Белки промежуточных филаментов
Добавил пользователь Дмитрий К. Обновлено: 29.01.2026
11.5.1. Промежуточные филаменты образуются из фибриллярных полипептидов четырех типов [51, 52]
В отличие от мономеров актина и тубулина, которые представляют собой глобулярные белки, субъединицы ПФ имеют вытянутую, фибриллярную форму. Они объединяются в продольные пучки, где перекрываются по длине, так что образуют длинные нити с высокой механической прочностью. В латеральных взаимодействиях, за счет которых строятся ПФ, нередко участвует лишь часть молекулы белковой субъединицы ПФ, поэтому структура остальной ее части может значительно варьировать, не изменяя общего строения нити. В связи с этим ПФ в отличие от актиновых филаментов и микротрубочек построены из полипептидов с весьма различной молекулярной массой - от 40 до 130 тыс. в зависимости от типа клеток.
Промежуточные филаменты по их первичной структуре делят на четыре большие группы (табл. 11-5). Белки ПФ типа I наиболее характерны для эпителиальных клеток и включают два подсемейства кератинов: кислые кератины и нейтральные или основные кератины. Кератиновые филаменты - всегда гетерополимеры, образованные поровну субъединицами каждого из этих двух подсемейств. Вообще кератины - самая обширная группа белков ПФ; известно уже не менее 19 различных форм в составе эпителиев человека и еще 8 в волосах и ногтях. Многие типы эпителиев, различающихся морфологически и функционально, синтезируют также разные формы кератинов.
Таблица 11-5. Главные типы белков промежуточных филаментов
Образующий полипептид (мол. масса)
Кислые кератины (40000-70000)
Нейтральные и основные кератины (40 000-70 000)
Эпителиальные клетки и производные эпидермиса (волосы, ногти и т.п.)
Многие клетки мезенхимного происхождения; часто экспрессируется клетками в культуре Мышечные клетки
Глиальный фибриллярный кислый белок (45000)
Глиальные клетки (астроциты и некоторые шванновские клетки)
Белки нейрофиламентов (около 130000 1) , 100000 1) и 60000)
Ядерные ламины А, В и С (65000 -75000)
Ядерная ламина во всех клетках
1) Из-за того, что эти белки при электрофорезе в гелях с додецилсульфатом натрия мигрируют аномально медленно, раньше их молекулярную массу считали большей.
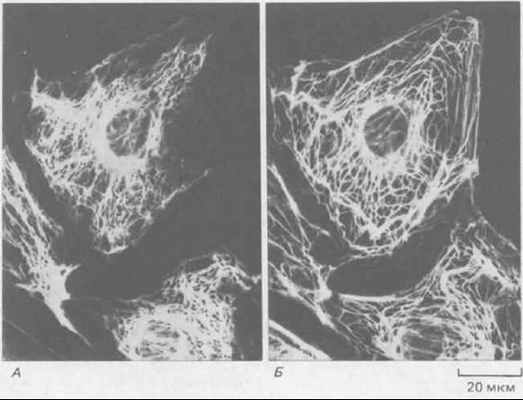
Рис. 11-73. Иммунофлуоресцентная микрофотография эпителиальных клеток кенгуровой крысы (PtK2) в интерфазе. Клетки окрашены одновременно антителами к виментину (А) и к кератину (Б). Обратите внимание, что содержащиеся в клетке системы виментиновых и кератиновых филаментов существуют раздельно, хотя и имеют сходное распределение. (С любезного разрешения Магу Osborn.)
К белкам ПФ типа II относятся 1) виментин, 2) десмин и 3) глиальный фибриллярный кислый белок. Виментин широко распространен в клетках мезенхимного происхождения, включая фибробласты, клетки эндотелия кровеносных сосудов и лейкоциты; он часто образуется в культивируемых клетках и временно появляется в различных клетках в ходе онтогенеза. Десмин содержится в клетках мышц, как гладких, так и поперечнополосатых, а глиальный фибриллярный кислый белок образует глиальные филаменты в определенного рода клетках глии (астроцитах и некоторых шванновских клетках) в нервной системе. Все эти белки способны in vitro к самосборке с образованием гомополимеров, а также к образованию гетерополимеров с другими белками типа II. Последняя способность проявляется и in vivo: в клетках некоторых типов были обнаружены сополимеры виментина с десмином и виментина с глиальным фибриллярным кислым белком.
Из белков ПФ типа III построены нейрофиламенты - важный компонент цитоскелета в аксонах и дендритах нервных клеток. У позвоночных три таких белка, их называют «нейрофиламентным триплетом». И наконец, белки ПФ типа lV-это ядерные ламины (разд. 11.5.5); они сходны с другими белками ПФ по аминокислотной последовательности, но имеют несколько характерных отличий. Наиболее примечательно то, что они образуют высокоупорядоченные двумерные сети из филаментов, подвергающиеся быстрой разборке и сборке на определенных стадиях митоза.
Все клетки эукариот синтезируют ядерные ламины и по крайней мере один тип цитоплазматических белков ПФ. В некоторых клетках есть цитоплазматические ПФ двух типов, образующие раздельные структуры. Таковы, например, некоторые эпителиальные клетки, содержащие отдельные системы кератиновых и виментиновых филаментов (рис, 11-73). 11-28
11.5.2. Промежуточные филаменты образуются из димерных субъединиц со стержневидным срединным доменом [53]
Несмотря на значительную разницу в размерах, все белки ПФ цитоплазмы кодируются генами одного мультигенного семейства. У всех этих белков в первичной структуре полипептида есть гомологичный срединный участок примерно из 310 аминокислот, образующий протяженную а-спираль с тремя короткими не-а-спиральными вставками (рис. 11-74). Кроме того, большие отрезки этой срединной области имеют последовательность, характерную для полипептидов, способных к образованию спирали из двух спиралей (см. разд. 11.1.6). Подобно тропомиозину или хвосту мышечного миозина, эта двухцепочечная спираль представляет собой димер из двух одинаковых полипептидов ПФ. Эти две цепи в гомодимере ПФ уложены параллельно друг другу, причем к срединному стержневидному домену примыкают на обоих концах глобулярные домены. При сборке ПФ стержневидные домены взаимодействуют друг с другом и формируют однородную сердцевину филамента, а глобулярные, величина которых сильно варьирует у разных белков ПФ, выступают с поверхности филамента наружу. Одна из моделей сборки ПФ из димерных субъединиц показана на рис. 11-75.
11.5.3. Промежуточные филаменты простираются от ядерной оболочки до периферии клетки [54]
Если окрасить культивируемые клетки антителами к одному из цитоплазматических белков ПФ (например, виментину), то обычно будет видна ажурная сеть нитей, окружающая ядро и охватывающая всю, цитоплазму (см. рис. 11-73). По структуре эта сеть отлична от других компонентов цитоскелета, хотя местами ее нити, по-видимому, идут параллельно микротрубочкам цитоплазмы. Вероятно, организация цитоплазматических ПФ зависит от взаимодействия с микротрубочками, так как деполимеризация микротрубочек при обработке веществами типа колхицина ведет к «осаждению» всей сети ПФ в виде околоядерной «шапки». Можно думать, что многие ПФ цитоплазмы связаны с ядерной оболочкой и в норме оттягиваются от нее к периферии клетки микротрубочками, с которыми они соединены.
Организация ПФ в цитоплазме может также определяться их взаимодействием с плазматической мембраной. В эритроцитах птиц (которые в отличие от эритроцитов млекопитающих имеют ядро и ПФ) виментин, как полагают, связан с плазматической мембраной через анкирин (разд. 6.2,4). В эпителиальных клетках кератиновые ПФ присоединены к плазматической мембране в десмосомах - специализированных межклеточных соединениях, помогающих удерживать соседние клетки вместе (разд. 14.1.4). Так как кератиновые филаменты каждой клетки через десмосомы соединены с такими же филаментами соседних клеток, они образуют непрерывную сеть, охватывающую весь эпителий.
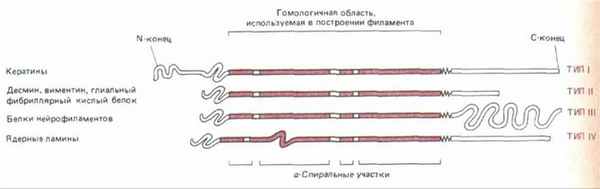
Рис. 11-74. У всех белков промежуточных филаментов имеется гомологичная центральная область (около 310 аминокислотных остатков), формирующая протяженную а-спираль с тремя короткими участками иной структуры. N-концевой и С-концевой домены не состоят из а-спирали и сильно варьируют по размерам и последовательности аминокислот у белков разных промежуточных филаментов.
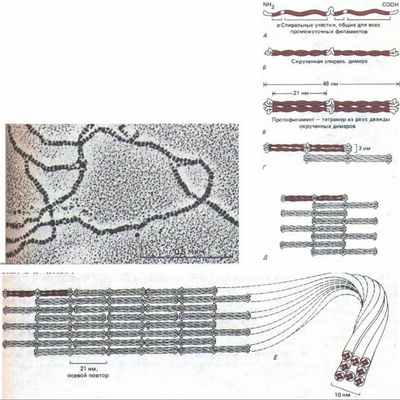
Рис. 11-75. Одна из современных моделей сборки промежуточных филаментов (ПФ). Мономер (А) объединяется с таким же мономером, образуя димер (Б), в котором консервативные а-спиральные участки лежат параллельно, обвиваясь друг около друга. Затем два таких димера укладываются бок о бок, образуя протофиламент длиной 48 нм и толщиной 3 нм, который состоит из четырех полипептидных цепей (В). Такие протофиламенты затем образуют все более крупные структуры, укладываясь с продольным сдвигом (Г и Д). Окончательная структура промежуточного филамента толщиной 10 нм состоит из восьми рядов протофиламентов (32 полипептидных цепей), соединенных в длинный тяж, похожий на канат (Е). Вверху представлена электронная микрофотография такого «окончательного» филамента. Неизвестно, являются ли ПФ полярными структурами, как актин и тубулин, или неполярными, как двойная спираль ДНК (или, что то же самое, лежат ли две скрученные спирали в составе протофиламента в параллельной ориентации или же в антипараллельной. (Микрофотография любезно предоставлена N. Geisler и К. Weber.)
11.5.4. Сборка промежуточных филаментов может контролироваться с помощью фосфорилирования [55]
Изолированные промежуточные филаменты (ПФ) в ионной среде, соответствующей цитоплазме, чрезвычайно стабильны; более того, сколько-нибудь значительного пула неполимеризованных белков ПФ (какой имеется в случае актина и тубулина) в клетке нет. И все же клетка явно может регулировать число, длину и расположение своих промежуточных филаментов, что указывает на ее способность контролировать их сборку и разборку. Важный фактор этого контроля-фосфорилирование определенных остатков в белках ПФ. Виментин, например, существует как в нефосфорилированной, так и в фосфорилированной форме. Если фосфорилировать изолированные виментиновые нити с помощью протеинкиназы, они распадаются на меньшие фрагменты. Однако самый впечатляющий пример того, насколько важную роль играет в контроле разборки ПФ фосфорилирование, - это ядерные ламины, которые подвергаются деполимеризации всякий раз, когда клетка вступает в митоз,
11.5.5. Ядерная ламина образована особым классом промежуточных филаментов [56]
Ядерная ламина - это белковая сеть (обычно толщиной от 10 до 20 нм), подстилающая изнутри поверхность внутренней ядерной мембраны (см. рис. 9-1). Она представляет собой прямоугольную решетку из промежуточных филаментов (рис. 11-76, Д), построенных у млекопитающих из трех белков ПФ типа Vl-ламинов А, В и С (см. рис. 11-74 и табл. 11-3), Ламины образуют димеры, у которых имеется стержневидный домен и две глобулярные головки на одном из концов (рис. 11-76, Б). При подходящих рН и ионной силе димеры самопроизвольно ассоциируют, образуя филаменты, которые по толщине и повторяющейся структуре сходны с цитоплазматическими ПФ.
Однако по ряду признаков ядерные ламины отличаются от белков ПФ цитоплазмы. Наиболее очевидное отличие-это организация образуемых филаментов в прямоугольную решетку (рис. 11-76, А), хотя для такой организации, видимо, необходимо объединение их с другими белками. Кроме того, ядерная ламина - структура очень динамичная. Когда клетки млекопитающих вступают в митоз, кратковременное фосфорилирование нескольких остатков серина в ламинах вызывает обратимую диссоциацию ядерной ламины на тетрамеры гиперфосфорилированных ламинов А и С и связанного с мембраной ламина В. При возвращении клетки в интерфазу ламины дефосфорилируются, и вокруг разошедшихся хромосом вновь образуется цельная ядерная оболочка (разд. 13.5.11).
11.5.6. Кератиновые филаменты удивительно разнообразны [52]
Из всех типов промежуточных филаментов наиболее стабильные и долгоживущие - кератиновые, они же и самые разнообразные. Эпителии с примитивной организацией, например в развивающемся эмбрионе, а также некоторые зрелые ткани (такие, как печень) содержат кератины двух типов - один кислый и один нейтральный. В эпителиях других органов (например, языка, мочевого пузыря, потовых желез) имеются шесть или больше различных кератинов, причем их конкретный спектр зависит от анатомической локализации. Кератиновые филаменты ввиду их многообразия и стабильности могут служить своего рода «отпечатками пальцев», позволяющими уточнить происхождение некоторых опухолей эпителиальной природы. Еще разнообразнее кератины в эпидермисе, который представляет собой плотный многослойный эпителий (разд. 17.4.2). В клетках разных слоев эпидермиса экспрессируются разные наборы кератинов. Кератиновые филаменты в них постепенно сшиваются поперечными ковалентными связями друг с другом и с ассоциированными белками, и по мере гибели клеток в самых наружных слоях эпидермиса поперечносшитый кератиновый скелет становится важнейшим защитным барьером на поверхности тела. Специализированные эпителиальные клетки, образующие такие поверхностные структуры, как волосы, когти и перья, обеспечивают дополнительные локальные вариации в наборе кератинов. Таким образом, промежуточные филаменты защищают животное от потери тепла и воды, предоставляют ему «оружие» и средства камуфляжа или, наоборот, привлечения партнера (окраска).
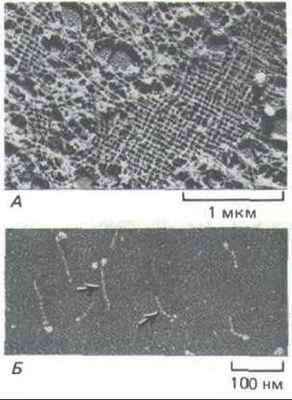
Рис. 11-76. А. Электронная микрофотография участка ядерной ламины в ооците Xenopus (препарат получен методом лиофилизации и напыления металлом). Ламину образует высокоупорядоченная прямоугольная сеть из промежуточных филаментов, состоящих из ядерных ламинов. Б. Изолированные димеры ламина (L) (электронная микрофотография, напыление металлом). По форме они напоминают мышечный миозин (М): у них есть стержневидный хвост и две глобулярные головки, но они гораздо меньше. Глобулярные головки образованы двумя большими С-концевыми доменами. (С любезного разрешения Ueli Aebi.)
11.5.7. Какова функция промежуточных филаментов?
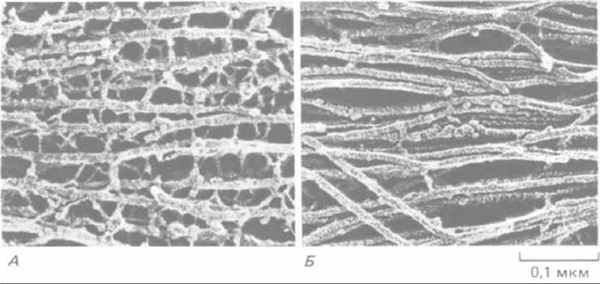
Рис. 11-77. Электронная микрофотография промежуточных филаментов двух типов, встречающихся в нервной ткани (препарат после быстрого замораживания и глубокого травления). А. Нейрофиламенты в аксоне соединены многочисленными поперечными белковыми сшивками; как полагают, такая организация придает этому длинному клеточному отростку большую прочность на разрыв. По-видимому, сшивки образованы длинными неспиральными участками С-концевой части наиболее крупного белка нейрофиламентов (см. рис. 11-74). Б. Промежуточные филаменты (называемые глиальными филаментами) в астроците. Они подвергаются меньшим механическим нагрузкам. Их поверхность довольно гладкая, и поперечных сшивок между ними мало. (С любезного разрешения N. Hirokawa.)
Различные потенции к связыванию других белков могут обеспечиваться вариабельными участками белков промежуточных филаментов, Влияя на свойства филамента, эти вариабельные участки определяют не только его способность к самосборке, но и то, как он будет взаимодействовать с другими компонентами клетки (например, с микротрубочками и плазматической мембраной). Это совершенно иная стратегии чем в случае двух других важнейших элементов цитоскелета - актиновных филаментов и микротрубочек; как мы уже знаем, эти полимеры в основном инвариантны по структуре, а к выполнению различных функций они приспосабливаются с помощью разных наборов актин-связывающих белков и белков, ассоциированных с микротрубочками. Таким образом, роль вариабельных участков в белках промежуточных филаментов та же, что и у вспомогательных белков актиновых филаментов и микро-трубочек, - разница лишь в том, что одни ковалентно связаны с субъединицами филамента, а другие представляют собой отдельные молекулы.
Промежуточные филаменты (ПФ) - это полимеры, по структуре подобные канатам, собранным из нитевидных полипептидов. По- видимому, они поддерживают структуру клеток или противостоят растягивающим нагрузкам. Существует много тканеспецифических форм ПФ, построенных из различных полипептидов: кератиновые филаменты эпителиальных клеток, нейрофиламенты нейронов, глиальные филаменты астроцитов и шванновских клеток, десминовые филаменты мышечных волокон и виментиновые филаменты фибробластов и клеток многих других типов. Отдельное семейство белков ПФ составляют ядерные ламины, из которых построена волокнистая пленка (ламина), выстилающая изнутри оболочку ядра; они имеются во всех эукариотических клетках.
Полипептиды, входящие в состав промежуточных филаментов различных типов, различаются по аминокислотной последовательности, а также - и очень сильно - по молекулярной массе. Однако у всех имеется гомологичный центральный домен, который при димеризации белка образует жесткую структуру из обвивающих друг друга спиралей. Такие димерные субъединицы складываются в большие пучки «внахлест», формируя промежуточные филаменты. Стержневидные домены субъединиц при этом создают структурную сердцевину ПФ, а глобулярные домены на обоих концах выступают наружу и обусловливают разнообразие свойств ПФ. Именно благодаря этой вариабельности механические свойства ПФ и взаимодействия их с другими клеточными компонентами приспособлены к специфическим нуждам клеток того или иного типа.
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Белки промежуточных филаментов
О проекте
Предмет клеточной биологии
Строение и химия клеточного ядра
Цитоплазма
Системы энергообеспечения клетки
Цитоскелет
Механизмы клеточного деления
Промежуточные филаменты (ПФ) строятся из фибриллярных мономеров. Поэтому основная конструкция промежуточных филаментов напоминает канат, имеющий толщину около 8—10 нм. Они локализуются главным образом в околоядерной зоне и в пучках фибрилл, отходящих к периферии клеток и располагающихся под плазматической мембраной (рис. 238, 240 и 241). Встречаются промежуточные филаменты во всех типах клеток животных, но особенно они обильны в тех клетках, которые подвержены механическим воздействиям: клетки эпидермиса, нервные отростки, гладкие и исчерченные мышечные клетки. В клетках растений ПФ не обнаружены.
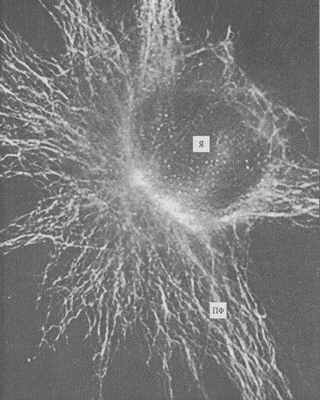
Рис. 240. Промежуточные филаменты (ПФ) фибробласта в культуре ткани, окрашенные флуоресцирующими антителами к виментину (фото Л.А. Горгидзе)
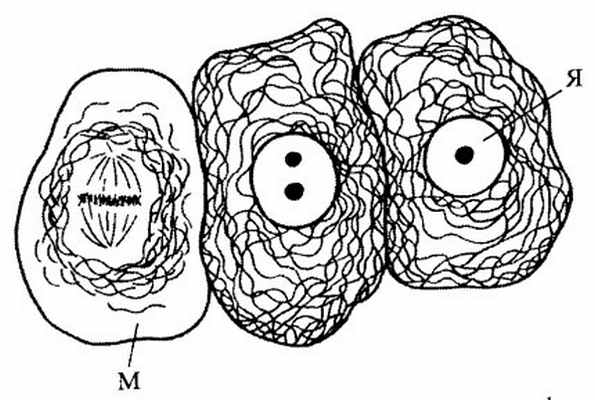
Рис. 241 . Расположение промежуточных филаментов в клетках эпителия
М — митоз; Я — ядро
В состав промежуточных филаментов входит большая группа изобелков (родственных белков), которую можно разделить на четыре типа. Первый тип составляют кератины, кислые и нейтральные, встречающиеся в эпителиальных клетках; они образуют гетерополимеры из этих двух подтипов. Кератины, кроме того, имеют, некоторую гетерогенность, зависящую от тканевого источника. Так, в эпителиях встречается до 20 форм кератинов, 10 форм других кератинов найдено в волосах и ногтях. Молекулярная масса кератинов колеблется от 40 до 70 тыс.
Второй тип белков ПФ включает в себя три вида белков, имеющих сходную молекулярную массу (45—53 тыс.). Это — виментин, характерный для клеток мезенхимного происхождения, входящий в состав цитоскелета клеток соединительной ткани, эндотелия, клеток крови. Десмин характерен для мышечных клеток, как гладких, так и исчерченных. Глиальный фибриллярный белок входит в состав ПФ некоторых клеток нервной глии — в астроциты и некоторые шванновские клетки. Периферинвходит в состав периферических и центральных нейронов.
Третий тип — белки нейрофиламентов (молекулярная масса от 60 до 130 тыс.), встречается в аксонах нервных клеток.
И наконец, четвертый тип — белки ядерной ламины. Хотя эти последние имеют ядерную локализацию, они сходны по строению и свойствам со всеми белками промежуточных филаментов.
Как уже говорилось, промежуточные филаменты построены из фибриллярных белков наподобие каната. При этом некоторые белки могут образовывать сополимеры, например виментин с десмином или виментин с глиальными белками.
Все белки промежуточных филаментов обладают сходной аминокислотной последовательностью из 130 остатков в центральной части фибриллярной молекулы, которая обладает α-спиральным строением. Концевые же участки молекул имеют разные последовательности аминокислот, разную длину и не имеют α-спирального строения. Наличие протяженных α-спиральных участков позволяет двум молекулам образовывать двойную спираль, подобно тому как это происходит в молекуле миозина, что приводит к образованию палочковидного димера длиной около 48 нм. Два димера, объединяясь бок о бок, образуют короткий протофиламент — тетрамер, толщиной около 3 нм. Такие протофиламенты могут объединяться в более толстые и длинные фибриллы, и в конечном итоге образуется промежуточный полный филамент, состоящий из восьми продольных протофиламентов (рис. 242).
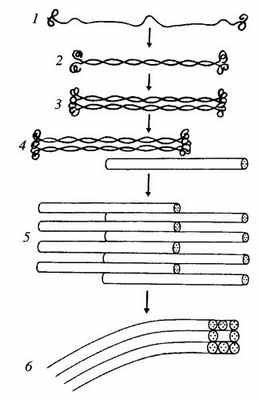
Рис. 242. Этапы полимеризации белков (1—5) и промежуточных филаментов
1 — отдельная молекула; 2 — димер; 3 — тетрамер-протофиламент; 4, 5 — полимеризация протофиламентов; 6 — сформированный промежуточный филамент
Иначе полимеризуются белки ядерной ламины: они образуют димеры с головками на одном конце и полимеризуются, создавая рыхлую прямоугольную решетку. Такие слои ламины быстро разрушаются во время митоза при фосфорилировании ламинов.
Цитоплазматические промежуточные филаменты относятся к самым стабильным и долгоживушим элементам цитоскелета. Однако in vivo наблюдается включение инъецированных меченых молекул кератина в состав ПФ эпителиальных клеток. ПФ устойчивы к действию солей низкой и высокой концентрации, разрушаются только после воздействия денатурирующих растворов, таких как мочевина.
Такая структура и химическая устойчивость промежуточных филаментов, вероятно, определяют и их физическую устойчивость. Они служат как бы истинно опорной системой в клетках, подвергающихся значительным физическим нагрузкам. В клетках кожного эпидермиса промежуточные филаменты образуют пучки (тонофиламенты), связанные с десмосомами, и создают жесткую внутриклеточную сеть (рис. 243). Так, в нервных аксонах, простирающихся на многие десятки сантиметров, ПФ, или нейрофиламенты, создают жесткую основу, обеспечивающую гибкость и целостность тонких цитоплазматических отростков нервных клеток. В поперечно исчерченных мышечных клетках десминовые филаменты входят в состав Z-дисков и связывают их друг с другом как в составе саркомера, так и в соседних миофибриллах, а также с плазматической мембраной.
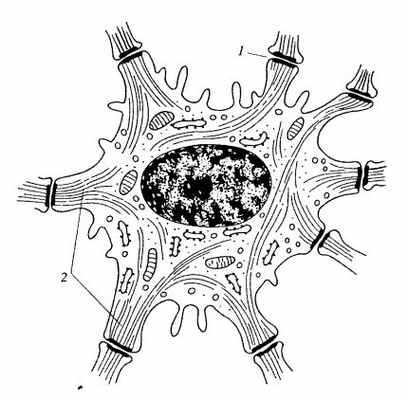
Рис. 243. Клетка шиповатого слоя эпидермиса кожи
1 — десмосомы; 2 — кератиновые промежуточные филаменты
Специфических ингибиторов полимеризации белков промежуточных филаментов пока еще не найдено. Поэтому остается неясным сам процесс сборки и разборки этих элементов цитоскелета в живой клетке. Вероятнее всего, что они подобно ламинам деполимеризуются при действии цитоплазматических киназ, приводящих к их фосфорилированию. Выделенные промежуточные филаменты под действием фосфорилаз могут распадаться на мономеры, деполимеризоваться.
Топографически в клетке расположение промежуточных филаментов повторяет расположение микротрубочек, они как бы идут бок о бок. При разрушении микротрубочек колхицином происходит так называемый коллапс промежуточных филаментов: они собираются в плотные пучки или кольца вокруг ядра. Восстановление новой сети промежуточных филаментов начинается от зоны клеточного центра. Это наводит на мысль, что центром их полимеризации или нуклеации могут быть центры, общие с микротрубочками.
Промежуточные филаменты.
Эта сложная система цитоскелетных нитей изучена относительно недавно. Белки промежуточных филамен- тов обладают сходным строением и построены по общему принципу. Они образованы из двух или трех разных белков, образующих скрученную спираль. Молекулы имеют удлиненную форму и содержат длинный а-спиральный центральный участок, ограниченный линейными головными и хвостовыми участками. Все они собираются в филаменты, диаметр которых около 10 нм. Молекулярная масса большинства белков промежуточных филаментов находится между 40 и 70 кД. Они не подвергаются постоянной сборке и разборке, а представляют собой сравнительно устойчивые цитоскелетные структуры. Важной особенностью промежуточных филаментов является их способность к самосборке без дополнительных источников энергии в виде энергии гидролиза ГТФ или АТФ.
Оказалось, что, в отличие от микротрубочек и актиновых филаментов, промежуточные филаменты построены в разных клетках из разных белков (рис. 3.18).
Выделяют шесть основных типов промежуточных филаментов (табл. 3.3). Они выполняют в клетках те же функции, которые характерны для других компонентов цитоскелета. Но их открытие и изучение имело чрезвычайно важные последствия для молекулярной биологии и медицины. Как видно из табл. 3.3, в разных клетках промежуточные филаменты построены из разных белков, а тубулины и актины у всех одинаковые. Более того, белки из ткани определенного типа у разных организмов более сходны, чем разные филаменты в тканях одного организма. При изменении формы, свойств и даже местоположения клеток в организме белки промежуточных филаментов сохраняются, определяя принадлежность клетки к определенному типу. Это постоянство очень важно для диагностики опухолей разного вида. Дело в том, что при перерождении клетки в раковую, она теряет многие черты своей изначальной организации и определить тип опухоли очень трудно. Но белки промежуточных филаментов остаются такими же, какими были в изначальной ткани. Исследуя белки филаментов в опухолевых клетках, можно точно определить, клетки какой ткани дали начало этой опухоли. Это правило распространяется и на метастазы опухолей, которые могут распространяться далеко от места первоначального образования опухолей. 1
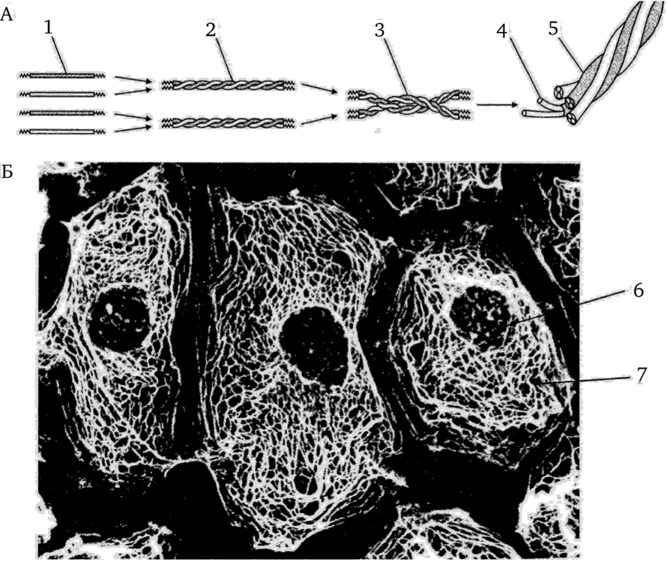
Рис. 3.18. Цитоскелет клетки. Промежуточные филаменты (строение, сборка, расположение в клетке):
А — схема сборки филементов; Б — микрофотография расположения сети промежуточных филатентов в клетке (иммуногистохимия, световой микроскоп): 1 — мономеры; 2 — димеры; 3 — тетрамеры; 4 — протофибриллы; 5 — зрелый промежуточный филамент; 6 — ядро эпителиальной клетки; 7 — сеть кератиновых филаментов в цитоплазме клетки [1]
Состав и принадлежность к определенным типам клеток и тканей промежуточных филаментов цитоскелета животных клеток
Класс промежуточных филаментов
Кератины (более 20 белков)
Клети покровного и кишечного эпителия. Другие виды эпителиев
В мышечных клетках (кроме гладких миоцитов)
В клетках соединительной ткани, хряща, кости, железах, эндотелии сосудов и гладких миоцитах
Глиальный кислый фибриллярный белок
В клетках глии нервной системы
В нервных клетках
Ламины (несколько белков)
Во всех клетках. Расположены в ядре клеток, образуя так называемую ядерную пластинку.
Цитоскелет прокариотической клетки. Долгое время считалось, что в бактериальной клетке нет элементов цитоскелета, однако в начале 1990-х гг. было установлено, что эубактерии и археи содержат элементы цитоскелета, гомологичные по составу и функции цитоске- летным белкам эукариот.
В 2001 г. у бактерии Bacillus subtilis были обнаружены белки — гомологи актина: МгеВ (англ, murein cluster В) и РагМ (англ, partitioning motor), которые формируют в клетке длинные фибриллы. Одной из основных функций белка МгеВ является поддержание палочкоподобной или спиральной формы бактериальной клетки. Однако, в отличие от цитоскелетных белков в эукариотической клетке, они не служат непосредственно каркасом для поддержания формы клетки, а являются местами для прикрепления ферментов, синтезирующих компоненты клеточной стенки бактерии, которая и определяет форму бактериальной клетки. Филаменты, образованные МгеВ, постоянно собираются и разбираются. Показано, что белки МгеВ участвуют в делении бактериальной клетки, концентрируясь в районе перетяжки.
Фибриллы из белка РагМ также постоянно собираются и разбираются в цитоплазме бактерий и состоят из двух протофиламентов, закрученных друг вокруг друга наподобие спирали F-актина. В клетках мономеры РагМ формируют длинные неразветвленные филаменты, которые служат для прикрепления плазмид и их распределения по полюсам клетки при делении.
У большинства эубатерий и архей обнаружен и гомолог тубулина — белок FtsZ, состоящий из двух субъединиц и формирующий филаменты, которые напоминают протофиламенты микротрубочек и обладают ГТФ-азной активностью. В бактериях эти филаменты образуют так называемое Z-колыдо в зоне перетяжки бактерии при ее делении. Еще один гомолог тубулина — белок BtubA/B обнаружен у бактерий из рода Prosthecobacter.
В составе цитоскелета бактерий было обнаружено еще несколько белков: семейство филаментозных белков Min (MinC, MinD, MinE), MamK, бактофилин, кренактин и ряд других. Все они в той или иной степени принимают участие в поддержании формы бактериальной клетки и в процессе деления.
Таким образом, цитоплазма и цитоскелет — это не застывшая, пассивная субстанция, а активная часть клетки, все время изменяющая свою структуру и приспосабливающаяся к постоянно меняющимся условиям внутренней и внешней среды.
Белки промежуточных филаментов
• Промежуточные филаменты представляют собой основные компоненты ядерного и цитоплазматического цитоскелета
• Промежуточные филаменты необходимы для поддержания правильной структуры тканей и их функционирования
• По диаметру промежуточные филаменты находятся между актиновыми филаментами и микротрубочками и образуют прочные сети
• Промежуточные филаменты представляют собой полимеры, состоящие из белковых субъединиц
• Белки, из которых состоят промежуточные филаменты, гетерогенны и кодируются семейством больших и сложно устроенных генов
• У человека более 50 болезней обусловлены возникновением мутаций в белках промежуточных филаментов
Микротрубочки, актиновые филаменты (микрофиламенты) и промежуточные филаменты представляют собой три основные системы белковых филаментов, составляющих цитоскелет. Промежуточные филаменты образуют в цитоплазме и ядре сеть и присутствуют во всех клетках метазоа (животных).
В отличие от микротрубочек и актиновых филаментов, которые необходимы даже для выживания изолированных клеток in vitro, основная функция промежуточных филаментов проявляется на уровне тканевой организации, где они необходимы для надлежащего функционирования тканей и органов. Некоторые типы промежуточных филаментов участвуют в скреплении клеток друг с другом, что необходимо для формирования тканей.
Белки промежуточных филаментов кодируются несколькими большими семействами генов. Эти белки образуют сложную систему филаментов, на долю которых в клетке в нормальных физиологических условиях приходится до 80% общего клеточного белка. Внутриклеточное распределение промежуточных филаментов отличается от характерного для актиновых филаментов и микротрубочек.
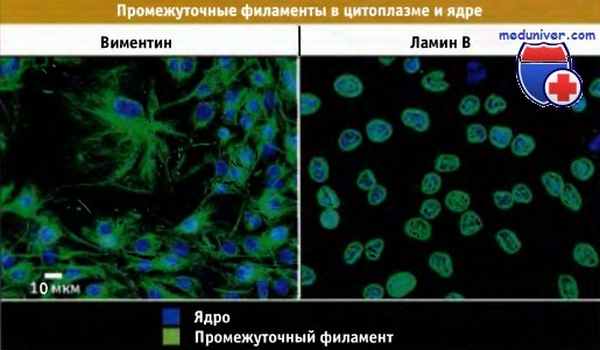
Распределение различных типов промежуточных филаментов в культивируемых фибробластах.
Иммунофлуоресцентное окрашивание на виментин и ламин В. Виментин находится в цитоплазме, а ламины в ядре.
Гистологи обнаружили их (в виде нейрофибрилл нейронов и тонофиламентов клеток эпидермиса) задолго до того, как в 1960-х гг. при электронно-микроскопическом исследовании мышечной ткани были описаны индивидуальные филаменты. В клетках мышц «промежуточные» филаменты занимали по диаметру среднее положение между «толстыми филаментами» миозина-II и «тонкими филаментами» актина. Их средний диаметр составляет около 10 нм, т. е. они толще, чем актиновые филаменты (около 8 нм), и тоньше микротрубочек (около 25 нм). Все три системы филаментов представлены рисунке ниже.
Белки промежуточных филаментов характеризуются общей молекулярной структурой и полимеризуются в филаменты, обладающие высокой механической прочностью. В электронном микроскопе они выглядят одинаково. У высших позвоночных семейство соответствующих белков организовано наиболее сложно, и этот вопрос будет рассмотрен в настоящей главе.
Похожие промежуточные филаменты также обнаружены у беспозвоночных, однако у них количество генов, кодирующих соответствующие белки, значительно меньше, чем у позвоночных. Также промежуточные филаменты беспозвоночных менее гетерогенны и обладают меньшей тканевой специфичностью, чем у млекопитающих. В геноме человека находится порядка 70 генов, кодирующих белки промежуточных филаментов. Принимая во внимание альтернативный сплайсинг для пары из них, общее количество этих белков приближается к 75.
Они представлены гораздо большим числом вариантов и более гетерогенны, чем актиновые или тубулиновые белки. Для всех белков промежуточных филаментов характерна тканеспецифическая экспрессия. Также их экспрессия изменяется в процессе дифференцировки.
Большинство сведений, касающихся экспрессии и биохимических свойств промежуточных филаментов, были получены до того, как были установлены их функции и связь с некоторыми заболеваниями. Сейчас показано, что мутации в генах белков промежуточных филаментов связаны с многими генетическими заболеваниями, которые характеризуются различными фенотипическими проявлениями. Они включают по меньшей мере 50 отдельных болезней, от фликтены до прогерии.
Почти все типы генов белков промежуточных филаментов связаны с той или иной формой проявления хрупкости тканей. Это позволяет предполагать, что для функционирования ткани in vivo необходима надлежащая механическая прочность и что в значительной степени она прямо или опосредованно связана с промежуточными филаментами. Принимая во внимание, что экспрессия генов белков промежуточных филаментов носит тканеспецифический характер, весьма возможно, что все эти белки придают клеткам тканей мельчайшие оттенки различия. Клеткам тканей необходимы различные свойства, такие как прочность, пластичность, быстрота сборки и разборки структур, обеспечивающих прочность.
Может быть, в этом кроется причина того, что в ходе эволюции возникло столь много генов, кодирующих белки промежуточных филаментов.

Основные компоненты цитоскелета в электронном микроскопе.
На ультратонком срезе клетки эпителия почки видны актиновые микрофиламенты, промежуточные филаменты К8/К18 и микротрубочки.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Промежуточные филаменты
Промежуточные филаменты (ПФ) — нитевидные структуры из особых белков, один из трех основных компонентов цитоскелета клеток эукариот. Содержатся как в цитоплазме, так и в ядре большинства эукариотических клеток. Средний диаметр ПФ — около 10 нм (9-11 нм), меньше, чем у микротрубочек (около 25 нм) и больше, чем у актиновых микрофиламентов (5-9 нм). Название получили из-за того, что толщина цитоскелетных структур, состоящих из ПФ, занимала промежуточное положение между толщиной миозиновых филаментов и микротрубочек [1] . В ядре известен только один тип ПФ — ламиновых, остальные типы — цитоплазматические.
Содержание
Структура
Доменная структура белковых молекул ПФ довольно консервативна. Полипептид обычно имеет два глобулярных домена на N- и C-концах, которые соединены протяженным суперскрученным палочковидным доменом, состоящим из альфа-спиралей. Основной строительный блок филамента — димер, а не мономер. Он образован двумя полипептидными цепями, обычно двух разных белков, которые взаимодействуют между собой своими палочковидными доменами, образующими двойную суперскрученную спираль. Цитоплазматические ПФ образованы из таких димеров, образующих неполярные нити, толщиной в один блок. Отсутствие полярности у ПФ обусловлено антипараллельной ориентацией димеров в тетрамере. Из них далее образуются более сложные структуры, в которых ПФ могут уплотняться, вследствие чего имеют непостоянный диаметр.
В отличие от актина и тубулина белки ПФ не имеют сайта сязывания нуклеозидтрифосфатов.
Распространение
Цитоплазматические ПФ есть не у всех эукариот, они обнаружены только у некоторых групп животных. Так, ПФ есть у нематод. моллюсков и позвоночных. но не найдены у членистоногих и иглокожих. У позвочноных ПФ отсутствуют в некоторых клетках (например, олигодендроцитах). В растительных клетках ПФ не обнаружены. [2] [3]
В большинстве животных клеток ПФ образуют «корзинку» вокруг ядра, откуда направлены к периферии клеток. ПФ особенно много в клетках, подверженных механическим нагрузкам: в эпителиях, где ПФ участвуют в соединении клеток друг с другом через десмосомы, в нервных волокнах, в клетках гладкой и поперечно-полосатой мышечной ткани.
В отличие от других основных элементов цитоскелета, ПФ в цитоплазме клеток разных тканей состоят из разных, хотя и похожих по своей структуре белков. Всего белки ПФ у человека кодируют около 70 генов. На основе особенностей аминокислотного состава и строения выделяют пять основных групп белков ПФ.
Тип I — кератины
Из кератинов с молекулярной массой 40 - 70 кДа состоит наиболее разнообразная группа ПФ. Данный тип белков делится на 2 подсемейства:
- кислые кератины,
- нейтральные и основные кератины.
Димер кератина состоит из одного кислого и одного основного кератина. Среди многочисленных изоформ кератина выделяют две основные группы — эпителиальные кератины (см. цитокератин), включающую около 20 видов кератинов, и кератины волос (примерно 10 видов), из которых построены также ногти, рога и чешуя пресмыкающихся.
Тип II
Второй тип белков ПФ включает в себя 4 вида белков:
- - белок с массой 45 - 53 кДа, характерный для клеток мезенхимного происхождения: входит в состав клеток соединительной ткани, эндотелия, клеток крови; ;
- глиальный фибриллярный кислый белок;
- периферин.
Тип III
- Альфа-интернексин
- Белки нейрофиламентов
- Нестин
- Синемин
- Синкойлин
Тип IV
Функции
ПФ в ряде случаев обеспечивают механическую прочность клеток, их отростков или эпителиальных слоев. Они участвуют в образовании межклеточных контактов — десмосом и гемидесмосом.
Медицинское значение
Мутации генов кератинов krt5 и krt14 связаны с развитием наследственного кожного заболевания, буллёзного эпидермолиза (Epidermolysis bullosa simplex). При этом заболевании нарушено прикрепление эпидермиса к базальной пластинке, на коже образуются заполненные серозным содержимым пузыри.
Примечания
См. также
Ссылки
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Промежуточные филаменты" в других словарях:
ФИЛАМЕНТЫ — (от позднелат. filamentum нитевидное образование, нить), общее назв. внутриклеточных цитоплазматич. фибриллярных (нитеподобных) белковых структур. Электронно микроскопически различают 3 класса Ф: микрофиламенты; толстые Ф. миозиноподобные,… … Биологический энциклопедический словарь
Цитоскелет — эукариот. Актиновые микрофиламенты окрашены в красный, микротрубочки в зелёный, ядра клеток в голубой цвет. Цитоскелет это клеточный каркас или скелет, находящийся в цитоплазме живой клет … Википедия
Кресцентин — Модели белка кресцентина из Caulobacter vibrioides Q … Википедия
Структурная функция белков — заключается в том, что белки участвуют в образовании практически всех органоидов клеток, во многом определяя их структуру (форму); образуют цитоскелет, придающий форму клеткам и многим органоидам и обеспечивающий механическую форму ряда тканей;… … Википедия
Кератины — Микроскопическое изображение нитей кератина внутри клетки Кератины семейство фибриллярных белков, обладающих механической прочностью, которая среди материалов биологического происхождения уступает лишь хитину. В основном из кератинов… … Википедия
Клетка — У этого термина существуют и другие значения, см. Клетка (значения). Клетки крови человека (РЭМ) … Википедия
Десмосомы — desmosomes … Википедия
СОЕДИНИТЕЛЬНАЯ ТКАНЬ — главная опорная и защитная ткань организма, основа всех его связующих и опорных структур. В широком смысле это несколько разных тканей, образующих соединительнотканные структуры кости, сухожилия, связки, суставы, дерму и кровеносные сосуды,… … Энциклопедия Кольера
Межклеточные контакты — соединения между клетками, образованные при помощи белков. Межклеточные контакты обеспечивают непосредственную связь между клетками. Кроме того, клетки взаимодействуют друг с другом на расстоянии с помощью сигналов (главным образом сигнальных… … Википедия
Межклеточные соединения — – соединения между клетками, образованные при помощи белков. Остальные виды взаимодействий клеток проходят посредством соединительной ткани. Межклеточные связи сводятся не только к электрическим взаимодействиям. Взаимосвязь между клетками… … Википедия
Читайте также: