
Наличие атланта и эпистрофея у пресмыкающихся
Класс Пресмыкающиеся (рептилии) включает около 9000 ныне живущих видов, которых разделяют на четыре отряда: Чешуйчатые, Крокодилы, Черепахи, Клювоголовые. Последний представлен только одним реликтовым видом — гаттерией. К чешуйчатым относятся ящерицы (в том числе хамелеоны) и змеи.
Общая характеристика рептилий
Пресмыкающиеся считаются первыми настоящими наземными животными, так как они не связаны в своем развитии с водной средой. Если и живут в воде (водные черепахи, крокодилы), то дышат легкими и для размножения выходят на сушу.
Рептилии расселены по суше намного больше, чем амфибии, занимают более разнообразные экологические ниши. Однако из-за холоднокровности они преобладают в теплом климате. При этом могут обитать в засушливых местах.
Пресмыкающиеся появились от стегоцефалов (вымершая группа земноводных) в конце каменноугольного периода палеозойской эры. Черепахи появились раньше, а змеи позже всех.
Расцвет пресмыкающихся пришелся на мезозойскую эру. В это время на Земле жили различные динозавры. Среди них были не только наземные и водные виды, но и летающие. Динозавры вымерли в конце мелового периода.
В отличие от земноводных у пресмыкающихся
улучшена подвижность головы за счет большего количества шейных позвонков и иного принципа их соединения с черепом;
кожа покрыта роговыми чешуями, которые защищают тело от высыхания;
дыхание только легочное; формируется грудная клетка, которая обеспечивает более совершенный механизм дыхания;
хотя сердце остается трехкамерным, венозный и артериальный кровотоки разделены лучше, чем у земноводных;
в качестве органов выделения появляются тазовые почки (а не туловищные как у амфибий); такие почки лучше сохраняют воду в организме;
мозжечок крупнее, чем у амфибий; увеличен объем переднего мозга; появляется зачаток коры больших полушарий;
внутреннее оплодотворение; пресмыкающиеся размножаются на суше преимущественно путем откладки яиц (некоторые живородящие или яйцеживородящие);
появляются зародышевые оболочки (амнион и аллантоис).
Кожа пресмыкающихся
Кожа пресмыкающихся состоит из многослойного эпидермиса и соединительно-тканной дермы. Верхние слои эпидермиса ороговевают, образуя чешуи и щитки. Основное назначение чешуй — защита тела от потери воды. В общей сложности кожа более толстая, чем у амфибий.
Чешуя пресмыкающихся не гомологична чешуе рыб. Роговые чешуи образуются эпидермисом, т. е. имеет эктодермальное происхождение. У рыб чешуя образуется дермой, т. е. имеет мезодермальное происхождение.
В отличие от земноводных в коже пресмыкающихся слизистых желез нет, поэтому их кожа сухая. Есть только малочисленные пахучие железы.
У черепах на поверхности тела (сверху и снизу) образуется костный панцирь.
На пальцах появляются когти.
Так как ороговевшая кожа сдерживает рост, то для пресмыкающихся характерна линька. При этом старые покровы отходят от тела.
Кожа рептилий плотно срастается с телом, не образуя лимфатических мешков, как у амфибий.
Скелет пресмыкающихся
По-сравнению с земноводными у пресмыкающихся в позвоночнике выделяют уже не четыре, а пять отделов, так как туловищный отдел подразделяют на грудной и поясничный.
У ящериц шейный отдел состоит из восьми позвонков (у различных видов их от 7 до 10). Первый шейный позвонок (атлант) похож на кольцо. В него заходит зубовидный отросток второго шейного позвонка (эпистрофея). В результате первый позвонок может относительно свободно вращаться вокруг отростка второго позвонка. Это дает большую подвижность головы. Кроме того, первый шейный позвонок соединен с черепом одним мышелком, а не двумя как у земноводных.
Все позвонки грудного и поясничного отдела имеют ребра. У ящериц ребра первых пяти позвонков присоединяются хрящами к грудине. Образуется грудная клетка. Ребра задних грудных и поясничных позвонков не соединены с грудиной. Однако у змей грудины нет, и, следовательно, не образуется грудной клетки. Такое строение связано с особенностями их передвижения.
Крестцовый отдел позвоночника у пресмыкающихся состоит из двух позвонков (а не из одного как у амфибий). К ним прикрепляются подвздошные кости тазового пояса.
У черепах позвонки туловища срастаются со спинным щитом панциря.
Положение конечностей относительно тела — по бокам. У змей и безногих ящериц конечности редуцированы.
Пищеварительная система пресмыкающихся
Пищеварительная система пресмыкающихся сходна с таковой земноводных.
В ротовой полости находится подвижный мускулистый язык, у многих видов раздвоенный на конце. Пресмыкающиеся способны его далеко выбрасывать.
У растительноядный видов появляется слепая кишка. Однако большинство являются хищниками. Например, ящерицы питаются насекомыми.
Слюнные железы содержат ферменты.
Дыхательная система пресмыкающихся
Пресмыкающиеся дышат только легкими, так как из-за ороговения кожа в дыхании принимать участие не может.
Легкие совершенствуются, их стенки образуют многочисленные перегородки. Такое строение увеличивает внутреннюю поверхность легких. Трахея длинная, в конце делится на два бронха. У пресмыкающихся бронхи в легких не ветвятся.
У змей имеется только одно легкое (правое, а левое редуцировано).
Механизм осуществления вдоха и выдоха у пресмыкающихся кардинальным образом отличается от такового у земноводных. Вдох происходит при расширении грудной клетки за счет растягивания межреберных и брюшных мышц. При этом воздух всасывается в легкие. При выдохе мышцы сокращаются, и воздух выталкивается из легких.
Кровеносная система пресмыкающихся
Сердце подавляющего большинства пресмыкающихся остается трехкамерным (два предсердия, один желудочек), и артериальная и венозная кровь все еще частично смешиваются. Но по-сравнению с земноводными у пресмыкающихся венозный и артериальный кровотоки лучше разделены, и, следовательно, кровь смешивается меньше. В желудочке сердца имеется неполная перегородка.
Пресмыкающиеся (как земноводные и рыбы) остаются холоднокровными животными.
У крокодилов в желудочке сердца имеется полная перегородка, и таким образом образуется два желудочка (его сердце становится четырехкамерным). Однако кровь все еще может смешиваться через дуги аорты.
От желудочка сердца пресмыкающихся самостоятельно отходят три сосуда:
От правой (венозной) части желудочка отходит общий ствол легочных артерий, который далее разделяется на две легочные артерии, идущие к легким, где кровь обогащается кислородом и возвращается по легочным венам в левое предсердие.
От левой (артериальной) части желудочка отходят две дуги аорты. Одна дуга аорты начинается левее (однако называется правой дугой аорты, так как загибается направо) и несет почти чистую артериальную кровь. От правой дуги аорты берут начало сонные артерии, идущие к голове, а также сосуды, снабжающие кровью пояс передних конечностей. Таким образом, эти части тела снабжаются почти чистой артериальной кровью.
Вторая дуга аорты отходит не столько от левой части желудочка, сколько из его середины, где кровь смешанная. Эта дуга находится правее правой дуги аорты, но называется левой дугой аорты, так как по выходу загибается налево. Обе дуги аорты (правая и левая) на спинной стороне соединяются в единую спинную аорту, ответвления которой снабжают смешанной кровью органы тела. Оттекающая от органов тела венозная кровь попадает в правое предсердие.
Выделительная система пресмыкающихся
У пресмыкающихся в процессе эмбрионального развития туловищные почки заменяются тазовыми. Тазовые почки имеют длинные канальцы нефронов. Их клетки дифференцированы. В канальцах происходит обратное всасывание воды (до 95%).
Основной продукт выделения пресмыкающихся — мочевая кислота. Она почти не растворяется в воде, поэтому моча кашицеобразная.
От почек отходят мочеточники, впадающие в мочевой пузырь, который открывается в клоаку. У крокодилов и змей мочевой пузырь недоразвит.
Нервная система и органы чувств пресмыкающихся
Совершенствуется головной мозг пресмыкающихся. В переднем мозге появляется кора больших полушарий из серого мозгового вещества.
У ряда видов промежуточный мозг образует теменной орган (третий глаз), который способен воспринимать свет.
Мозжечок у пресмыкающихся лучше развит, чем у амфибий. Это связано с более разнообразной двигательной активностью рептилий.
Условные рефлексы вырабатываются с трудом. Основа поведения — инстинкты (комплексы безусловных рефлексов).
Глаза снабжены веками. Есть третье веко — мигательная перепонка. У змей веки прозрачные и срастаются.
У ряда змей на переднем конце головы есть ямки, воспринимающие тепловое излучение. Они хорошо определяют разницу между температурами окружающих предметов.
Орган слуха образует внутреннее и среднее ухо.
Хорошо развито обоняние. В ротовой полости есть специальный орган, различающий запахи. Поэтому многие пресмыкающиеся высовывают раздвоенный на конце язык, беря пробы воздуха.
Размножение и развитие рептилий
Для всех пресмыкающихся характерно внутреннее оплодотворение.
Большинство откладывают яйца в грунт. Бывает так называемое яйцеживорождение, когда яйца задерживаются в половых путях самки, и при выходе из них сразу вылупляются детеныши. У морских змей наблюдается настоящее живорождение, при этом у зародышей формируется плацента, сходная с плацентой млекопитающих.
Развитие прямое, появляется молодое животное, сходное по строению со взрослым (но с недоразвитой половой системой). Это связано с наличием большого запаса питательных веществ в желтке яйцеклетки.
В яйце пресмыкающихся формируются две эмбриональные оболочки, которых нет в икринках земноводных. Это амнион и аллантоис. Эмбрион окружен амнионом, наполненным амниотической жидкостью. Аллантоис образуется как вырост заднего конца кишечника эмбриона и выполняет функции мочевого пузыря и органа дыхания. Наружная стенка алантоиса прилегает к оболочке яйца и содержит капилляры, через которые осуществляется газообмен.
Забота о потомстве у пресмыкающихся редка, заключается в основном в охране кладки.

--> Соревнования по полевой ботанике "ВЕСЕННЯЯ ФЛОРА" пройдут в мае-июне 2020 года в онлайн-формате (определение растений по фотографиям). К участию в соревновании приглашаются школьники и взрослые любители природы, проживающие в средней полосе Европейской части России. Подробнее >>>
| [ sp ] : ml об : |
Наши авторские методические материалы по рептилиям и амфибиям России:
В нашем Интернет-магазине по некоммерческим ценам (по себестоимости производства)
можно приобрести следующие методические материалы по рептилиям Северной Евразии:
Анатомия, морфология и экология пресмыкающихся
4. Скелет пресмыкающихся
Позвоночный столб пресмыкающихся разделяется на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой.
Образование гибкой шеи и усиление подвижности головы имело первостепенное значение при добывании пищи и при ориентировке. Подвижность головы обеспечивается дифференцировкой первых двух шейных позвонков — атласа, или атланта (atlas), и эпистрофея (epistropheus). Атлас имеет вид костного кольца, разделенного плотной связкой на верхнюю и нижнюю половины; через верхнее отверстие головной мозг соединяется со спинным; передняя поверхность нижней половины сочленяется с затылочным мыщелком черепа, а сзади в нижнее отверстие входит зубовидный отросток второго шейного позвонка— эпистрофея. Голова может поворачиваться в стороны на зубовидном отростке, а ее перемещение в вертикальной плоскости обеспечивается сочленением черепного мыщелка с атласом. Все это обеспечивает сложные движения головы, усиливаемые подвижностью всей шеи. Изучение эмбрионального развития показало, что зубовидный отросток образуется путем прирастания к эпистрофею тела атласа.
Движения шеи обусловлены числом и строением позвонков шейного отдела; они различны в разных группах. У гаттерии позвонки еще амфицельные (рыбьего типа) с остатками хорды между ними. У крокодилов и большинства чешуйчатых позвонки процельные (передневогнутые) и лишь у немногих низших форм амфицельные. Часть шейных позвонков несет короткие ребра. У скрытошейных черепах, изгибающих шею в вертикальной плоскости, шейные позвонки сохраняют лишь зачатки поперечных отростков. Наоборот, у бокошейных черепах, изгибающих шею в сторону, поперечные отростки и связанная с ними мускулатура сильно развиты. Сложные движения шеи черепах обеспечиваются и разнообразием позвонков: задние позвонки процельны, передние — опистоцельны (задневогнутые), а средний — амфицельный.
К грудным позвонкам причленяются длинные ребра, брюшные концы которых с помощью хрящевых отделов прикрепляются к грудине, образуя замкнутую грудную клетку (грудной клетки нет у змей). К грудине же причленяется и плечевой пояс. Поясничные позвонки тоже несут ребра, которые не доходят до грудины. К крестцовому отделу, состоящему из двух позвонков, прикрепляется тазовый пояс. Хвостовой отдел помогает сохранять равновесие при передвижении, а иногда служит и движителем (у водных змей, крокодилов и некоторых водных ящериц).
У ящериц, способных к аутотомии (отбрасыванию хвоста), хвостовые позвонки могут разламываться посредине, где имеются тонкие хрящевые прослойки, делящие тело позвонка на две части.
Общее число позвонков различно у разных видов и достигает 50-80 (7-10 шейных, 16-25 грудино-поясничных, 2 крестцовых, 15-40 хвостовых). У змей и безногих ящериц число позвонков увеличивается, а позвоночник делится только на туловищный и хвостовой отделы. Все туловищные позвонки снабжены подвижными ребрами, упирающимися в брюшные щитки, что важно при змееобразном движении. Общее число позвонков возрастает до 140 (у толстых и коротких змей) — 435 (у змей с длинным телом).
Парные конечности и их пояса
Плечевой пояс пресмыкающихся сходен с поясом земноводных, но в нем сильно развиты окостенения. Коракоид (coracoideum) в месте соединения с лопаткой (scapula) образуют суставную впадину для причленения головки плеча (рис. 3).
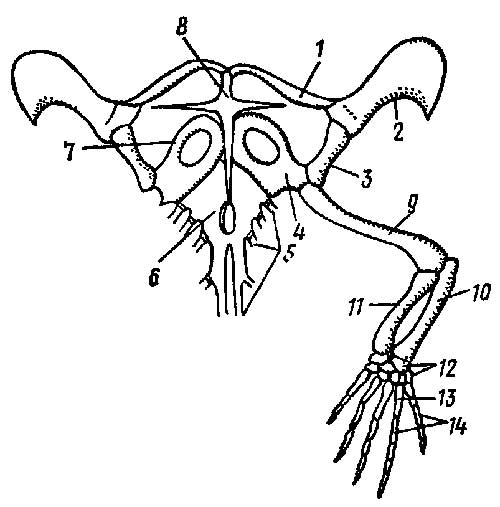
Рис. 3. Плечевой пояс и передняя конечность ящерицы Lacerta: 1 — ключица, 2 — надлопаточный хрящ, 3 — лопатка, 4 — коракоид, 5 — ребра, 6.— грудина, 7 — прокоракоидный хрящ, 8 — надгрудинник, 9 — плечо, 10 — локтевая кость, 11 — лучевая кость, 12 — запястье, 13 — пясть, 14 — фаланги пальцев
Сверху к лопатке прирастает уплощенный надлопаточный хрящ (cartilage suprascapularis), а спереди к коракоиду — хрящевой прокоракоид (cart. procoracoidea). Коракоид и прокоракоид каждой стороны срастаются с непарной костной грудиной (sternum); через грудную клетку пояс передних конечностей крепится к осевому скелету. Снизу к грудине прирастает крестообразная покровная кость — надгрудинник (episternum). Парные покровные ключицы (clavicula) соединяют передний конец надгрудинника с дорзальной частью каждой лопатки.
Такая конструкция усиливает прочность плечевого пояса. У черепах надгрудинник и ключицы входят в брюшной щит панциря; у крокодилов хорошо развиты лишь коракоиды и лопатки.
Тазовый пояс состоит из двух безымянных костей; каждая из них образуется слиянием трех тазовых костей — подвздошной (ilium), седалищной (ischium) и лобковой (pubis), совместно образующих вертлужную впадину, составляющую сустав с головкой бедра (рис. 14).
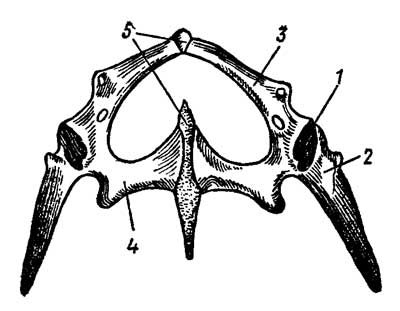
Рис. 4. Тазовый пояс ящерицы Lacerta: 1 — суставная впадина для головки бедра, 2 — подвздошная, 3 — лобковая, 4 — седалищная кости, 5 — симфиз
Подвздошные кости сочленяются с поперечными отростками крестцовых позвонков. У всех современных пресмыкающихся таз закрытый: правые и левые лобковые и седалищные кости соединяются друг с другом по средней линии симфизом — хрящевой перемычкой.
Парные конечности отличаются у разных видов и групп пресмыкающихся в зависимости от преобладания тех или иных способов передвижения. Но обычно они сохраняют общую схему строения парных конечностей наземных позвоночных.
В отличие от земноводных у пресмыкающихся в передней конечности подвижный сустав расположен между двумя рядами косточек запястья (интеркарпальный сустав), а в задней конечности — между рядами косточек предплюсны (интертарзальный сустав).
Череп пресмыкающихся видоизменялся главным образом в зависимости от характера питания и способов добывания пищи. От черепа амфибий он отличается вытянутыми челюстями, образующими относительно длинное рыло; широкий и короткий череп амфибий был необходим при их ротоглоточном механизме дыхания; в то же время широкий рот способствовал захвату мелкой добычи при броске на нее. У пресмыкающихся дыхание обеспечивается работой грудной клетки, а захват добычи связан с активным преследованием, при котором вытянутое рыло имеет преимущество. Такая форма челюстей давала к тому же возможность отрывать куски от крупной добычи. То и другое требовало более мощной жевательной мускулатуры. С ее развитием и усложнением органов чувств связаны основные изменения черепа пресмыкающихся.
Череп почти полностью окостеневает (рис. 5).
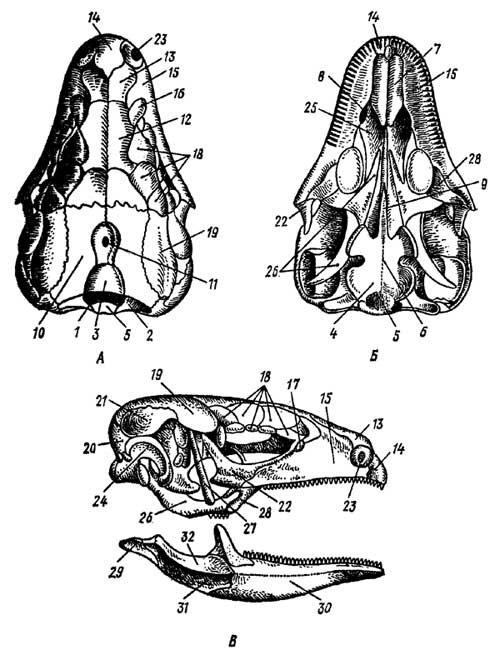
Рис. 5. Череп ящерицы Lacerta (по Паркеру). А — вид сверху; Б — вид снизу, В — вид сбоку
1 — затылочное отверстие, 2 — боковая затылочная кость, 3 — верхняя затылочная кость, 4 — основная затылочная кость, 5 — затылочный мыщелок, 6 — основная клиновидная кость, 7 — сошник, 8 — хоана, 9 — парасфеноид, 10 — теменная кость, 11 — межтеменная кость с отверстием для теменного органа, 12 — лобная кость, 13 — носовая кость, 14 — межчелюстная кость, 15 — верхнечелюстная кость, 16 — предлобная кость, 17 — слезная кость, 18 — надглазничные кости, 19 — заднелобная, или заглазничная кость, 20 — чешуйчатая кость, 21 — надвисочная кость, 22 — скуловая кость, 23 — ноздря, 24 — квадратная кость, 25 — небная кость, 26 — крыловидная кость, 27 — верхнекрыловидная, или столбчатая кость, 28 — поперечная кость, 29 — сочленовная кость, 30 — зубная кость, 31 — надугловая кость, 32 — венечная кость
Затылочная область состоит из четырех затылочных костей (occipitalia) хондрального происхождения: верхнезатылочной, основной и двух боковых. Они окаймляют затылочное отверстие, книзу от которого лежит единственный затылочный мыщелок, образованный основной и обеими боковыми затылочными костями. Покровная основная клиновидная кость (basisphenoideum) лежит впереди основной затылочной, образуя дно черепа. Впереди к ней прирастает небольшой парасфеноид (parasphenoideum) и располагаются парные сошники (vomer), сбоку от которых лежат хоаны. В области слуховой капсулы возникают три ушные кости (otici); переднеушная, сохраняющая самостоятельность; заднеушная, срастающаяся с боковой затылочной, и верхнеушная, срастающаяся с верхнезатылочной. В обонятельной области костей нет; она остается хрящевой.
Крыша черепа образована парными покровными костями: носовыми (nasalia), предлобными (praefrontalia), лобными (frontalia) и заднелобными (postfrontalia); далее лежат теменные (parietalia) и непарная межтеменная (interparietale) кости; последняя имеет отверстие для теменного органа.
Бока черепа образуют покровные кости: парные межчелюстные (intermaxillare) у части видов сливающиеся, парные верхнечелюстные (maxillare), надглазничные (supraorbitale), скуловые (jugale), квадратно-скуловые (quadratojugale) (квадратно-скуловая кость есть в анапсидном и типичном диапсидном черепе гаттерий и крокодилов, но исчезает у ящериц, имеющих диапсидный тип черепа с редуцированной нижней дугой) и чешуйчатые (squamosum). Небно-квадратный хрящ в задней части дает начало парным хондральным окостенениям — квадратным костям (quadratum), верхним отделом связанным с мозговой коробкой, а нижним — с нижней челюстью.
Передняя часть нёбно-квадратного хряща заменяется покровными костями, образующими дно черепа: парными небными (palatini) и крыловидными (pterygoidei). Поперечные кости (transversi) соединяют крыловидные кости с верхнечелюстными, а у ящериц и гаттерий еще и верхнекрыловидные, или столбчатые (epipterygoidei), кости связывают крыловидные кости с теменными.
У черепах и особенно у крокодилов разрастанием небных отростков межчелюстных и верхнечелюстных костей, а также небных костей образуется вторичное костное небо, разделяющее ротовую полость на верхний отдел — носоглоточный, и нижний — собственно ротовую полость. Поэтому хоаны сдвигаются назад, к гортани, что позволяет дышать, когда из воды выставлен лишь конец головы с ноздрями.
Нижняя челюсть образуется из меккелева хряща, замещаемого хондральной костью — сочленовной (articulare), причленяющейся к квадратной кости, и ряда кожных костей: зубной (dentale), угловой (angulare), надугловой (supraangulare), венечной (coronare) и иногда еще нескольких косточек.
Верхний отдел подъязычной дуги (гиомандибуляре), как и у земноводных, превращен в слуховую косточку среднего уха — стремечко (stapes).
Подъязычный аппарат состоит из хрящевой пластинки (гомолога копулы) и трех пар рожков, гомологичных гиоидам и остаткам жаберных дуг.
Эволюция жевательной мускулатуры сопровождалась перестройкой крыши и частично боковых стенок первичносплошного — стегального — черепа, имевшегося у земноводных — предков рептилий. В отдельных ветвях пресмыкающихся эволюция шла разными путями. В группе анапсида (ископаемые котилозавры и их непосредственные потомки — черепахи) первичная стегальная (сплошная) крыша черепа сохранялась. В ней образовалась лишь задняя вырезка, освободившая место для жевательной мускулатуры. В группе диапсида (гаттерия, крокодилы) образовались две височные ямы (рис. 6), ограниченные двумя костными дугами.
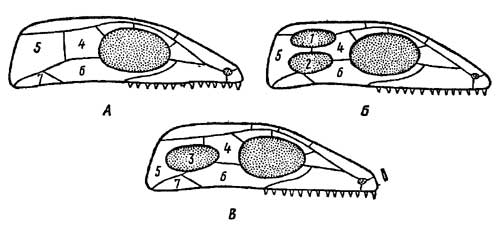
Рис. 6. Схема эволюционных преобразований стегального черепа у пресмыкающихся (по Гудричу). А—стегальный череп; с изменениями свойствен анапсидам; Б — диапсидный череп с двумя височными ямами; В — синапсидный череп с одной боковой височной ямой: 1 — верхняя височная яма, 2 — нижняя височная яма, 3 — единственная боковая височная яма, 4 — заднелобная (заглазничная) кость, 5 — чешуйчатая кость, 6 — скуловая кость, 7 — квадратно-скуловая кость
У ящериц диапсидный тип черепа с редуцированной нижней дугой, а у птиц — с редуцированной верхней дугой; у змей обе дуги редуцируются. В группе синапсид — у звероящеров (тероморфных рептилий) и происшедших от них млекопитающих — образовалась одна боковая яма и ограничивающая ее сложная дуга из элементов верхней и нижней дуг: jugale — quadratojugale — squamosum. Строение скуловых дуг еще раз подчеркивает своеобразие группы звероподобных пресмыкающихся, противопоставляя их другим группам этого класса.
Параллельно усложнению скелета изменилась структура костной ткани. Общий тип трубчатых костей конечностей сохранился, но грубоволокнистая костная ткань земноводных заместилась тонковолокнистой костью, обладающей значительно большей прочностью. При этом кости конечностей приобрели остеонную структуру и зонарное (слоистое) строение.
Костный элемент — остеон — представляет собой костную трубочку, стенки которой образованы тонкими, но прочными костными пластинками, а полость занята питающим костную ткань кровеносным сосудом и нервом. Остеоны в стенках трубчатой кости располагаются более или менее концентрически, образуя зонарную структуру. Внутренняя полость трубчатой кости занята костным мозгом двух типов: красный выполняет функцию кроветворной ткани, желтый участвует в костеобразовании, разрушая кость со стороны внутренней полости. Обратный процесс — нарастание кости в толщину — происходит с наружной поверхности и обеспечивается ее оболочкой — периостом. Рост трубчатых костей в длину у рептилий происходит в средней части трубки (у млекопитающих в местах пере хода трубки — диафиза в сочленовые головки —эпифизы) (рис. 7).
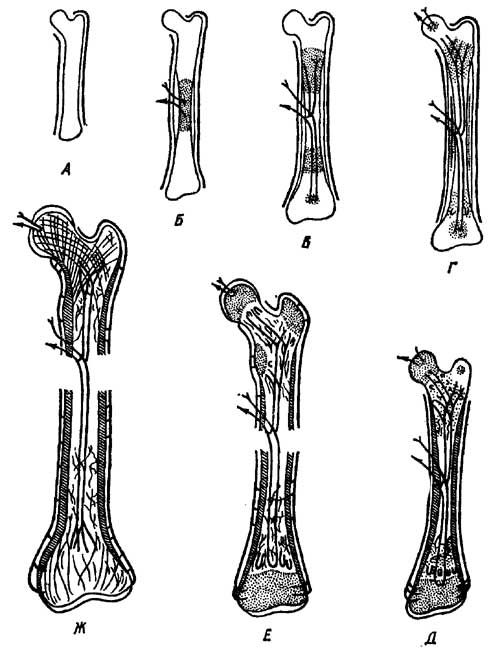
Рис. 7. Развитие трубчатой кости пресмыкающихся. А — хрящевой зачаток; Б-Ж — последовательные стадии развития периостального и эндохоидрального окостенений (Заварзин и Румянцев, 1946).
Кость такого строения при одинаковом весе и размерах может выдержать значительно большее напряжение, нежели кость земноводных. За счет совершенствования костной ткани, не увеличив веса скелета, пресмыкающиеся могут выполнять большую работу. Это увеличило их подвижность. Уменьшение относительной массы скелета позволило перейти даже к полету, чему способствовала пневматизация костей, внутренние полости которых у птерозавров частично заполнялись воздухом.
Мускулатура пресмыкающихся, по сравнению с земноводными, претерпела существенные перестройки. Исходное метамерное расположение туловищной мускулатуры, характерное для рыб, в большей части утеряно уже земноводными. Сложная дифференциация продолжалась и у рептилий. Метамерность сохранилась лишь в размещении мышц, соединяющих рядом расположенные позвонки, и отчасти в мускулатуре брюшной стенки.
На черепе в височных ямках развивается мощная жевательная мускулатура, приводящая в движение челюсти. Появление подвижного шейного отдела сопровождалось развитием шейной мускулатуры, особенно мощной у видов с длинной и гибкой шеей. Лентовидные мышцы туловищной мускулатуры участвуют в изгибании тела и в движении конечностей. Непосредственно на последних размещена тоже мощная мускулатура, особенно парные сгибатели и разгибатели. Их совокупная работа обеспечивает разнообразную и во многих случаях весьма сложную работу конечностей. Появляются зачатки подкожной мускулатуры, изменяющей положение роговых чешуи. Заметно сильнее она развита у змей и безногих ящериц и участвует в перемещении тела, обеспечивая координированные движения брюшных щитков.
В связи с переходом рептилий полностью на легочное дыхание потребовалось усиление вентиляции легких. Последнее было обеспечено возникновением грудной клетки, приводимой в движение специальной межреберной мускулатурой. На изменении объема грудной полости сказывается и движение конечностей (особенно у черепах). Быстрое передвижение требовало стабилизации положения внутренних органов. Это было достигнуто значительным усилением мышц брюшного пресса.
Наши авторские методические материалы по рептилиям и амфибиям России:
В нашем Интернет-магазине по некоммерческим ценам (по себестоимости производства)
можно приобрести следующие методические материалы по рептилиям Северной Евразии:
В разделе Методические материалы Вы также можете познакомиться с описаниями разработанных экологическим центром "Экосистема" печатных определителей растений средней полосы, карманных определителей объектов природы средней полосы, определительных таблиц "Грибы, растения и животные России", компьютерных (электронных) определителей природных объектов, полевых определителей для смартфонов и планшетов, методических пособий по организации проектной деятельности школьников и полевых экологических исследований (включая книгу для педагогов "Как организовать полевой экологический практикум"), а также учебно-методических фильмов по организации проектной исследовательской деятельности школьников в природе. Приобрести все эти материалы можно в нашем некоммерческом Интернет-магазине. Там же можно приобрести mp3-диски Голоса птиц средней полосы России и Голоса птиц России, ч.1: Европейская часть, Урал, Сибирь.
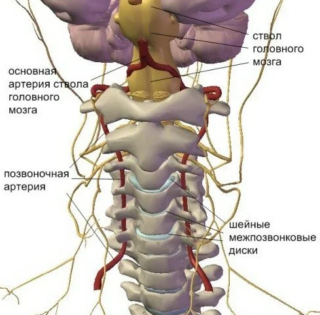
Позвоночный столб — результат сложного процесса эволюции. С развитием прямого хождения эта ось приняла на себя нагрузку, осуществляемую тяжестью тела. Шейные позвонки являются самыми подвижными сегментами позвоночника. Они могут выдержать массу в 20 раз больше, чем железобетонный столб. Краниовертебральное сочленение позвоночного столба, локализующееся в верхнем отделе шеи, включает такие составляющие, как атлант (первый сегмент шеи сверху) и аксис (второй позвонок). Соединение первого и второго шейного позвонка с затылочным основанием черепа обеспечивает функцию вращения головы. В этом же отделе локализуется сосудистая стенка, позвоночная артерия, а также продолговатый мозг. Они отвечают за регуляцию вегетативных функций на рефлекторном уровне.
Особенности развития 1 и 2 шейного позвонка у человека
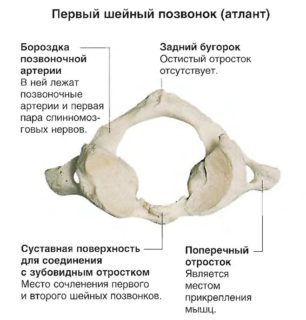
Шейный отдел включает семь позвонков. Сегменты отличаются особой морфологией. Помимо этого, образования обладают собственным обозначением. Сегменты маркируются латинской буквой С. Каждому позвонку присвоен порядковый номер. Таким образом, С1–С7 представляют обозначение цервикального отдела. Сегменты С1 и С2 имеют собственные названия – атлант и аксис.
Отличие атланта заключено в отсутствии тела, так как в период эмбрионального развития происходит срастание сегмента со вторым позвонком, что формирует его зуб. По этой причине диаметр позвоночного отверстия увеличен, а тело представляет короткую переднюю дугу.
Передняя дуга сочленяется с задней с боковых сторон посредством пары утолщений — боковых масс. С внешней стороны передняя дуга обладает бугорком. Внутренняя сторона снабжена ямкой зуба, которая предназначена для соединения атланта с зубом аксиса. На дуге сзади располагается бугорок , представляющий остистый отросток с недоразвитой морфологией. На верхней плоскости задней дуги атланта находиться борозда позвоночной артерии.
С верхней и нижней части латеральной массы первого шейного позвонка располагаются суставные поверхности. Верхние сегменты обладают овальной формой. Они соединены с мыщелками кости затылка, формируя атлантозатылочный сустав. Нижние сегменты обладают круглой формой. Сочленяются с суставными поверхностями второго позвонка шеи, образуя латеральный атлантоосевой сустав.
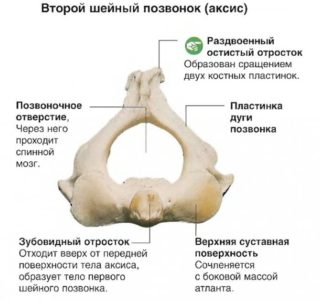
Эпистрофей (аксис) выделяется отличным строением благодаря зубовидному отростку, вокруг которого осуществляется вращение атланта вместе с сочленяющимся с ним черепом. Зуб включает верхушку и пару суставных поверхностей. К внутренней стороне прикреплены связки, придающие ей шероховатую структуру. Поперечные отростки шейных сегментов снабжены отверстиями, формирующими костный канал. Через эту область проходят артерии и нервы позвоночного столба.
С верхней и нижней части дуги выступают суставные отростки, формирующие дугоотросчатые сегменты. Поверхность суставов этих отростков расположена горизонтально. Это обеспечивает подвижность позвонков шеи в горизонтальной плоскости, позволяет достичь большого угла скручивания зоны шеи, что способствует подвижности головы. Подобная анатомия стала важной ступенью в эволюционном процессе. Функция позволяет держать под зрительным контролем большой ареал при низкой подвижности тела.
У положительного признака есть и своя отрицательная сторона: атлант и аксис обладают небольшой массой и высоким уровнем подвижности, что делает их уязвимыми для травм и повреждений.
Строение и соединение позвонков
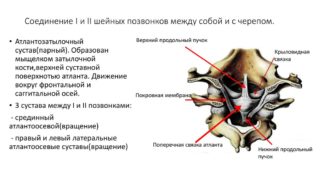
Соединение атланта и осевого позвонка с черепом отличается сложностью. Сочленение мыщелков костей затылка с суставными поверхностями атланта образует комбинированный атлантоаксиально-затылочный комплекс. Он обладает формой эллипса.
Соединение атланта и второго шейного позвонка обеспечивается тремя суставами:
- Четно-комбинированным парным латеральным атланто-осевым. Плоская структура локализуется между нижними поверхностями суставов атланта и верхними боковыми поверхностями второго шейного позвонка.
- Нечетно-цилиндрическим срединным атланто-осевым. Располагается между зубом осевого позвоночного сегмента и суставной ямкой первого позвонка.
Суставы скреплены посредством прочных связок. Между передними и задними дугами первого сегмента и краем большого отверстия в затылке пролегают передняя и задняя атланто-затылочные перепонки, мембрана. Между боковыми массами образования проходит поперечная связка.
От верхнего свободного края поперечной связки отходит тяж на основе волокнистой ткани. Он примыкает к полудуге большого затылочного отверстия, которое располагается спереди. От нижнего края до тела эпистрофея также пролегает волокнистая связка. Мышечные пучки сверху и снизу в совокупности с поперечной связкой образуют крестообразное соединение. От верхней части боковых поверхностей зубообразных отростков отходят две крыловидные связки, примыкающие к мыщелкам кости затылка.
Таким образом, атлант и эпистрофей у человека обладают тремя суставами. Некоторые анатомы выделяют четвертый вид, располагающийся между задней поверхностью сустава зуба и поперечной связкой первого позвонка.
Функции и роль в организме
Латеральные атлантоаксиальные суставы, обладающие парным строением, осуществляют вращение головы по оси относительно позвоночного столба.
Между дугами атлантоосевого сустава и боковыми сегментами, в углублении, пролегает спинномозговая артерия и ганглии спинного мозга, представляющие узлы спинномозговых нервов. Рядом с шейными позвонками расположено множество сухожилий, связок и хрящевых тканей. Эти соединения обладают подвижностью и обеспечивают круговые движения высокой амплитуды, а также наклонные и вращательные движения головы. Во время осуществления поворотов первый шейный позвонок как бы начинает скользить вместе с черепом по гладкой поверхности суставов вокруг оси эпистрофея.
Также на расположение первого шейного позвонка влияет функциональность атлантозатылочного сустава. Он осуществляет соединение мыщелков (отростков пластины снаружи), кости затылка и суставных фасеток (углублений) первого шейного позвонка. В атлантозатылочном суставе осуществляется движение вокруг фронтальной и сагиттальной осей. Первая отвечает за кивание, то есть наклоны головы вперед и назад. Вторая ось осуществляет наклоны в разные стороны.
Передний конец сагиттальной оси расположен несколько выше, чем задний. Благодаря косой топографии оси параллельно с наклоном вбок головой осуществляется небольшой поворот в противоположное направление.
Отростки и впадины на первом позвонке повторяют соответствующие изгибы на поверхности дуги второго сегмента. При смещении выступы атланта не попадают точно в выступы эпистрофея. В результате подвижность головы и шеи становится ограниченной.
Группа связок обеспечивает прочность сочленения шейных позвонков. Вместе с атлантом и аксисом они формируют костно-связочный аппарат. Связки препятствуют смещению атланта вперед. Они выдерживают нагрузку до 130—160 кг. Защитное свойство не обеспечивает сегменту С1 абсолютную неуязвимость.
Читайте также: