
Парасимпатическая нервная система крестцового отдела
Крестцовый отдел парасимпатической нервной системы включает:
Парасимпатические крестцовые ядра (nuclei parasympathici sacrales).
Тазовые внутренностные нервы (nn. splanchnici pelvini) (преганг- лионарные волокна).
Экстрамуральные и интрамуральные тазовые узлы.
Крестцовые парасимпатические ядра заложены в сером веществе 2–4 крестцовых сегментов спинного мозга (1-й нейрон). Аксоны клеток этого ядра (преганглионарные волокна) выходят из спинного мозга вначале в пе- редних корешках крестцовых нервов, далее следуют в составе их передних ветвей. Пройдя вместе с ними через тазовые крестцовые отверстия, пре- ганглионарные волокна отделяются от крестцовых нервов в виде тазовых внутренностных нервов.
Тазовые внутренностные нервы достигают экстрамуральных и ин- трамуральных тазовых узлов (2-й нейрон) вегетативных сплетений орга- нов малого таза и некоторых органов брюшной полости.
Нисходящей ободочной кишки.
Сигмовидной ободочной кишки.
Тазовых отделов мочеточников и мочевого пузыря.
Наружных и внутренних половых органов: у женщин — клитор, матка, маточные трубы, яичники, влагалище, у мужчин — половой член, яички, семявыносящие протоки, простата, семенные пузырьки (рисунок 12). Постганглионарные парасимпатические волокна тазовых внутренно-
стных нервов, достигающие пещеристых тел полового члена (клитора), об- ладают сосудорасширяющим эффектом и тем самым способствуют запол- нению пещеристых тел кровью. Эти реакции лежат в основе эрекции, а са- ми волокна именуются возбуждающими нервами.
Перечень органов, получающих парасимпатическую иннервацию из кре- стцового отдела парасимпатической нервной системы представлен в таблице 2.
Таблица 2 — Перечень органов, получающих парасимпатическую иннер- вацию из крестцового отдела парасимпатической нервной системы
Органы брюшной полости
Органы малого таза
Левая половина поперечной ободочной кишки, нисходящая ободочная кишка, сигмовидная ободочная кишка
Прямая кишка, мочевой пузырь, тазовые отделы мочеточников, мочеиспускательный канал; жен- ские половые органы — матка, маточные трубы, яичники, влагалище, клитор; мужские половые ор-
ганы — яички, семявыносящие протоки, простата, семенные пузырьки, половой член
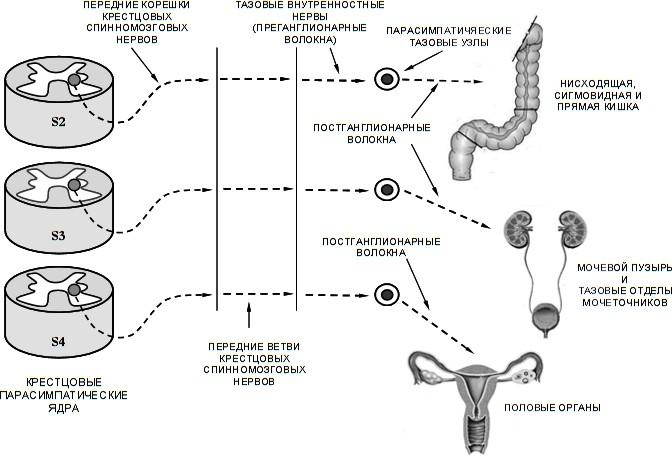
Рисунок 12 — Схема крестцового отдела периферической части парасимпатической нервной системы
Классификация симпатической нервной системы Латеральное Промежуточное ядро боковых столбов (с8 – l2)
Как отмечалось ранее, симпатическая нервная система включает 2 час- ти: центральную и периферическую (рисунок 13).


Рисунок 13 — Схема строения симпатической нервной системы
Центральная часть
Центральная часть симпатической нервной системы целиком локали- зована в спинном мозге и представлена правым и левым латеральными промежуточными ядрами (nuclei intermedia laterales), расположенными в боковых столбах 8-го шейного, всех грудных и верхних двух поясничных сегментов (C8 –L2) (рисунок 14).


Рисунок 14 — Схема центральной части симпатической нервной системы

Периферическая часть симпатической нервной системы объединяет:
Симпатический нервный ствол (truncus sympahticus) с его узлами.
Ветви и нервы симпатического ствола.
Висцеральные сплетения (вокруг сосудов и органов) (рисунок 15).
Рисунок 15 — Схема периферической части симпатической нервной системы
СТРОЕНИЕ ПЕРИФЕРИЧЕСКОЙ ЧАСТИ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
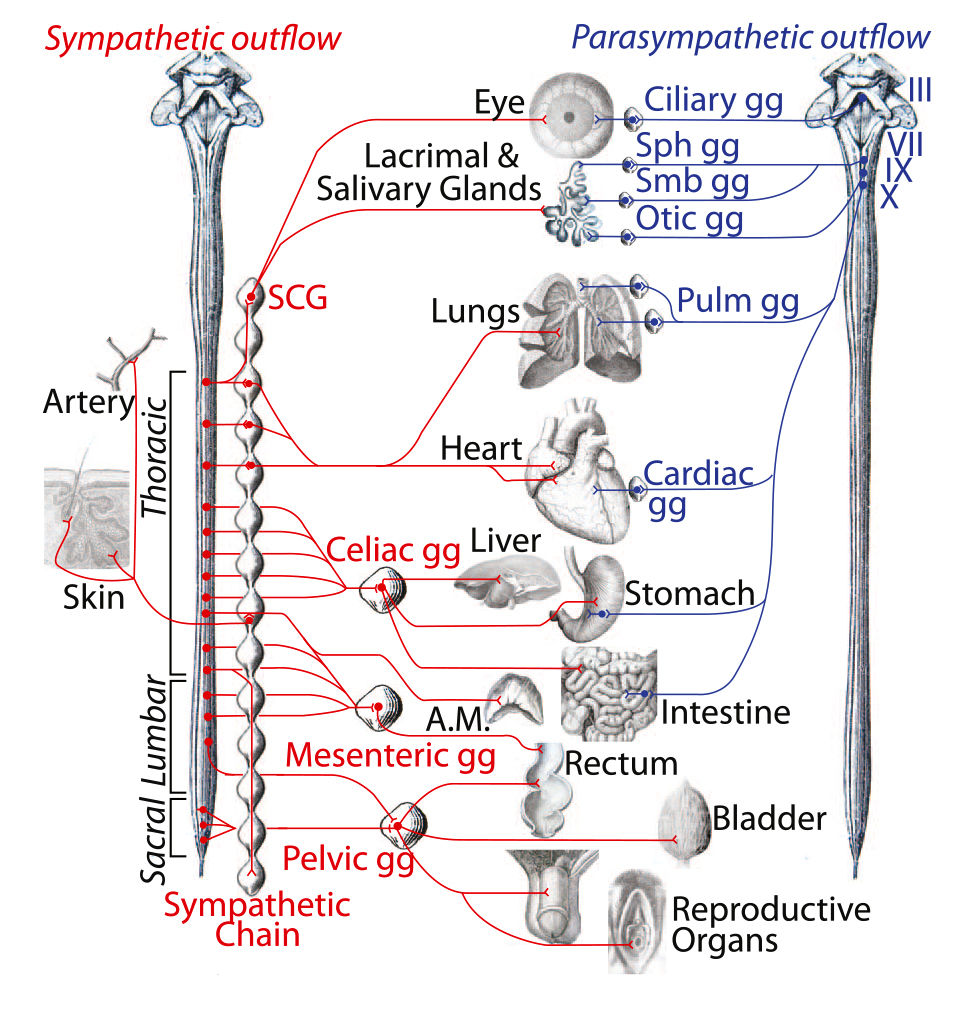
Симпатический нервный ствол — парная цепочка из 20–25 паравертеб- ральных узлов, расположенных на боковых поверхностях позвоночного стол- ба от основания черепа до верхушки копчика. Узлы соединены между собой вертикальными, горизонтальными и диагональными межузловыми ветвями (рисунок 15). Кроме того, каждый узел при помощи серой и белой соедини- тельных ветвей связан с рядом расположенным спинномозговым нервом.
Белая соединительная ветвь (БСВ) несет преганглионарные волокна из латерального промежуточного ядра в составе спинномозгового нерва, за- тем отделяется от него и заканчивается в узле симпатического ствола. БСВ свое название получила из-за белого цвета волокон, покрытых миелином.
Серая соединительная ветвь (ССВ) несет постганглионарные во- локна из узла симпатического ствола к спинномозговому нерву и соеди- няется с ним. Далее волокна ССВ следуют в составе ветвей спинномозго- вого нерва к коже и мышцам (рисунок 16).

Рисунок 16 — Схема формирования белой и серой соединительных ветвей
Симпатический ствол включает 4 топографических отдела:

В течение столетия считалось, что черепные и крестцовые висцеральные нервы позвоночника — родственные образования. Согласно этому, крестцовые преганглионарные нейроны, как и ганглии, расположенные в области малого таза и контролирующие функции прямой кишки, мочевого пузыря, половой системы, считаются парасимпатическими. В этой публикации раскрываются 15 фенотипических и онтогенетических особенностей, которые отличают пре- и постганглионарные нейроны краниального отдела парасимпатической системы от симпатических нейронов грудопоясничного отдела у мышей. Каждая из особенностей доказывает, что крестцовый отдел неотличим от грудопоясничного. Таким образом, парасимпатическая нервная система получает импульсы исключительно от черепных нервов, а симпатическая нервная система от спинномозговых, в том числе от торакальных и крестцовых. Это упрощенное, двухкомпонентное строение, предлагает новую концепцию нейрофизиологии тазового дна, а также эволюции и развития вегетативной нервной системы.
Отнесение крестцового отдела к парасимпатической вегетативной нервной системе — как и связь черепного отдела с крестцовым — имеет древнюю, хотя и простую историю: это убеждение, уходящее корнями в работу Уолтера Гаскелла [1], было документально оформлено Лэнгли [2], и с тех пор стало общепринятым [3]. Аргументом послужили некоторые сходства крестцового отдела с краниальным: (I) анатомическое — область иннервации, в отличие от грудопоясничного отдела, менее рассеяна, отличается от него отсутствием иннервации конечностей, а также недостатком проекций на паравертебральный симпатический ствол [1] (II); физиологическое — влияние на некоторые органы противоположно грудопоясничному отделу [4]; и (III), фармакологическое — повсеместная чувствительность к антагонистам мускариновых рецепторов [2]. Однако, не хватало анализа клеточного фенотипа. В этой статье исследователи определили дифференциальные генетические признаки и взаимосвязи парасимпатических и симпатических нейронов, как пре-, так и постганглионарных. Когда они рассмотрели крестцовый отдел вегетативной нервной системы мышей с такого ракурса, то обнаружили, что его лучше отнести к симпатической, а не к парасимпатической системе.
Черепные парасимпатические преганглионарные нейроны образуются в прогениторной области заднего мозга “pMNv”[5], экспрессирующей гомеоген Phox2band и, ко всему прочему, формирующей бранхиомоторные нейроны [6]. Постмитотические предшественники мигрируют дорсально [7] с образованием ядер (таких как дорсальное двигательное ядро блуждающего нерва) и через дорсолатеральные точки выхода выступают в качестве ветвей некоторых черепных нервов, иннервирующих парасимпатические и энтеральные ганглии. В противоположность этому, симпатические преганглионарные нейроны грудного и верхнего поясничного отдела (далее просто “грудного”), предположительно, имеют общее происхождение с соматическими мотонейронами [8, 9]. Подразумевается, что они образуются в прогениторной области pMN (дорсальнее р3) — то есть из прогениторных клеток, которые экспрессируют основной фактор транскрипции OLIG2 типа спираль-петля-спираль (bHLH) [10]. Симпатические преганглионарные предшественники затем отделяются от соматических мотонейронов, формируя интермедиолатеральный столб у млекопитающих [11], затем вступают в вентральные корешки спинномозговых нервов вместе с аксонами соматических мотонейронов, и с помощью белых соединительных ветвей образуют синапсы на нейронах паравертебральных и превертебральных симпатических ганглиев.
Исследователи стремились сравнить генетический состав и взаимосвязи нижних поясничных и крестцовых преганглионарных нейронов (ниже просто “крестцовых”) с черепными (парасимпатическими) и грудными (симпатическими). В качестве представителя черепных преганглионарных нейронов они избрали дорсальное двигательное ядро блуждающего нерва, скопление нейронов, заметное уже на 13–14 день эмбрионального развития (E13.5) и экспрессирующее везикулярный переносчик ацетилхолина (VAChT) (рис. 1В). Грудные и крестцовые преганглионарные нейроны, которые образуют медиолатеральный столб спинного мозга, несмотря на их возможную холинергическую природу на данном этапе не экспрессируют VAChT. Для того, чтобы определить их локализацию, был использован общий маркер — синтаза оксида азота (NOS) [12] (рис.1, A и B), который отсутствовал в дорсальном двигательном ядре блуждающего нерва на E13.5 (рис.1В) или позже. Таким образом, экспрессия NOS характерна для грудных и крестцовых, но не для черепных преганглионарных нейронов.
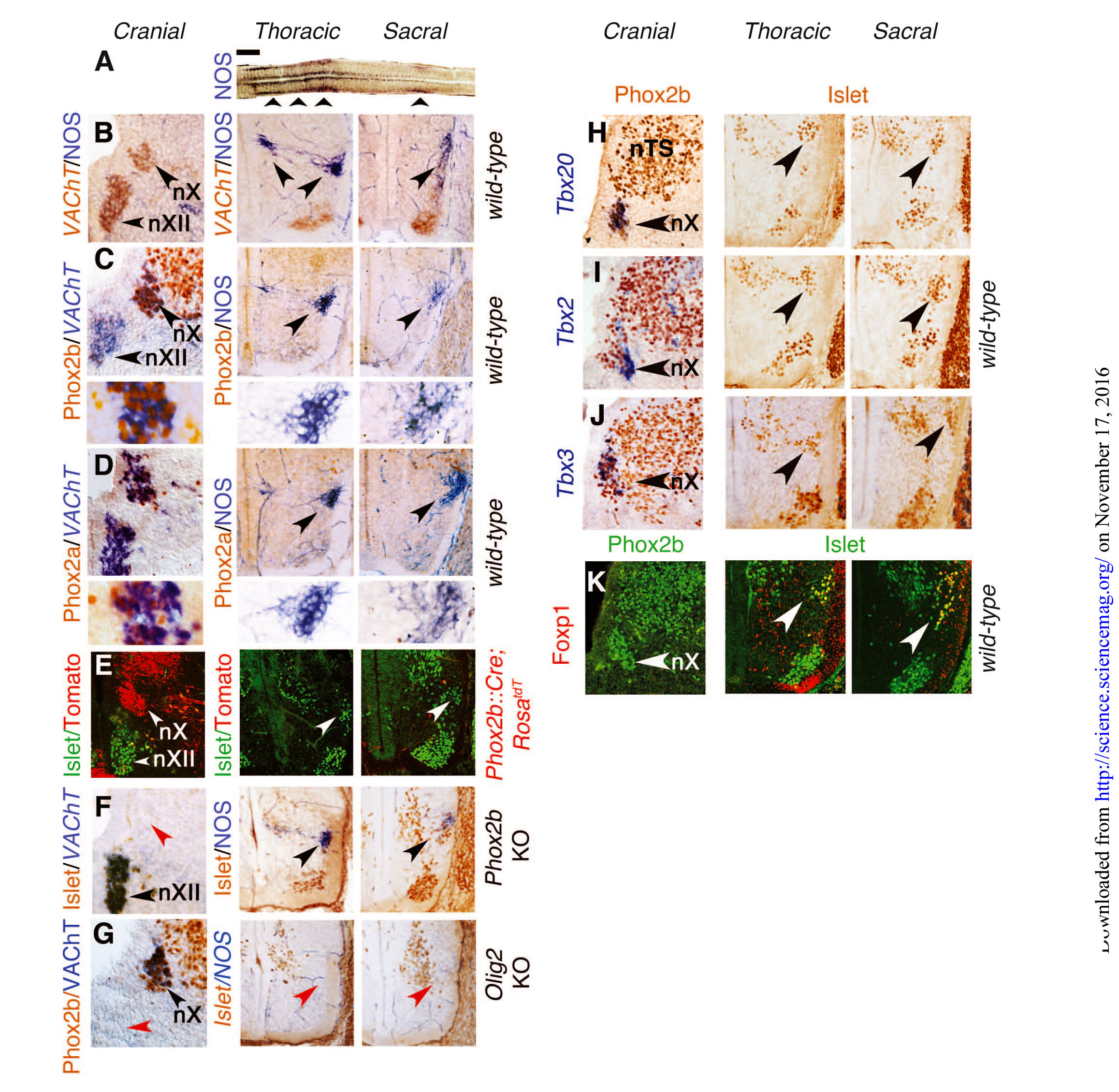
В отличие от черепных (парасимпатических) преганглионарных нейронов, грудные (симпатические) преганглионарные нейроны не только не экспрессируют Phox2b или его паралог Phox2a на E13.5, но также образуются из Phox2b-негативных прогениторов, а вместо Phox2b их дифференцировка (рис. 1, от C до F, левый и средний столбцы) зависит от OLIG2 (рис. 1G). Крестцовые преганглионарные нейроны в этом плане схожи с грудными (рис. 1, от С до G, средний и правый столбцы). На E13.5, экспрессия факторов транскрипции Tbx20, Tbx2 и Tbx3 семейства T-box была выражена в черепных (парасимпатических) нейронах, но отсутствовала как в грудных (симпатических), так и в крестцовых преганглионарных нейронах (рис. 1, от Н до J). Фактор транскрипции Foxp1 семейства F-box — определяющий фактор преганглионарных нейронов грудного отдела [13] — экспрессировали крестцовые, но не черепные преганглионарные нейроны (рис. 1К). Различия в экспрессии Phox2b, Tbx20 и FoxP1 между черепными и всеми спинномозговыми преганглионарными нейронами грудного и крестцового отделов, также наблюдается на E16.5. В целом, онтогенез и транскрипционные характеристики крестцовых преганглионарных нейронов были неотличимы от грудных, а, следовательно, и от симпатических нейронов.
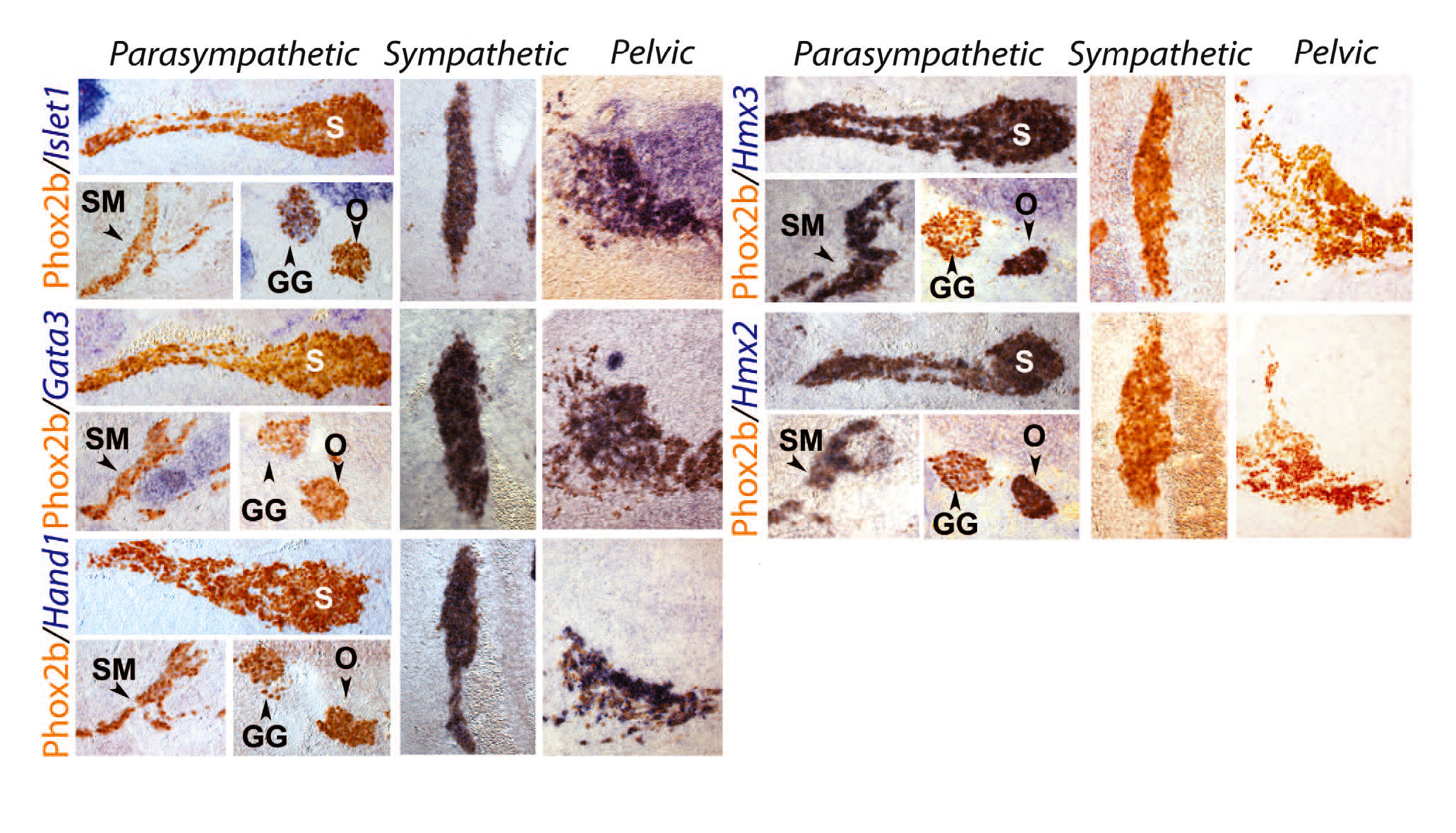
Грудные и крестцовые преганглионарные нейроны располагаются в медиолатеральной области спинного мозга, их аксоны выходят вентрально, в то время как топография черепных преганглионарных нейронов менее систематизирована, а их аксоны выходят дорсально. Эти сходства крестцового отдела с грудным, а также их отличия от краниального противоречат представлению о краниосакральном сходстве с момента его первого описания [1].
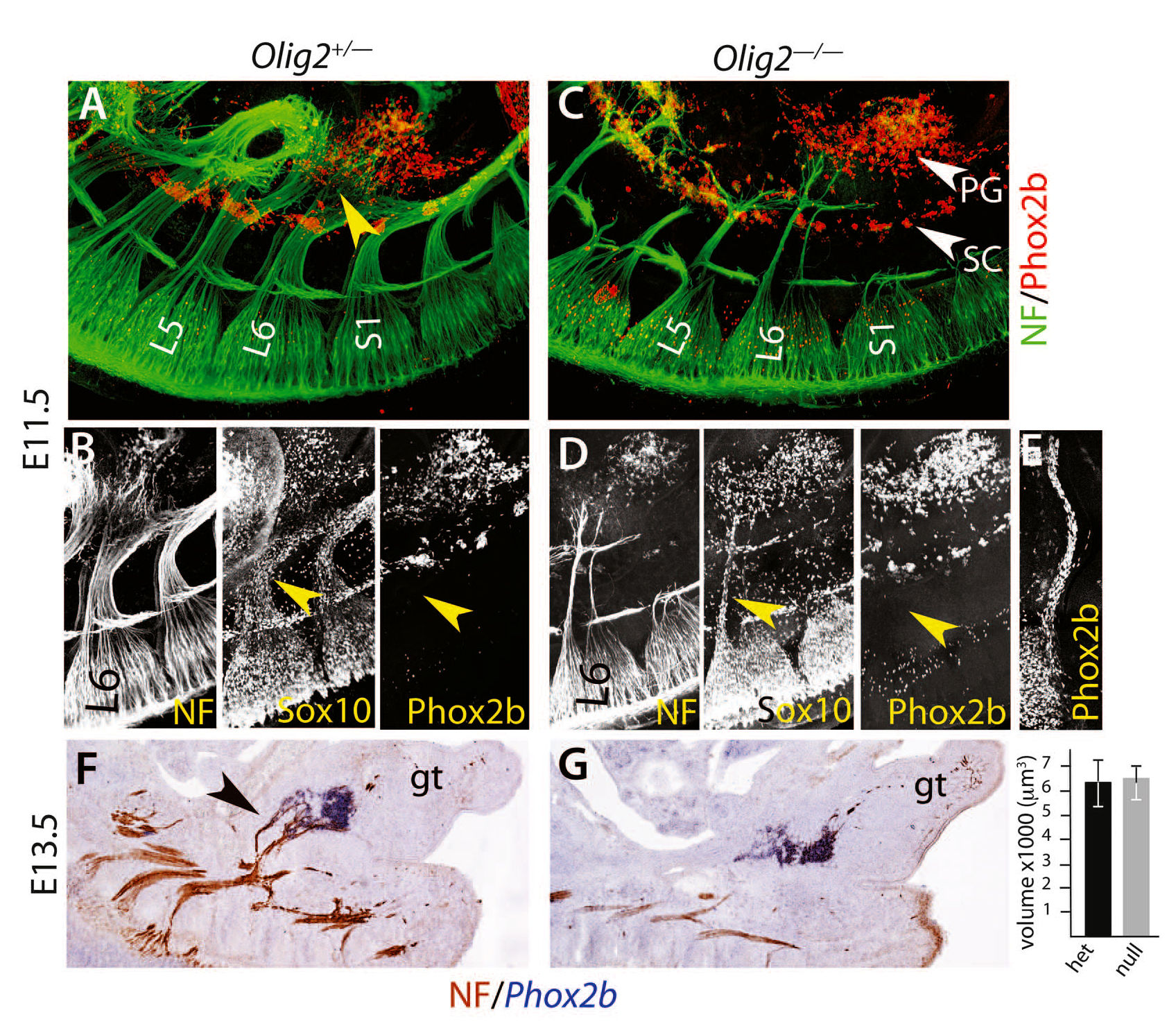
Наконец, опираясь на различия в режимах развития симпатических и парасимпатических ганглиев был проверен тазовый ганглий. Парасимпатические ганглии, в отличии от симпатических, возникают в результате миграции предшественников шванновских клеток Sox10+/Phox2b+ вдоль их будущего преганглионарного нерва по направлению к месту формирования ганглия и не образуют его, если эти нервы отсутствуют [24, 25]. На E11.5 пояснично-крестцовое сплетение, которое дает начало тазовому нерву, распростерло волокна, которые достигли бокового и рострального краев зачатка тазового ганглия и большинство из которых уже расположилось далеко впереди него (рис. 3А и видео S1). Эти волокна были покрыты клетками, экспрессирующими Sox10+, но (рис. 3B), в отличие от черепно-мозговых нервов, которые образуют парасимпатические ганглии на той же стадии, но не экспрессируют Phox2b (рис. 3E). Удаление всех моторных волокон у OLIG2-/- эмбрионов оставило только два тонких, предположительно, сенсорных, выступа от крестцового сплетения (рис. 3C), а также сильно уменьшило количество клеток с Phox2b+ (рис. 3D и рис. S10). Несмотря на эту массовую атрофию, тазовый ганглий оказался интактен (рис. 3C видео S2). Это было количественно проверено на E13.5 (рис. 3, F и G).
Хотя на 50% это клетки постганглионарного тазового нерва, тазовый ганглий формируется раньше и независимо от нерва, как и подобает симпатическому ганглию, а не парасимпатическому. Таким образом, крестцовый отдел вегетативной нервной системы является нижним форпостом симпатического отдела (рис. 4), а сама вегетативная нервная система разделена на краниальную и спинальную части, в соответствии с определенными эволюционными теориями [26]. Это новое понимание анатомии учитывает многие данные, которые противоречили предыдущим. Например, хотя обычно схемы представляют крестцовый путь к прямой кишке как дисинаптический, т.е. вагус-подобный [3], на самом деле он преимущественно [27, 28] трисинаптический, т.е. симпато-подобный [29]. Несмотря на догму об антагонизме между поясничным и крестцовым отделами, проявляющемся действием на мышцу-детрузор мочевого пузыря, в эксперименте торможение поясничного отдела отсутствует [4] или имеет сомнительную функциональную значимость [30]. Синергизм поясничного и крестцового отделов, проявляющийся расширением сосудов во внешних половых органах [29] показывает преемственность действия, а не антагонизм, как предлагает старая модель нервных импульсов грудопоясничного и крестцового отделов.
Симпатическая идентичность всех крестцовых и тазовых вегетативных нейронов, которую раскрывают новые данные, представляет собой основу для открытий в области нейроанатомии и физиологии таза.
- W. H. Gaskell, J. Physiol. 7, 1–80.9 (1886).
- J. N. Langley, The Autonomic Nervous System: Part I (W. Heffer, Cambridge, 1921).
- E. Kandel, J. Schwartz, T. Jessell, S. Siegelbaum, A. J. Hudspeth, Principles of Neural Science, Fifth Edition (McGraw-Hill Professional, 2012).
- J. N. Langley, H. K. Anderson, J. Physiol. 19, 71–139 (1895). 5. J. Briscoe et al., Nature 398, 622–627 (1999).
- A. Pattyn, M. Hirsch, C. Goridis, J. F. Brunet, Development 127, 1349–1358 (2000).
- S. Guthrie, Nat. Rev. Neurosci. 8, 859–871 (2007).
- Prasad, M. Hollyday, J. Comp. Neurol. 307, 237–258 (1991).
- A. Markham, J. E. Vaughn, J. Neurobiol. 22, 811–822 (1991).
- W. A. Alaynick, T. M. Jessell, S. L. Pfaff, Cell 146, 178–178.e1 (2011).
- P. E. Phelps, R. P. Barber, J. E. Vaughn, J. Comp. Neurol. 330, 1–14 (1993).
- C. R. Anderson, Neurosci. Lett. 139, 280–284 (1992).
- J. S. Dasen, A. De Camilli, B. Wang, P. W. Tucker, T. M. Jessell, Cell 134, 304–316 (2008).
- J. R. Keast, Int. Rev. Cytol. 248, 141–208 (2006).
- J. R. Keast, Neuroscience 66, 655–662 (1995).
- Kuntz, R. L. Moseley, J. Comp. Neurol. 64, 63–75 (1936).
- W. C. De Groat, A. M. Booth, J. Krier, in Integrative Functions of the Autonomic Nervous System, C. M. Brooks, K. Koizumi, A. Sato, Eds. (University of Tokyo Press, Tokyo, 1979), pp. 234–245.
- U. Ernsberger, H. Rohrer, Cell Tissue Res. 297, 339–361 (1999).
- K. Huber et al., Dev. Biol. 380, 286–298 (2013).
- K. Tsarovina et al., Development 131, 4775–4786 (2004).
- E. Doxakis, L. Howard, H. Rohrer, A. M. Davies, EMBO Rep. 9, 1041–1047 (2008).
- L. Huber, M. Ferdin, J. Holzmann, J. Stubbusch, H. Rohrer, Dev. Biol. 363, 219–233 (2012).
- C. L. Yntema, W. S. Hammond, J. Exp. Zool. 129, 375–413 (1955).
- V. Dyachuk et al., Science 345, 82–87 (2014).
- Espinosa-Medina et al., Science 345, 87–90 (2014).
- S. Nilsson, in Autonomic Nerve Function in the Vertebrates, Zoophysiology, vol. 13, D. S. Farner, Ed. (Springer-Verlag, New York, 1983), chap. 2.
- C. Olsson et al., J. Comp. Neurol. 496, 787–801 (2006). 28. K. Fukai, H. Fukuda, J. Physiol. 362, 69–78 (1985).
- W. Jänig, The Integrative Action of the Autonomic Nervous System: Neurobiology of Homeostatis (Cambridge Univ. Press, Cambridge, UK, 2006).
- W. C. de Groat, W. R. Saum, J. Physiol. 220, 297–314 (1972).
- A. Pattyn, X. Morin, H. Cremer, C. Goridis, J. F. Brunet, Nature 399, 366–370 (1999).
Поясничный и крестцовый ( тазовый ) отделы симпатического ствола.
Поясничный, или брюшной, отдел симпатического ствола состоит из четырех, иногда из трех узлов. Симпатические стволы в поясничном отделе расположены на более близком расстоянии один от другого, чем в грудной полости, так что узлы лежат на переднебоковой поверхности' поясничных позвонков вдоль медиального края m. psoas major. Rami communicdntes albi имеются только с двумя или тремя верхними . поясничными нервами.
От брюшного отдела симпатического ствола на всем протяжении отходит большое количество ветвей, которые вместе с nn. splanchnici major et minor и брюшными отделами блуждающих нервов образуют самое большое непарное чревное сплетение, plexus coeliacus. В формировании чревного сплетения участвуют также многочисленные спинномозговые узлы (С5 — L3), аксоны их нейроцитов (Д. М. Голуб, 1963). Оно лежит на передней полуокружности брюшной аорты, позади поджелудочной железы, и окружает начальные части чревного ствола (truncus coeliacus) и верхней брыжеечной артерии. Сплетение занимает участок между почечными артериями, надпочечниками и аортальным отверстием диафрагмы и включает- парный чревный узел, ganglion coeliacum, и иногда непарный верхний брыжеечный узел, ganglion mesentericum superius. \
От чревного сплетения отходит ряд меньших парных сплетений к диафрагме, надпочечникам, дочкам, а также plexus testicularis (ovaricus), следующих по ходу одноименных артерий. Имеется также ряд непарных сплетений к отдельным органам по стенкам артерий, название которых они носят. Из последних верхнее брыжеечное сплетение, plexus mesentericus superior, иннервирует поджелудочную железу, тонкую и толстую кишку до половины протяжения поперечной ободочной.
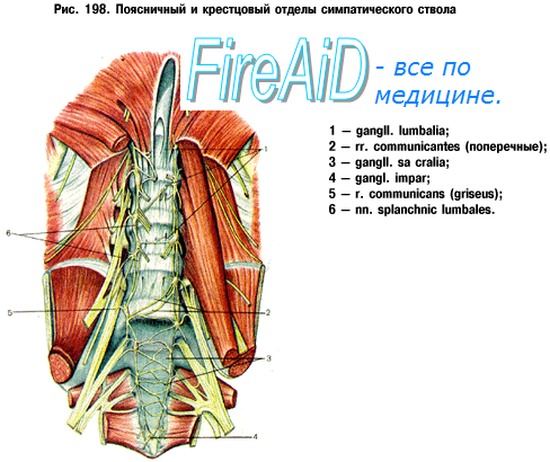
Вторым главным источником иннервации органов полости живота является сплетение на аорте, plexus aorticus abdominalis, составленное из двух стволов, отходящих от чревного сплетения, и веточек от поясничных узлов симпатического ствола. От аортального сплетения отходит нижнее брыжеечное сплетение, plexus mesentericus inferior, для поперечной и нисходящей части ободочной кишки, сигмовидной и верхних отделов rectum (plexus rectals superior). У места отхождения plexus mesentericus inferior располагается одноименный узел, gangl. mesentericum inferius. Его постганглионарные волокна идут в тазе в составе nn. hypogastrics
Аортальное сплетение продолжается вначале в непарное верхнее подчревное сплетение, plexus hypogastricus superior, которое у мыса раздваивается и переходит в сплетение таза, или нижнее подчревное сплетение (plexus hypogastricus inferior s. plexus pelvinus).
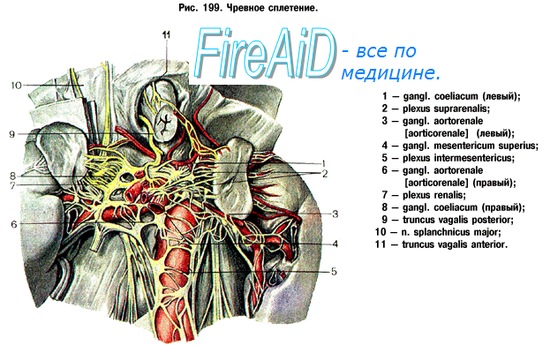
Волокна, происходящие из верхних поясничных сегментов, по своей функции являются сосудодвигательными (вазоконстрикторами) для полового члена, двигательными для матки и сфинктера мочевого пузыря.
Крестцовый, или тазовый, отдел имеет обычно четыре узла; располагаясь на передней поверхности крестца вдоль медиального края передних крестцовых отверстий, оба ствола книзу постепенно сближаются друг с другом и затем оканчиваются в одном общем непарном узле — ganglion impar, находящемся на передней поверхности копчика. Узлы тазового отдела, как и поясничного, связаны между собой не только продольными, но и поперечными стволиками.

От узлов крестцового отдела симпатического ствола отходит ряд ветвей, которые соединяются с ветвями, отделяющимися от нижнего брыжеечного сплетения, и образуют пластинку, протягивающуюся от крестца к мочевому пузырю; это так называемое нижнее подчревное, или тазовое, сплетение, plexus hypogastrics inferior s. plexus pelvinus. Сплетение имеет свои узелки — ganglia pelvina. В сплетении различают несколько отделов: 1) передненижний отдел, в котором выделяют верхнюю часть, иннервирующую мочевой пузырь. — plexus vesicalis, и нижнюю, снабжающую у мужчин предстательную железу (plexus prostaticus), семенные пузырьки и семявыносящий проток (plexus deferentialis) и пещеристые тела (nn. cavernosi penis); 2) задний отдел сплетения снабжает прямую кишку (plexus readies medii et inferiores). У женщин выделяют еще средний отдел, нижняя часть которого дает ветви к матке и влагалищу (plexus uterovaginal), пещеристым телам клитора (nn. cavernosi clitoridis), а верхняя — к матке и яичникам.
От узлов крестцового отдела симпатического ствола отходят соединительные ветви, rami communicantes, присоединяющиеся к спинномозговым нервам, иннервирующим нижнюю конечность. Эти соединительные ветви составляют соматическую часть симпатического отдела вегетативной нервной системы, иннервирующую нижнюю конечность. В составе rami communicantes и спинномозговых нервов нижней конечности находятся постганглионарные волокна, которые распространяются в сосудах, железах и мышцах волос кожи, а также в скелетной мускулатуре, обеспечивая ее трофику и тонус.
Парасимпатическая часть (pars parasympathica) вегетативной (автономной) нервной системы подразделяется на головной и крестцовый отделы (рис. 515). К головному относятся вегетативные ядра и парасимпатические волокна глазодвигательного, языкоглоточного и блуждающего нервов, а также ресничный, крыло-небный, поднижнечелюстной, подъязычный и ушной узлы и их ветви (рис. 516). Крестцовый отдел парасимпатической части образован крестцовыми пара- симпатическими ядрами (nuclei parasympathici sacrales) второго, третьего и четвертого крестцовых сегментов спинного мозга (Sn-SIV), внутренностными тазовыми нервами (nn. splanchnici pelvini), парасимпатическими тазовыми узлами (ganglia pelvica), с их ветвями.
Парасимпатическую часть глазодвигательного нерваобразуют добавочное (парасимпатическое) ядро (nucl. accessorius), ресничный узел и отростки клеток, тела которых лежат в этом ядре и в узле. Аксоны клеток добавочного ядра, залегающего в покрышке среднего мозга, проходят в составе глазодвигательного нерва, заканчиваются на клетках ресничного узла (ganglion ciliare), расположенного у латеральной полуокружности зрительного нерва. Отростки клеток этого узла (постганглионарные волокна) в составе трех-пяти коротких ресничных нервов (nn. ciliares breves) направляются в глазное яблоко и иннервируют ресничную мышцу и сфинктер зрачка.
Парасимпатическая часть лицевого нервасостоит из верхнего слюноотделительного ядра, крыло-небного, поднижнечелюстного и подъязычного узлов и парасимпатических нервных волокон. Аксоны клеток верхнего слюноотделительного ядра, лежащего в покрышке моста, в виде преганглионарных волокон проходят в составе лицевого нерва. В области коленца лицевого нерва часть парасимпатических волокон отделяется в виде большого каменистого нерва (n. petrosus major) и выходит из лицевого канала, идет в крыловидный канал, где вместе с сим- патическим глубоким каменистым нервом образует нерв крыловидного канала (n. canalis pterygoidei). Этот нерв выходит в крыло-небную ямку и направляется к крылонебному узлу (ganglion pterygopalatinum), отростки клеток которого (постганглионарные парасимпатические) присоединяются к верхнечелюстному нерву и следуют в составе его ветвей. Из скулового нерва парасимпатические волокна направляются к слезной железе, которую иннервируют. Носонебный нерв (n. nasopalatinus), большой и малые небные нервы (nn. palatini major et minores), задние лате-
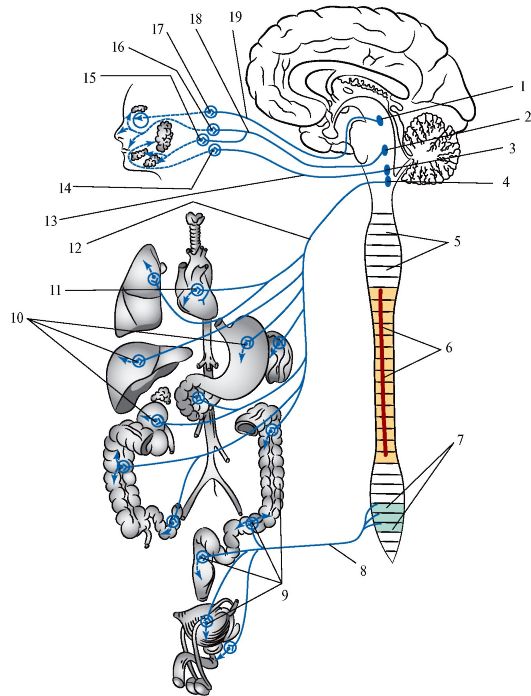
Рис. 515.Схема парасимпатической части вегетативной нервной системы.
1 - добавочное (парасимпатическое) ядро глазодвигательного нерва, 2 - верхнее слюноотделительное ядро, 3 - нижнее слюноотделительное ядро, 4 - дорсальное (заднее) ядро блуждающего нерва, 5 - сегменты спинного мозга, 6 - боковой промежуточный столб спинного мозга, 7 - крестцовые парасимпатические ядра, 8 - тазовые внутренностные нервы, 9, 10 - вегетативные (парасимпатические) узлы, 11 - сердечное сплетение, 12 - парасимпатические волокна блуждающего нерва, 13 - парасимпатические волокна языкоглоточного нерва, 14 - ушной (парасимпатический) узел, 15 - поднижне- челюстной (парасимпатический) узел, 16 - крылонебный (парасимпатический) узел, 17 - ресничный (парасимпатический) узел, 18 - парасимпатические волокна лицевого нерва, 19 - парасимпатические волокна глазодвигательного нерва.
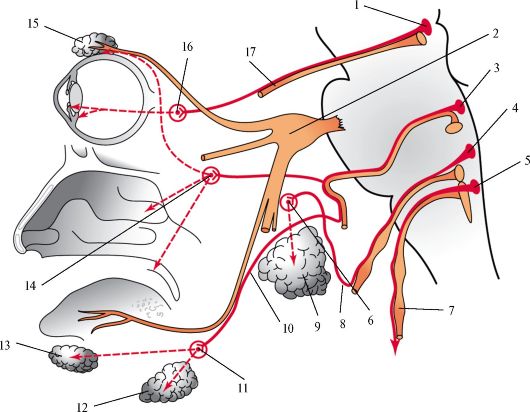
Рис. 516.Парасимпатические вегетативные узлы головы. Схема.
1 - добавочное ядро глазодвигательного нерва, 2 - тройничный узел, 3 - верхнее слюноотделительное ядро, 4 - нижнее слюноотделительное ядро, 5 - дорсальное ядро блуждающего нерва, 6 - ушной узел, 7 - блуждающий нерв, 8 - барабанный нерв, 9 - околоушная железа, 10 - барабанная струна, 11 - поднижнечелюстной узел, 12 - поднижнечелюстная железа, 13 - подъязычная железа, 14 - крылонебный узел, 15 - слезная железа, 16 - ресничный узел, 17 - глазодвигательный нерв.
ральные и медиальные носовые нервы (nn. nasales posteriores laterales et mediales), глоточная ветвь (γ. pharyngeus) иннервируют железы слизистой оболочки полости носа, неба и глотки.
Другая часть преганглионарных парасимпатических волокон, отходит от лицевого нерва в составе барабанной струны (chorda tympani), которая присоединяется к язычному нерву и в его составе направляются к поднижнечелюстному и подъязычному узлам. Поднижнечелюстной узел (ganglion submandibulare) расположен на медиальной стороне поднижнечелюстной слюнной железы, отростки его клеток (постганглионарные нервные волокна) в составе железистых ветвей направляются к поднижнечелюстнои слюнной железе для секреторной ее иннервации. Пoдъязычный узел (ganglion sublinguale) располагается на наружной стороне подъязычной слюнной железы, его железистые ветви идут к одноименной слюнной железе.
Парасимпатическая часть языкоглоточного нерва образована нижним слюноотделительным ядром, ушным узлом и отростками залегающих в них клеток. Аксоны нижнего слюноотделительного ядра, находящегося в продолговатом мозге, идут в составе языкоглоточного
нерва, затем в составе барабанного нерва (n. tympanicus), проникающего в барабанную полость и участвующего в образовании барабанного сплетения. Эти волокна выходят из барабанной полости через расщелину канала малого каменистого нерва в виде малого каменистого нерва (n. petrosus minor), который покидает полость черепа через хрящ рваного отверстия, подходит к ушному узлу, где преганглионарные нервные волокна заканчиваются на клетках ушного узла. Ушной узел (ganglion oticum) прилежит к медиальной стороне нижнечелюстного нерва под овальным отверстием. Постганглионарные волокна клеток этого узла направляются к околоушной слюнной железе в составе околоушных ветвей ушно-височного нерва.
Парасимпатическая часть блуждающего нервасостоит из заднего (парасимпатического) ядра блуждающего нерва, многочисленных узлов, входящих в состав органных вегетативных сплетений, и отростков клеток, расположенных в ядре и в этих узлах. Аксоны клеток заднего ядра блуждающего нерва, находящегося в продолговатом мозге, в виде преганглионарных волокон достигают околоогранных и внутриорганных парасимпатических узлов вегетативных сплетений (сердечного, пищеводного, легочного, желудочного, кишечного и др.). В парасимпатических узлах околоогранных и внутриорганных сплетений располагаются клетки второго нейрона эфферентного пути, отростки которых образуют пучки постганглионарных волокон, иннервирующих гладкую мускулатуру и железы внутренних органов шеи, груди и живота.
Крестцовый отделпарасимпатической части вегетативной нервной системы образован крестцовыми парасимпатическими ядрами (nuclei parasympathici sacrales), расположенными в латеральном промежуточном веществе второго-четвертого крестцовых сегментов спинного мозга, а также тазовыми (парасимпатическими) узлами (ganglia pelvica) и отростками залегающих в них клеток. Аксоны крестцовых парасимпатических ядер выходят из спинного мозга в составе передних корешков, затем идут в составе передних ветвей крестцовых спинномозговых нервов, образующих тазовые внутренностные нервы (nn. splanchnici pelvini). Эти нервы подходят к парасимпатическим узлам нижнего подчревного сплетения и к узлам вегетативных сплетений, расположенных возле внутренних органов, находящихся в полости малого таза. Отростки клеток тазовых паросимпатических узлов (ganglia pelvini) в виде постганглионарных парасимпатических волокон направляются к органам и иннервируют их гладкие мышцы и железы.
Читайте также: