Антигенность иммуноглобулинов. Антигенные детерминанты антител
Добавил пользователь Валентин П. Обновлено: 27.01.2026
Презентация на тему: " Виды антигенных детерминант иммуноглобулинов Изотипические – отражают разнообразие антител на уровне биологического вида (изотипы IgA, IgM, IgG, IgD," — Транскрипт:
3 Виды антигенных детерминант иммуноглобулинов Изотипические – отражают разнообразие антител на уровне биологического вида (изотипы IgA, IgM, IgG, IgD, IgE). Аллотипические – обусловлены генетическим разнообразием внутри вида (аллотипы IgG 1, IgG 2, IgG 3, IgG 4 ). Идиотипические – участки в антигенсвязывающем центре молекулы иммуноглобулина, которые являются антигенными детерминантами. Антитела против таких антигенных детерминант называются антиидиотипическими.
5 Клон - группа генетически идентичных клеток. Фенотип – поверхностная характеристика клетки. CD – кластеры дифференцировки Популяция клеток – типы клеток с наиболее общими свойствами Субпопуляция клеток – более специализированные однородные клетки
6 Т-система включает: Тимус Различные субпопуляции Т-клеток Цитокины, продуцируемые Т- клетками
7 Т-лимфоциты Составляют 60-80% лимфоидных клеток крови Имеют диаметр 6,5 мкм Плотное интенсивно окрашенное ядро занимающее почти всю клетку Эксцентрически расположенную цитоплазму
9 Функции Т-лимфоцитов эффекторная иммунорегуляторная распознавание антигена и запуск иммунного ответа (взаимодействие с макрофагами (АПК). продукция цитокинов
10 Типы Т-лимфоцитов хелперы (CD 4) супрессоры (CD 8) контр-супрессоры эффекторы Т-клетки-памяти
11 Маркеры - поверхностные (или внутриклеточные) структуры, характеризующие как отдельные типы лимфоцитов, так и определенные этапы их развития.
12 Варианты маркеров 1) поверхностные антигены, в т.ч. дифференцировочные, появляющиеся и исчезающие в зависимости от стадии развития клетки или сохраняющиеся на всех стадиях клеточного цикла; 2) поверхностные рецепторы (распознающие структуры), с помощью которых клетки узнают антиген и воспринимают другие стимулы, необходимые для их жизнедеятельности.
13 Рецепторы Т-лимфоцитов – макромолекулярные белковые структуры на поверхности лимфоцитов, с помощью которых распознаются антигены и др. иммунологически значимые молекулы. 1) антиген распознающие рецепторы на Тлф - (TCR), на Влф - рец.Ig природы; позволяют специфически распознавать антиген; 2) рецепторы для иммунологически значимых продуктов иммунной системы (рец. к FcIg, компонентам комплемента, цитокинам и др.) - необходимы для реализации различных функций иммунной системы; 3) рецепторы для продуктов неиммунного происхождения (гормонов, нейропептидов и др.), рецепторы адгезии.
14 Основные рецепторы Т-лимфоцитов TCR К эритроцитам барана (CD 2) К Т-митогенам (ФГА, Кон А) К цитокинам (ИЛ-1, ИЛ-2) HLA I класса
15 Т-клеточный антиген-распознающий рецептор (TCR) представляет комплекс, состоящий из ά, β цепей и молекулы CD3. Функциональная роль CD3 – передача сигнала от ά, β цепей внутрь клетки, запуская процессы активации и пролиферации клетки.
17 Дифференцировка Т-лимфоцитов Антигеннезависимая – костный мозг: ранний преТ-лимф. (экспрессия ГП-33 – β цепь TCR), незрелый преТ-лимф. (экспрессия ГП-33 – α и β цепей TCR), – тимус: развитие толерантности Т-лимфоцитов к аутоантигенам (негативная и позитивная селекция), дифференцировка Т-лимфоцитов в Т-хелперы (CD4) Т-киллеры/супрессоры (CD8) Антигензависимая (Т-зоны периферических лимфоидных органов)
18 Наивные Т-лимфоциты – мигрирующие из тимуса клетки еще не контактировавшие с антигеном и не вступившие в иммунный ответ. Армированные Т-лимфоциты – клетки, активированные антигеном и цитокинами в условиях реального иммунного ответа.
19 Дифференцировка Т-хелперов «Наивный» Тх0 (Тх-индуктор) – комплекс цитокинов, продуцируемых Тх1, Тх2 Тх1 – ИЛ-2, ИФН-γ, ФНО-α Тх2 – ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10, ИЛ-13
20 Дифференцировка Т-хелперных клеток
21 Роль Т-хелперов в иммунном ответе
22 Патология в системе Т-клеточного иммунитета. врожденная недостаточность Т-лимфоцитов (первичный ИД); приобретенные иммунодефициты; инфицирование Т-клеток вирусами (ВИЧ, лейкоза и др.); Т-клеточные иммунопролиферативные заболевания (лимфолейкоз, лимфомы) нарушение соотношения регуляторных Т-лимфоцитов (аутоиммунные и аллергические заболевания).
23 Методы идентификации Т-клеток -Розеткообразование (РОК) -Цитофлюорометрия -Иммуноферментный анализ
Антигенность иммуноглобулинов. Антигенные детерминанты антител
В процессе формирования приобретенного инфекционного иммунитета важная роль принадлежит антителам (анти - против, тело - русское слово, т. е. вещество). И хотя чужеродный антиген блокируется специфическими клетками организма и подвергается фагоцитозу, активное действие на антиген возможно лишь при наличии антител.
Антитела - специфические белки, иммуноглобулины, образующиеся в организме под воздействием антигена и обладающие свойством специфически с ним связываться и отличающиеся от обычных глобулинов наличием активного центра.
Антитела являются важным специфическим фактором защиты организма против возбудителей болезней и генетически чужеродных веществ и клеток.
Антитела образуются в организме в результате инфицирования (естественная иммунизация), или вакцинации убитыми и живыми вакцинами (искусственная иммунизация), или контакта лимфоидной системы с чужеродными клетками, тканями (трансплантанты) либо с собственными поврежденными клетками, ставшими аутоантигенами.
Антитела относятся к определенной фракции белка, главным образом к a -глобулинам, обозначаемым IgY.
- первая - небольшие молекулы с константой седиментации 7S (a-глобулины);
- вторая - большие молекулы с константой седиментации 19 S (a - глобулины).
Молекула антитела включает четыре полипептидные цепи, состоящие из аминокислот. Две из них тяжелые (м.м. 70000 дальтон) и две легкие (м.м. 20000 дальтон). Легкие и тяжелые цепи связаны между собой дисульфидными мостиками. Легкие цепи являются общими для всех классов и подклассов. Тяжелые цепи имеют характерные особенности строения у каждого класса иммуноглобулинов.
В молекуле антитела имеются активные центры, располагающиеся на концах полипептидных цепей и специфически реагирующие с антигеном. Неполные антитела одновалентны (антидетерминанта одна), полные имеют две, реже более антидетерминантны (рис.4).
Рис. 4. Структура иммуноглобулина.
Отличие специфических иммуноглобулинов в строении тяжелых цепей, в пространственном рисунке антидетерминант. Согласно классификации Всемирной организации здравоохранения (ВОЗ), различают пять классов основных иммуноглобулинов: IgG циркулируют в крови, составляют 80% всех антител. Проходят через плаценту. Молекулярная масса 160000. Размер 235 х 40А o . Важны как специфический фактор иммунитета. Обезвреживают антиген путем его корпускуляризации (преципитации, осаждения, агглютинации), что облегчает фагоцитоз, лизис, нейтрализацию. Способствуют возникновению аллергических реакций замедленного типа. По сравнению с другими иммуноглобулинами IgG относительно термоустойчив - выдерживает нагревание при 75 o С 30 мин.
Ig M, - циркулирует в крови, составляя 5-10% всех антител. Молекулярная масса 950000, константа седиментации 19 S, функционально пятивалентен, первым появляется после заражения или вакцинации животного. Ig M не участвует в аллергических реакциях, не проходит через плаценту. Действует на грамположительные бактерии, активизирует фагоцитоз. К классу Ig M относят антитела групп крови человека - А, В, О.
Ig A, - включает два вида: сывороточный и секреторный. Сывороточный Ig A имеет молекулярную массу 170000, константа седиментации 7 S. Не обладает способностью преципитировать растворимые антигены, принимает участие в реакции нейтрализации токсинов, термоустойчив, синтезируется в селезенке, лимфатических узлах и в слизистых оболочках и поступает в секреты - слюну, слезную жидкость, бронхиальный секрет, молозиво.
Секреторный Ig A (S Ig A) характеризуется наличием структурного добавочного компонента, представляет собой полимер, константа седиментации 11 S и 15 S, молекулярная масса 380000, синтезируется в слизистых оболочках. Биологическая функция S Ig A заключается в основном в местной защите слизистых оболочек, например при заболеваниях желудочно-кишечного тракта или дыхательного. Обладают бактерицидностью и опсоническим эффектом.
Ig D, - концентрация в сыворотке крови не более 1%, молекулярная масса 160000, константа седиментации 7 S. Ig D обладает активируемой активностью, не связывается с тканями. Отмечено увеличение его содержания при миеломной болезни человека.
Ig E, - молекулярная масса 190000, константа седиментации 8,5 S. Ig E термолабилен, прочно связывается с клетками тканей, с тканевыми базофилами, принимает участие в реакции гиперчувствительности немедленного типа. Ig E играет защитную роль при гельминтозах и протозойных болезнях, способствует усилению фагоцитарной активности макрофагов и эозинофилов.
Антитела лабильны к температуре 70 0 С, и спирты денатурируют их. Активность антитела нарушается при изменении (отключении) pH среды, электролитов и др.
Все антитела имеют активный центр - площадь участка в 700 А o , что составляет 2% поверхности антитела. Активный центр состоит из 10-20 аминокислот. Чаще всего в них присутствуют тирозин, лизин, триптофан. К положительно заряженным гаптенам антитела имеют отрицательно заряженную группировку - СООН - . К гаптенам, заряженным отрицательно, присоединяется группировка NH4 + .
Антитела обладают способностью отличать один антиген от другого. Они взаимодействуют только с теми антигенами (за редким исключением), против которых они выработаны и подходят к ним по пространственной структуре. Эта способность антитела получила название комплиментарности.
Специфичность антитела обусловлена химической структурой, пространственным рисунком антидетерминант. Она связана с первичной структурой (чередованием аминокислот) белковой молекулы антитела.
Тяжелые и легкие цепи иммуноглобулинов обусловливают специфичность активного центра.
В последнее время обнаружено, что существуют антитела против антител. Они останавливают действие обычных антител. На основе этого открытия возникает новая теория - сетевая регуляция иммунной системы организма.
Теория образования антител затрагивает ряд вопросов из различных смежных дисциплин (генетики, биохимии, морфологии, цитологии, молекулярной биологии), стыкующихся в настоящее время с иммунологией. Существует несколько гипотез синтеза антител. Наибольшее признание получила клонально-селекционная гипотеза Ф. Бернета. Согласно ей, в организме присутствует более 10000 клонов лимфоидных и иммунологически компетентных клеток, способных реагировать с различными антигенами или их детерминантами и вырабатывать антитела. Допускается, что клоны таких клеток способны вступать в реакцию с собственными белками, в результате чего уничтожаются. Так погибают клетки, образующие антиагглютинины против А - антигена у организмов с группой крови А и анти - В - агглютины с группой крови В.
Если эмбриону ввести какой- либо антиген, то аналогичным образом он уничтожает соответствующий клон клеток, и новорожденный в течение всей последующей жизни будет толерантным к данному антигену. Теперь у новорожденного осталось только "свое", либо попавшее извне "чужое", которое распознается мезенхимными клетками, на поверхности которых имеются соответствующие рецепторы "флажки" - антидетерминанты. По мнению Ф. Бернета, мезенхимная клетка, получившая антигенное раздражение, дает начало популяции дочерних клеток, которые вырабатывают специфические (соответствующие антигену) антитела. Специфичность антител зависит от степени их взаимодействия с антигеном.
В формировании комплекса антиген-антитело участвуют возникающие между ионными группами кулоновские силы и силы притяжения Ван-Дер-Ваальса, полярные силы и силы Лондона, межатомные ковалентные связи.
Известно, что взаимодействуют они как целые молекулы. Поэтому на одну молекулу антигена приходится значительное количество молекул антител. Они создают слой толщиной до 30 А o . Комплекс антиген-антитело разъединим с сохранением первоначальных свойств молекул. Первая фаза соединения антитела с антигеном неспецифическая, невидимая, характеризуется абсорбцией антитела на поверхности антигена или гаптена. Протекает при температуре 37 o С за несколько минут. Вторая фаза специфическая, видимая, завершается феноменом агглютинации, преципитации или лизиса. В этой фазе необходимо присутствие электролитов, а в некоторых случаях и комплемента.
Несмотря на обратимость процесса, комплексообразование между антигеном и антителом играет положительную роль в защите организма, которая сводится к опсонизации, нейтрализации, иммобилизации и ускоренной элиминации антигенов.
- коагулирующие (преципитины, агглютинины), облегчают фагоцитоз;
- лизирующие (комплементсвязывающие: бактериолизисы, цитолизисы, гемолизисы), вызывают растворение антигена;
- нейтрализующие (антитоксины), лишают антиген токсичности.
Реакция антиген-антитело может быть для организма полезной, вредной или индифферентной. Положительное влияние реакции в том, что она нейтрализует яды, бактерии, облегчая фагоцитоз, преципитирует белки, лишая их токсичности, лизирует трепонемы, лептоспиры, животные клетки.
Комплекс антиген-антитело может быть причиной лихорадки, расстройства клеточной проницаемости, интоксикации. Может возникнуть гемолиз, анафилактический шок, крапивница, сенная лихорадка, бронхиальная астма, аутоиммунное расстройство, отторжение трансплантата, аллергические реакции.
В иммунной системе нет готовых структур, вырабатывающих антитела и осуществляющих реакции иммунитета. Антитела образуются в ходе иммуногенеза.
Антигенность иммуноглобулинов. Антигенные детерминанты антител
Антигенность иммуноглобулинов. Антигенные детерминанты антител
Антигенные свойства иммуноглобулинов послужили теми фенотипическими признаками, изучение которых позволило установить закономерности генетической регуляции биосинтеза иммуноглобулинов. Любая молекула иммуноглобулина обладает, по-видимому, той или иной антительной специфичностью, т. е. способна взаимодействовать с чужеродными для данного организма веществами — антигенами. Однако и сама молекула иммуноглобулина способна выступать в роли антигена в тех случаях, когда иммуноглобулины одного вида (например, человека) вводятся особям другого вида (например, кролика).
Различают три типа антигенных детерминант молекул иммуноглобулина: изотипы, аллотипы, идиотипы. Изотипическими антигенными детерминантами являются те участки молекул иммуноглобулинов, антигенные свойства которых идентичны у всех особей данного вида.
Каждый класс иммуноглобулинов имеет свои, характерные только для данного класса, изотипические антигены, которые локализованы на постоянной области тяжелых цепей. Изотипические детерминанты, характерные для легких цепей каппа- и ламбда-типа, также локализованы на постоянной области цепи. Разные классы иммуноглобулинов и разные типы легких цепей не имеют общих антигенных детерминант, несмотря на наличие гомологичных последовательностей.
Однако подклассы иммуноглобулинов имеют как общие для разных подклассов антигенные детерминанты, так и детерминанты, специфичные только для данного подкласса.
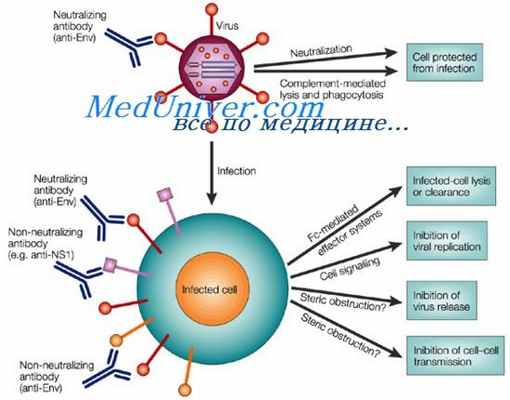
К аллотипическим антигенным детерминантам (аллотипам) относятся те антигенные детерминанты молекул иммуноглобулинов, которые имеются у одних особей данного вида и отсутствуют у других, и эти различия определяются аллельными генами. Наличие аллотипов является отражением внутривидового полиморфизма в антигенном строении молекул иммуноглобулинов.
И, наконец, третий тип антигенных детерминант — это идиотипические детерминанты (идиотипы). К идиотипам относятся те индивидуальные антигенные свойства, которые присущи только молекулам антител данной специфичности или индивидуальным миеломным иммуноглобулинам. Антигенная специфичность идиотипов зависит от строения вариабельной области молекулы антитела, и в ряде случаев имеются определенные доказательства, что идиотипы являются отражением антигенных свойств активного центра молекулы антитела.
Антитела к изотипическим детерминантам используются для идентификации различных классов и подклассов иммуноглобулинов и типов легких цепей. Антитела же к аллотипам служат для обнаружения генетических вариантов иммуноглобулинов, причем аллотипические маркеры локализованы, как правило, на постоянной части полипептидных цепей иммуноглобулинов. Что же касается идиотипических детерминант, то их локализация на вариабельной части молекулы иммуноглобулина позволяет их использовать в качестве генетических маркеров вариабельной части.
История обнаружения генетических маркеров полипептидных цепей иммуноглобулинов вкратце такова. Уже давно было известно, что в сыворотке больных ревматоидным артритом часто содержатся так называемые агглютинаторы, которые способны специфически взаимодействовать с аутологичным IgG. Для обнаружения агглютинаторов используются эритроциты людей Rh+, покрытые неполными aHTH-Rh-антителами, т. е. антителами, которые неспособны агглютинировать эритроциты. Агглютинация наступает только после добавления агглютинатора, способного взаимодействовать с анти-Rh-антителами на поверхности эритроцитов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Антигенность антител
Иммуноглобулин, как и всякий белок, обладает антигеностью и выраженной иммуногенностью. В молекуле Ig различают 4 типа атигенных детерминант: видовые, изотипические, аллотипические и идиотипические. Видовые антигенные детерминанты характерны для Ig всех особей данного вида (например, кролика, собаки, человека). Они определяются строением легкой и тяжелой цепи. Поэтим детерминантам можно идентифицировать видовую принадлежность антител.
Изотипические антигенные детерминанты являются групповыми. Они локализуются в тяжелой цепи и служат для дифференцировки семейства Ig на 5 изотипов (классов) и множество подклассов (см. разд. 11.1.3).
Аллотипические антигенные детерминанты являются индивидуальными, т. е. присущими конкретному организму. Они располагаются в легкой и тяжелой полипептидных цепях. На основании строения аллотипических детерминант можно различать особи внутри одного вида.
Идиотипические антигеннные детерминанты отражают особенности строения ан-тигенсвязывающего центра самой молекулы Ig. Они образованы V-доменами легкой и тяжелой цепи молекулы Ig. Обнаружение идиотипических антигенных детерминант послужило основанием для создания теории «идиотип-антиидиотипической» регуляции биосинтеза антител.
Механизм взаимодействия антитела с антигеном
В процессе взаимодействия с антигеном принимает участие не вся молекула Ig, а лишь ее ограниченный участок — антигенсвязывающий центр, или паратоп, который локализован в Fab-фрагменте молекулы Ig. Co своей стороны, антитело взаимодействует не со всей молекулой антигена сразу, а лишь с ее антигенной детерминантой.
Антитела отличает специфичность взаимодействия, т. е. способность связываться соСвязь антигена с антителом осуществляется за счет слабых взаимодействий (ван-дерваальсовы силы, водородные связи, электростатические взаимодействия) в пределах антигенсвязывающего центра. Такая связь отличается неустойчивостью — образовавшийся иммунный комплекс (ИК) может легко диссоциировать на составляющие его компоненты. Поэтому взаимодействие антигена и антитела может быть представлено в виде уравнения:
[АГ] + [AT] « [ИК].
Продолжительность существования иммунного комплекса определяется целым рядом факторов. При этом важное значение имеют особенности антитела, антигена и условия, в которых происходит их взаимодействие.
К особенностям антитела следует отнести его аффинность и авидность.
Аффинность — сила специфического взаимодействия антитела с антигеном (или энергия их связи). Эта характеристика зависит от степени стерического, или пространственного, соответствия (комплементарности) структуры антигенсвязывающего центра и антигенной детерминанты. Чем выше их комплементарность, т. е. чем больше они подходят друг другу, тем больше образуется межмолекулярных связей и тем выше будет устойчивость и продолжительность жизни образовавшегося иммунного комплекса. Структурные несоответствия антигенсвязывающего центра и антигенной детерминанты существенно снижают число образующихся связей и прочность взаимодействия антитела с антигеном. Иммунный комплекс, образованный низкоаффинными антителами, чрезвычайно неустойчив, имеет малую продолжительность существования и быстро распадается на исходные компоненты.
• Установлено, что в условиях макроорганизма с одной и той же антигенной детерминантой способны одновременно прореагировать и образовать иммунный комплекс около 100 различных клонов антител. Все они будут отличаться структурой антигенсвязывающего центра и аффинностью.
Аффинность антител существенно меняется в процессе иммунного ответа в связи с селекцией наиболее специфичных клонов В-лимфоцитов. Наименее аффинными считаются нормальные антитела. По расчетам, общее число различных антигенспецифических клонов В-лимфоцитов достигает 10 6 —10 7 .
Под термином «авидность» понимают прочность связывания антитела и антигена. Эта характеристика определяется аффинностью Ig и числом антигенсвязывающих центров. При равной степени аффинности наибольшей авид-ностью обладают антитела класса М, так как они имеют 10 антигенсвязывающих центров.
Особенности антигена также влияют на эффективность его взаимодействия с антителом. Так, важное значение имеют стерическая (пространственная) доступность антигенной детерминанты для антигенсвязывающего центра молекулы Ig и число эпитопов в составе молекулы антигена.
Эффективность взаимодействия антитела с антигеном существенно зависит от условий, в которых происходит реакция, и прежде всего от рН среды, осмотической плотности, солевого состава и температуры среды. Оптимальными для реакции антиген—антитело являются физиологические условия внутренней среды макроорганизма: близкая к нейтральной реакция среды, присутствие фосфат-, карбонат-, хлорид- и ацетат-ионов, осмолярность физиологического раствора (концентрация раствора 0,15 М), а также температура (36—37 °С).
научная статья по теме Методы поиска антигенных детерминант для белков с известной первичной структурой Химия

Текст научной статьи на тему «Методы поиска антигенных детерминант для белков с известной первичной структурой»
БИООРГАНИЧЕСКАЯ ХИМИЯ, 2000, том 26, М 4, с. 243-262
МЕТОДЫ ПОИСКА АНТИГЕННЫХ ДЕТЕРМИНАНТ ДЛЯ БЕЛКОВ С ИЗВЕСТНОЙ ПЕРВИЧНОЙ СТРУКТУРОЙ
Московская государственная академия тонкой химической технологии им. М.В. Ломоносова,
117571, Москва, просп. Вернадского, 86; * Российский кардиологический научно-производственный комплекс МЗ РФ, 121552, Москва, 3-я Черепковская ул., 15а Поступила в редакцию 10.04.98 г. Принята к печати 11.12.99 г.
Рассмотрены теоретические и экспериментальные методы определения антигенных детерминант белков с известной аминокислотной последовательностью. Проведена систематизация методов на основе используемых теоретических подходов. Приводятся данные по сравнительной оценке эффективности различных предсказательных методов, а также примеры нахождения эпитопов экспериментальным путем для ряда белков.
Ключевые слова: белки, аминокислотная последовательность, предсказание антигенной структуры; антигенная детерминанта; эпшпопное картирование; пептидные библиотеки.
1. Факторы, влияющие на антигенность белка.
2. Теоретические методы поиска антигенных детерминант белков (предсказание антигенной структуры белка).
2.1. Методы локализации антигенных детерминант, основанные на определении физико-химических свойств полипептидной цепи.
2.2. Статистические методы предсказания вторичной структуры белка.
2.3. Предсказание третичной структуры белка.
2.4. Практическое применение теоретических предсказаний.
2.5. Сравнительная оценка методов предсказания антигенных детерминант.
3. Экспериментальные методы поиска антигенных детерминант.
3.1. Фрагментация комплекса антиген-антитело.
3.2. Генно-инженерные методы.
3.3. Рентгеноструктурный анализ контактных участков антигенного комплекса.
Сокращения: ароС-Н- аполипопротеин С-11; ароЕ - аполи-лопротеин Е; 5норм - температурный фактор; ELISA - метод твердофазного иммуноферментного анализа; ßo-gpl, gp41 и gpC3a - гликопротеины ß2-gp-I, 41 и СЗа; HEV - вирус гепатита Е; HIV - вирус иммунодефицита человека; HTLV - вирус Т-клеточного лейкоза; MAP - мультиплет-ный антигенный пептид; МНС - главный комплекс гисто-совместимости; ORF3 - белок вируса гепатита Е; Р2-С -белок вируса Коксаки; SCR - структурно-консервативный регион.
3.4. Использование синтетических пептидов для локализации антигенных детерминант.
Понимание биологических функций белков -один из центральных вопросов биохимии. Белки реализуют свои функции через связывание с молекулой-мишенью, образуя с ней комплекс. Образование комплекса может быть временным (фермент - субстрат), может быть необратимым (гаптоглобин - гемоглобин) или же комплекс может проявлять устойчивость, среднюю между двумя этими крайними состояниями.
В любом случае, определение структурных параметров участка белка, ответственного за его связывание с мишенью, необходимо для понимания способа действия молекулы в целом. Разработка подхода к синтетической имитации фрагмента, определяющего биологическую роль белка, открывает массу возможностей для воспроизведения активности этого белка и манипулирования ею. Особенно большое значение такой подход имеет для изучения антигенных свойств белков при решении иммунологических проблем. Успешная замена белка-антигена синтетическим пептидным фрагментом позволяет создать систему диагностики заболевания, способствует эффективной защите от инфекций путем создания вакцины.
Белок является антигеном, если он способен реагировать с антителами или Т-лимфоцитами. Способность белка индуцировать иммунный ответ организма определяет его иммуногенность [1].
Иммунный ответ организма, вызываемый действием на него антигена, осуществляется при участии иммунокомпетентных клеток - В- и Т-лим-фоцитов. В-Лимфоциты и антител^ (иммуноглобулины) распознают антиген, находящийся в нативном виде в физиологической жидкой среде (гуморальный иммунитет). Специфические участки аминокислотной последовательности белка-антигена, с которыми реагируют антитела, называются В-эпитопами. Т-Лимфоциты распознают такие антигены, которые уже претерпели про-цессинг в клетке и утратили нативную форму. Образующиеся в результате протеолиза пептиды (Т-эпитопы) перемещаются на поверхность анти-генпрезентирующей клетки, связываются с молекулами главного комплекса гистосовместимости (МНС) и становятся доступными для восприятия Т-лимфоцитами (клеточный иммунитет).
Для успешной имитации белка-антигена необходимо прежде всего понимание антигенных свойств нашивной молекулы, т.е. локализация в последовательности белка участков, взаимодействующих с иммуноглобулинами и В-лимфоцита-ми. Такие участки были названы антигенными детерминантами, или эпитопами, а те фрагменты гипервариабельной области антитела, которые контактируют с эпитопами, получили название паратопов [2].
Исторически эпитопы были разделены на секвенциальные - отвечающие линейному отрезку полипептидной цепи, и конформационные, включающие в себя аминокислотные остатки, кото-
рые находятся в разных участках полипептидной цепи, но собраны в эпитоп в результате ее упаковки цепи в нативном белке [3]. Атасси подразделил эпитопы на непрерывные и прерывающиеся [4]. На рис. 1 показаны эпитопы разного типа в миоглобине по данным работы [5].
Тот факт, что антитела к белку-антигену способны взаимодействовать также с небольшими линейными пептидами, соответствующими фрагментам его последовательности, подтверждает существование непрерывных эпитопов [6].
Выявление конформационных эпитопов требует рентгеноструктурного исследования комплекса антиген-антитело, что является весьма серьезной задачей, поэтому наибольшее количество информации об антигенной структуре белков основано на сведениях о непрерывных эпитопах. В настоящее время для локализации В-эпитопов предложено значительное количество методов, рассмотрению некоторых из них посвящен данный обзор. Эпитопы, распознаваемые Т-лимфо-цитами (Т-эпитопы), изучены хуже, однако проблема их локализации чрезвычайно важна в свете создания противовирусных вакцин. Вопросы поиска Т-эпитопов в обзоре затронуты лишь частично.
1. ФАКТОРЫ, ВЛИЯЮЩИЕ НА АНТИГЕНIIОСТЬ БЕЛКА
Современная иммунология предлагает двойственную модель узнавания антигена антителом [1]. С одной стороны, реакция антигена с антителом происходит благодаря пространственной компле-ментарности их молекул, т.е. топографическому соответствию взаимодействующих участков. Комплементарность предполагает, что антигенная детерминанта должна находиться на поверхности белковой глобулы, иначе говоря, должна быть доступна для антитела.
С другой стороны, при взаимодействии антиген-антитело происходит конформационная перестройка в том участке полипептидной цепи, который охватывает детерминанта, что предполагает его подвижность. Такая укладка полипептидной цепи, которая обеспечивает экспонирование антигенных детерминант, соответствует определенной вторичной и третичной структуре белка.
Сворачивание полипептидной цепи белков, функционирующих в основном в водной среде, происходит таким образом, что гидрофобные остатки плотно контактируют друг с другом и оказываются внутри белковой глобулы, а гидрофильные (полярные) группы попадают на поверхность белка. Иначе говоря, неполярные остатки выдавливают воду из первоначально "рыхлого" клубка полипептидной цепи, что приводит к компактности и стабильности гидрофобного ядра, а экспони-
рование полярных групп на поверхности глобулы облегчает гидратацию белка. Стабильная и биологически активная структура белка поддерживается благодаря кооперативности водородных, ионных, дисульфидных связей, ван-дер-ваальсовых сил и гидрофобных взаимодействий [7].
Рассмотренные выше факторы, влияющие на антигенность белка, положены в основу различных методов локализации эпитопов. Условно методы разделены на теоретические и экспериментальные (полуэмпирические).
2. ТЕОРЕТИЧЕСКИЕ МЕТОДЫ ПОИСКА АНТИГЕННЫХ ДЕТЕРМИНАНТ БЕЛКОВ (ПРЕДСКАЗАНИЕ АНТИГЕННОЙ СТРУКТУРЫ БЕЛКА)
Как уже упоминалось, антигенная детерминанта должна находиться на поверхности белка. Однако выявить поверхностно-локализованные участки возможно только при рентгеноструктурном анализе кристаллов белков, что является особой проблемой. В этой связи широкое распространение получили методы теоретического предсказания антигенных детерминант, использующие только сведения о первичной структуре белков.
Развитие теоретических методов локализации эпитопов стало возможным благодаря прогрессу техники секвенирования ДНК и белков.
В основе теоретических предсказаний лежит установление определенных физико-химических и статистических характеристик участков полипептидной цепи и выяснение корреляции этих характеристик с антигенностью. Такие свойства, как поверхностная доступность, гидрофильность, сегментная подвижность, конформационные статистические параметры, рассчитываются с помощью компьютерных программ для каждого аминокислотного остатка и представляются в виде шкал (профилей), по экстремумам которых выбираются предполагаемые антигенные детерминанты.
Любое теоретическое предсказание начинается с обращения к банкам данных, содержащим сведения об известных белках, таким, как "Atlas of Protein Sequence and Structure" Дэйхофа [8], "Brookhaven Protein Data Bank" [9], "SWISS-PROT" [10] и др. Объем банков данных неуклонно расширяется; в настоящее время число известных аминокислотных последовательностей превышает 100000 [10], однако количество белков с расшифрованной трехмерной структурой составляет всего 15000 [I I]. Ниже описаны различные подходы к теоретическому предсказанию ант
Для дальнейшего прочтения статьи необходимо приобрести полный текст. Статьи высылаются в формате PDF на указанную при оплате почту. Время доставки составляет менее 10 минут. Стоимость одной статьи — 150 рублей.
Читайте также: