очередь, адекватный уровень кальция (Са 2+ ) и фосфора необходим для нормальной минерализации скелета [11–13].
Многочисленные исследования показали, что назначение препаратов кальция и/или витамина D способствует уменьшению потери костной ткани [14–19]. У женщин в поздней постменопаузе с низким употреблением пищевого кальция прием кальция предотвращает потерю костной ткани в позвоночнике [20, 21]. В свою очередь, назначение добавок кальция лицам старше 60 лет приводит к снижению потери костной массы в области бедра среди белых мужчин и женщин в возрасте моложе 72 лет [22]. Эффект назначения цитрата кальция на минеральную плотность кости (МПК) у женщин в раннем (до 5 лет) и среднем (от 5 до 10 лет) постменопаузальном периоде в течение двух лет проявлялся в виде прироста МПК в поясничном отделе на 1%, наряду со значимым снижением МПК на 2,4% в группе, получавшей плацебо [23]. Метаанализ 9 рандомизированных клинических исследований с общей выборкой более 50 тыс. человек, в 6 из которых сравнивалось комбинированное лечение витамином D (400 или 700–800 МЕ/сут) и кальцием с группами плацебо или без лечения, продемонстрировал достоверное снижение риска перелома бедра на 18% (RR 0,82 [95% ДИ 0,71–0,94], р = 0,0005) и риска внепозвоночных переломов на 12% (RR 0,88 [95% ДИ 0,78–0,99], р = 0,036) в группах, получавших комбинированную терапию, по сравнению с группами без добавок [24]. В исследованиях, где применялся витамин D в дозе 700–800 МЕ/сут, эффект на риск перелома бедра был выше, чем при приеме 400 МЕ (21% и 18% соответственно). Соответственно, в исследованиях, в которых пациенты получали только витамин D или плацебо (4 РКИ с общей численностью 9083 пациента), не было получено снижения риска внепозвоночных переломов как при применении дозы 400 МЕ (RR 1,14 [95% ДИ 0,87–1,49]), так при использовании 700–800 МЕ (RR 1,04 [95% ДИ 0,75–1,46]), что подтверждает ранее представленные данные о том, что витамин D без добавления кальция не снижает риск переломов [24].
Известно, что 60–65% магния находится именно в скелете и от обеспеченности костей магнием зависит обмен кальция и витамина D. Являясь структурным компонентом значительного числа ферментов, магний образует кристаллы с фосфатами, принимает участие в росте и стабилизации кристалла гидроксиапатита — структурной единицы минерального компонента костной ткани [25, 26]. Магний регулирует секрецию паратгормона (ПГ), повышает чувствительность клеток-мишеней к ПГ и витамину D, стимулирует действие кальцитонина. Длительное во времени нарушение соотношения Mg/Ca в сторону дефицита магния сопровождается замедлением обменных процессов в кости. Специальные магний-дефицитные диеты, сопровождающиеся уменьшением сывороточной концентрации магния, способствуют системной потере костной массы, снижению толщины надкостницы, характерным изменениям провоспалительных маркеров и маркеров резорбции кости. Уже по истечении достаточно короткого срока (4 недели), магний-дефицитная диета приводит к значимому снижению содержания минеральных веществ кости (р
М. И. Шупина, кандидат медицинских наук
Г. И. Нечаева 1 , доктор медицинских наук, профессор
Д. В. Шупин
Е. В. Надей
А. А. Семенкин, доктор медицинских наук, профессор
ГБОУ ВПО ОмГМУ МЗ РФ, Омск
Кость — структура, подвергающаяся непрерывным перестройкам на протяжении всей жизни. Прочность костей обеспечивает функционирование опорно-двигательного аппарата. Кроме того, костная ткань служит резервуаром, поддерживающим гомеостаз кальция, магния, фосфата, натрия и других ионов. Кости пронизаны большим количеством сосудов; костный кровоток составляет до 10% системного.

Свойства костной ткани определяются межклеточными компонентами — минеральным веществом и органическим матриксом.
На долю коллагена I типа приходится 90—95% всего белка органического матрикса.
К неколлагеновым белкам матрикса относятся: белки плазмы (альбумин и фетуин), белки, содержащие остатки γ-карбоксиглутаминовой кислоты (остеокальцин и γ-карбоксиглутамат-содержащий белок матрикса), гликопротеид остеонектин, фосфопротеид остеопонтин, сиалопротеиды, тромбоспондин и другие, менее изученные белки.
Некоторые из этих белков участвуют в минерализации матрикса. Минеральное вещество костной ткани состоит из частично кристаллизованного гидроксиапатита, в котором молярное отношение кальция к фосфату меньше, чем в чистом гидроксиапатите (формула чистого гидроксиапатита Са10(РО4)6(ОН)2).
За образование кости отвечают остеобласты — клетки мезенхимного происхождения, синтезирующие белки органического матрикса. Для остеобластов характерны:
1) определенные локализация и морфология;
2) присутствие костного изофермента ЩФ;
3) наличие рецепторов ПТГ и 1,25(OH)2D3;
4) способность синтезировать белки матрикса, такие, как коллаген I типа, остеокальцин и остеопонтин.
Минерализация новосинтезированного матрикса начинается сразу после его секреции (первичная минерализация) и продолжается в течение нескольких недель (вторичная минерализация). Остеобласт, окруженный минерализующимся матриксом, превращается в остеоцит, который сохраняет связь с другими клетками через систему канальцев.
Резорбция костной ткани осуществляется остеокластами — многоядерными клетками, образованными слиянием моноцитов.
В остеокластах обнаружены:
1) Н+-АТФаза и карбоангидраза II, обеспечивающие закисление среды в зоне резорбции;
2) тирозинкиназа Src, продукт онкогена SRC, а также ее субстрат р80/85;
3) устойчивая к тартрату кислая фосфатаза;
4) мембранные рецепторы кальцитонина;
5) Nа+,К+-АТФаза, сходная с аналогичным почечным ферментом;
6) Сl/HCO3-обменник, относящийся к семейству белков полосы 3. Остеокласты прикрепляются к белкам костного матрикса, например остеопонтину, с помощью аγβз-интегрина. При этом каждый остеокласт образует на поверхности кости зону прикрепления в виде кольца; цитоплазма остеокласта в области прикрепления светлая (светлая зона), и в ней содержатся сократительные белки.
Внутри этого кольца мембрана остеокласта образует так называемую гофрированную каемку; в ограниченном пространстве между гофрированной каемкой и костью и создается кислая среда, происходит растворение минерального вещества, а затем под действием кислых гидролаз рассасывается органический матрикс. В результате разрушительной деятельности остеокластов в кости образуются как бы разъеденные полости — гаушиповы, или резорбционные лакуны. В этих лакунах и располагаются остеокласты.
Описано много гуморальных регуляторов образования остеобластов и остеокластов из клеток-предшественников, а также функции дифференцированных клеток. Из них наиболее известны ИЛ-6, ИЛ-11 и колониестимулирующие факторы.
В костной ткани содержится много факторов роста. На функцию остеобластов действуют: трансформирующий фактор роста р типов I и II, кислый и основный факторы роста фибробластов, тромбоцитарный фактор роста, ИФР-I и ИФР-II. В перестройке кости участвуют, по-видимому, остеоглицин и ряд белков — регуляторов морфогенеза кости. Функцию остеокластов и процесс резорбции кости регулируют: ИЛ-1, ФНО, интерферон у и колониестимулирующие факторы.
В ряде случаев регуляция функции остеокластов осуществляется опосредованно, через остеобласты или фибробласты стромы костного мозга, — так действует, в частности, ПТГ, рецепторов к которому в зрелых остеокластах нет. Через взаимодействие с рецепторами в клетках-предшественниках l,25(OH)2D3 стимулирует их дифференцировку в моноциты и затем в остеокласты.
Некоторые цитокины, например ИЛ-1 и трансформирующий фактор роста а, индуцируют локальное образование простагландинов и других цитокинов, таких, как ИЛ-6 и колониестимулирующие факторы. Так называемые факторы активации остеокластов, как теперь очевидно, — это группа цитокинов, в том числе ИЛ-1, ФНОα и β, а также, возможно, другие вещества.
У плода и ребенка кость образуется путем замещения обызвествленного хряща (хрящевой остеогенез) или формируется непосредственно из мезенхимы (перепончатый остеогенез).
В новообразованной костной ткани (у детей, а также у взрослых при быстром росте, например при образовании костной мозоли) относительно много клеток и мало матрикса, для нее характерно беспорядочное расположение толстых переплетающихся пучков коллагеновых нитей (ретикулофиброзная кость).
В зрелой кости пучки коллагеновых нитей организованы в параллельные или концентрические пластинки (пластинчатая кость). В длинных костях костные пластинки, расположенные концентрически вокруг
кровеносных сосудов, формируют остеоны (гаверсовы системы). Толщина кости увеличивается, если скорость остеогенеза в надкостнице выше скорости резорбции на эндостальной поверхности. Удлинение костей происходит за счет пролиферации клеток эпифизарного хряща и последующего хрящевого остеогенеза.
У взрослых эпифизы зарастают, удлинение костей и хрящевой остеогенез прекращаются; некоторая активность клеток сохраняется только в суставном хряще. Однако и у взрослых продолжается перестройка как остеонов, так и губчатых костей. Новообразованная костная ткань отличается гладкой поверхностью, поглощает тетрациклин, имеет относительно низкое содержание минерального вещества и покрыта активными остеобластами. Толщина нового неминерализованного органического матрикса (остеоида) составляет около 12 мкм. Для определения скорости образования костной ткани больному дважды с некоторым интервалом дают тетрациклин, а затем измеряют расстояние между флюоресцирующими слоями на срезах костного биоптата.
Зоны резорбции отличаются неровной поверхностью и присутствием остеокластов. Резорбция предшествует остеогенезу, она протекает более интенсивно, но не столь долго, как последний. Если активная резорбция у взрослых происходит приблизительно на 4% поверхности губчатой кости (например, гребня подвздошной кости), то неминерализованным органическим матриксом покрыто 10—15% поверхности костных балок.
Изотопные исследования показывают, что ежегодно замещается до 18% всего кальция костей. Таким образом, в костной ткани происходит активный обмен веществ, требующий хорошего кровоснабжения. Перестройка костей каким-то образом зависит от механической нагрузки. Кроме того, костная ткань служит резервуаром неорганических ионов, например кальция, играющего важную роль во многих физиологических процессах.
Реакция костной ткани на повреждение (переломы, инфекцию, нарушение кровоснабжения, метастазы) весьма ограничена. Процесс замещения мертвой костной ткани на новую сопровождается прорастанием новых кровеносных сосудов в пораженную область.
При значительном нарушении структуры ткани, например при переломе со смещением костных отломков или их патологической подвижностью, стромальные клетки-предшественники дифференцируются не в остеобласты, а в клетки, формирующие соединительную ткань и хрящ.
При хорошем совмещении и фиксации костных отломков срастание осуществляется преимущественно за счет остеогенеза, и рубцовой деформации не происходит. Перестройка кости зависит от механической нагрузки, которая каким-то образом влияет на биологическую активность ткани.
На границе растущей опухоли с костью происходит резорбция последней. Прогибающие деформации стимулируют остеогенез на вогнутой и резорбцию на выпуклой поверхностях, что способствует сохранению механической прочности.
Даже при таком деструктивном заболевании, как болезнь Педжета, перестройка кости зависит от механических сил. Таким образом, пластичность костной ткани определяется взаимодействием клеток друг с другом и с внешней средой.
Механизмы остеогенеза и резорбции кости
Остеогенез — это упорядоченный процесс образования и минерализации органического матрикса кости. В состав кости входят кальций и фосфат, поэтому скорость минерализации зависит от концентрации этих ионов в плазме и внеклеточной жидкости.
В искусственных условиях для кристаллизации гидроксиапатита достаточно тех концентраций кальция и фосфата, которые имеются в плазме. Концентрация ионов в очагах минерализации неизвестна, очевидно только, что она регулируется остеобластами и остеоцитами.
Коллаген различного происхождения способствует возникновению центров кристаллизации, и минерализация начинается в определенных участках упорядоченной структуры матрикса — промежутках между молекулами коллагена. Степень и характер минерализации, по-видимому, зависят от организации коллагена.
Первичная структура коллагенов I типа из кожи и из кости практически одинакова, однако эти белки претерпевают различные постгрансляционные изменения и поэтому различаются по степени гидроксилирования, гликозилирования, по типу, числу и распределению межмолекулярных поперечных связей. Кроме того, размер промежутков в упорядоченной структуре матрикса больше в минерализованном коллагене кости и дентина, чем в неминерализованном коллагене, например, сухожилий.
Важность коллагенового матрикса для нормального развития кости доказывается тем, что замена единичных аминокислот в спиральной части α1 — или α2-цепей коллагена I типа (в результате мутаций генов COL1A1 или COL1A2 соответственно) приводит к выраженному нарушению структуры кости, что проявляется как несовершенный остеогенез.
Неколлагеновые белки — остеокальцин, остеонектин и остеопонтин — также участвуют в минерализации. ЩФ служит маркером остеобластов, и чем больше ее активность, тем выше остеогенная способность этих клеток. Р
оль ЩФ в процессе минерализации до конца не выяснена, однако при врожденной недостаточности ЩФ (гипофосфатазии) остеогенез нарушается. ЩФ при нейтральном pH способна гидролизовать неорганический пирофосфат — мощный ингибитор минерализации.
Возможно, что функция ЩФ в остеогенезе сводится к регуляции уровня пирофосфата. Процесс минерализации подавляют также макромолекулярные комплексы, например протеогликаны. При обызвествлении хряща минерализация начинается в мембранных внеклеточных везикулах.
Минерализация начинается с отложения брушита (СаНР04 х 2Н20). Затем появляется слабокристаллизованый гидроксиапатит с низким (около 1,2) молярным отношением кальция к фосфату. С возрастом степень кристаллизации и отношение кальция к фосфату увеличиваются. Включение анионов фтора снижает долю аморфного фосфата кальция и усиливает кристаллическую структуру.
Если внеклеточные концентрации кальция и фосфата ниже пороговых, то минерализации не происходит. Произведение растворимости для минерального вещества кости рассчитать очень трудно, так как его состав непостоянен, а растворимость зависит от других, пока неизвестных факторов внеклеточной жидкости. При избыточной концентрации кальция и фосфата во внеклеточной жидкости иногда наблюдается эктопическая минерализация.
При резорбции кости кальций и фосфат поступают во внеклеточную жидкость, а органический матрикс рассасывается. Для растворения минерального вещества необходима кислая среда, которая создается в ограниченном пространстве между остеокластом и поверхностью кости. ЩФ — фермент, выделяемый клетками кости во внеклеточную среду.
Стимуляция остеогенеза сопровождается увеличением в крови активности костного изофермента ЩФ, остеокальцина и С-концевого пептида проколлагена I типа. Маркеры костной резорбции в моче: гидроксипролин, гидроксилизин и его гликозиды, пиридинолин и дезоксипиридинолин.
Читайте также: