Эндотоксины. Эндотоксины микроорганизмов. Эндотоксиновый шок. Эндотоксинемия. Экзоферменты. Суперантигены.
Добавил пользователь Алексей Ф. Обновлено: 27.01.2026
резистентності організму може викликати гнійні отити, пневмонії, перитоніти, абсцеси.
м'яких тканин і навіть сепсис (нерідко зі смертельними наслідками).
Багато вчених робили спроби провести чіткі грані між патогенними, умовно-
патогенними, або мікробами-опортуністами, і непатогенними мікроорганізмами, але ці
грані настільки розмиті і настільки важливу роль відіграє стан макроорганізму в
виникнення інфекції, що на рівні сучасних знань це навряд чи можливо.
В даний час у світовій літературі з'явився термін Іклінічна мікробіологія⌡, під
яким мають на увазі той розділ мікробіології, який вивчає інфекційні процеси,
Збудниками таких гнійно-запальних захворювань людини можуть бути представники
нормальної мікрофлори людського тіла та мікроорганізми, що мешкають у навколишньому середовищі,
які мають слабку патогенність для людини: всі стафілококи, багато стрептококи,
деякі нейссерії, ешерихії, клебсієли, протей, енте-
робактери, цитобактери, псевдомонади, бактероїди, гриби та ін. Ці слабопатогенні
мікроорганізми можуть викликати гнійно-запальні захворювання у тих випадках, коли їх
концентрація дуже велика, а природна опірність макроорганізму різко знижена.
Люди зі зниженою резистентністю називаються імунокомпромісними господарями (від англ.
immune compromised host). Цей стан може бути викликаний тривалим захворюванням
(хронічні пневмонія, пієлонефрит та ін.), широким хірургічним втручанням,
онкологічними хворобами, вродженими імунодефіцитами тощо. У багатьох випадках
лікарем), оскільки неправильні дії лікаря можуть спровокувати хворобу.
Необґрунтоване призначення гормональних препаратів, антибіотиків може викликати
імунодепресивний стан, не кажучи про ті випадки, коли лікар змушений пригнічувати
імуногенез для запобігання відторгненню трансплантату (наприклад, пересадка нирки) або
в інших випадках.
Велика кількість імунокомпромісних господарів знаходиться у різних стаціонарах
(хірургічні відділення, опікові центри, відділення реанімації та інтенсивної терапії,
відділення недоношених дітей тощо). Тому опортуністичні інфекції зазвичай носять
характер госпітальних інфекцій. Зазвичай рівень захворюваності на госпітальні інфекції
коливається від 5 до 500 на 10 000 госпіталізованих. Тому опортуністичні
інфекції є серйозною проблемою сучасної клінічної медицини у всьому
Розповсюдженню госпітальних інфекцій чимало сприяють антисанітарні умови
перебування хворих у стаціонарах, похибки медичних працівників в асептиці та
антисептиці. Часто медичний інструментарій та апаратура не піддаються достатньою
дезінфекції та стерилізації, іноді умовно- патогенні бактеріїможна виявити навіть на
перев'язувальному та шовному матеріалі, у готових лікарських формах.
У деяких випадках умовно-патогенні мікроорганізми кон-тамінують (заселяють)
лікарняну апаратуру, що служить для обстеження та лікування хворих, або аптечні
прилади для приготування лікарських форм. Наприклад, контамінація
псевдомонадами дистилятора призводить до того, що дистильована вода
використовують для приготування багатьох лікарських засобів для хворих, що містить P.
aemginosa і разом з лікарським засобомпотрапляє до організму хворого.
Діагностика опортуністичних інфекційпов'язана з багатьма труднощами через різноманіття
локалізації запальних процесіві, отже, симптоматика захворювань. Крім
того, виявлення у гноє або мокротинні стафілокока не є доказом того, що
стафілокок викликав це захворювання, оскільки стафілокок (як і більшість інших
умовно-патогенних мікроорганізмів) є представником нормальної мікрофлори
людського тіла. Для доказу етіологічної значущості виділених
мікроорганізмів-опортуністів необхідні кількісні дослідження, що доводять
високу концентрацію даного мікроорганізму у досліджуваному субстраті. Іншим
доказом етіологічної значущості виділеної культури може бути
позитивна сероконверсія: наростання титрів антитіл до цього мікроорганізму при
спостереженні за хворим протягом 2.3 тиж. Одноразове дослідження титру антитіл не може
служити доказом етіологічної значущості, оскільки діагностичні титри
невідомі через їх варіабельність у різних людей. Зазвичай титри антитіл до представників
нормальної мікрофлори людини дуже низькі і не служать цілям діагностики.
Більшість клінічних мікробіологічних лабораторій та лабораторій санітарно-
епідеміологічних станцій мало використовують техніку анаеробних посівів, тому рідко
виявляють у досліджуваному матеріалі облігатних анаеробних збудників гнійно-
запальних захворювань. Одні з найпоширеніших нормальних мешканців
кишечника людини бактерії роду Bacteroides нерідко у поєднанні з іншими мікробами-
опортуністами викликають локальні абсцеси черевної порожнини, порожнини малого тазу,
Умовно-патогенні бактерії можуть бути виявлені не тільки в патологічному матеріалі,
взятому від хворого, а й у змивах із предметів довкілля: халатів, рук персоналу
лікарні або аптеки, інструментів, посуду, лабораторної апаратури, перев'язувального та
шовного матеріалу готових лікарських форм. Змив виробляють стерильним тампоном,
змоченим стерильним ізотонічним розчином хлориду натрію, з наступним посівом на
середовища накопичення, диференціально-діагностичні та елективні середовища. Виділені чисті
культури ідентифікують до вигляду. На вимогу лікарів-епідеміологів, що лікують, лабораторія
проводить додаткові дослідження щодо визначення чутливості виділених бактерій
до антибіотиків (антибіотикограма) або фагів, бактеріоцинів та інших маркерів для
розшифровки епідеміологічної ситуації у стаціонарі.
У цю групу входять патогенні та умовно-патогенні для людини круглі форми бактерій.
Серед них є суворі анаероби (пептококи, пептострептококи, вейлонели),
факультативні анаероби та аероби (стафілококи, стрептококи, нейс-серії). Усі вони можуть
викликати у людини гнійно-запальні захворювання, що відрізняються за локалізації та
Стафілококи. Відділ Firmicutes, сімейство Micrococcaceae, рід Staphylococcus. У рід
Staphylococcus за класифікацією Байрд. Паркер входять 3 види: S. aureus, S. epidermidis та
S. saprophyticus. Запропоновані нещодавно інші класифікації включають більшу кількість
видів стафілококів, але вони використовуються поки що тільки в наукових дослідженнях.
Всі види стафілококів є округлими клітинами діаметром 0,5.1 мкм. У мазку
розташовуються зазвичай несиметричними гронами (Ігрозді винограду), але зустрічаються
поодинокі клітини, пари клітин. Грампозитивні. Суперечку не утворюють, джгутиків не мають. У
деяких штамів можна знайти капсулу. Можуть утворювати L-форми. Клітинна стінка
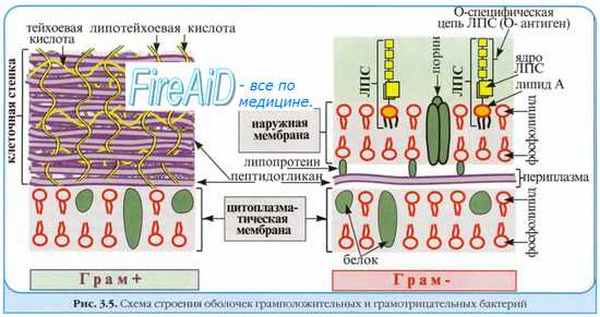
містить велику кількість пептидоглікану, пов'язаних з ним тейхоєвих кислот, протеїн А.
Стафілококи добре ростуть на простих середовищах (рН 7,0.7,5); факультативні анаероби. на
щільних середовищах утворюють гладкі круглі опуклі колонії з різним пігментом. Пігмент
не має таксономічного значення. Можуть зростати на агарі з високим вмістом (8.10%)
NaCl. Продукують сахаролітичні та протеолітичні ферменти. Стафілококи
виробляють гемолізини, фібринолізин, фосфатазу, р-лактамазу, бактеріоцинін,
ентеротоксини, коагулазу, ДНК-азу, лейкоциди-ни, лецитовітелазу та ін.
Стафілококи дуже пластичні: швидко виробляють стійкість до антибактеріальних
препаратів. Істотну роль у цьому відіграють плазміди, що передаються за допомогою трансдуци-
фагів від однієї клітини до іншої. R-плазміди детермінують стійкість до одного
або декільком антибіотикам, у тому числі і за рахунок екстрацелюлярної продукції р-лакта-
мази - ферменту, що руйнує пеніцилін, що розриває його р-лактамне кільце.
Антигенна структура стафілококів складна та варіабельна. Відомо близько 30 антигенів,
білки, полісахариди, тейхоеві кислоти. Протеїн А має
властивістю міцно зв'язуватися із Fc-фрагментом будь-якої молекули IgG. При цьому Fab-фрагмент
молекули імуноглобуліну залишається вільним і може поєднуватися зі специфічним
антигеном. У зв'язку з цією властивістю протеїн А знайшов застосування в діагности-
чеських методах (коаглютинація). Більшість позаклітинних речовин, що продукуються
стафілококами, також має антигенну активність.
Чутливість до бактеріофагів (фаготип) є стабільною генетичною.
характеристикою, що базується на поверхневих рецепторах Багато штами стафілококів
є лізогенними. З а-токсину (екзотоксин) можна приготувати анатоксин.
При мікробіологічній діагностиці віднесення культури до роду стафілококів ґрунтується
на типовій морфології та забарвленні клітин, їх взаємному розташуванні та анаеробному
ферментації глюкози Для видової ідентифікації використовують переважно 3.4 тести:
продукцію плазмокоагулази, лецитовітел-лази, анаеробну ферментацію маніту та глюкози. У
В сумнівних випадках проводять тести на наявність ДНК-ази, а-токсину.
Збудником стафілококових інфекційчастіше буває S. aureus, дещо рідше - - S.
Епідермідіс, дуже рідко. S. saprophyticus. Стафілококи є представниками
нормальної мікрофлори людського тіла, тому мікробіологічна діагностика
стафілококових інфекцій не може обмежитися виділенням та ідентифікацією збудників;
необхідні кількісні методи дослідження, тобто визначення числа мікроорганізмів у
Лікування стафілококових інфекцій зазвичай проводять антибіотиками та сульфаніламідними.
препаратами. У Останніми рокамивід хворих часто виділяють стафілококи, резистентні до
більшості хіміотерапевтичних препаратів. У таких випадках для лікування використовують
антитоксичну протистафілококову плазму або імуноглобулін, отримані з крові
донорів, імунізованих стафілококовим анатоксином Для активної імунізації
(планових хірургічних хворих, вагітних жінок) може бути використаний
адсорбований стафілококовий анатоксин.
Стрептококи. Відділ Firmicutes, сімейство Streptococcaceae, рід Streptococcus. У рід
Streptococcus входять понад 20 видів, серед яких є представники нормальної
мікрофлори людського тіла та збудники важких інфекційних епідемічних
Стрептококи. дрібні (менше 1 мкм) кулясті клітини, що розташовуються ланцюжками або
попарно, грампозитивні, суперечка не утворюють, нерухомі. Більшість штамів
стрептококів утворюють капсулу, що складається з гіалуронової кислоти. Клітинна стінка
містить білки (М-, Т-і R-антигени), вуглеводи (группоспецифічні) і пептидоглікани.
Легко переходять у L-форми.
Зростають на середовищах, збагачених вуглеводами, кров'ю, сировато-
ної, асцитичної рідиною. На щільних середовищах зазвичай формують дрібні сірі колонії.
На рідких середовищах для стрептс коків характерний придонний ріст. Стрептококи. факульту
тивні анаероби. На кров'яному агарі викликають а-гемоліз (зе лінивий) і р-гемоліз (повний).
Гинуть при пастеризації при 56 °С протягом 30 хв.
Генетичний обмін можливий за рахунок трансформації та трансдукції, але не кон'югації.
Эндотоксины. Эндотоксины микроорганизмов. Эндотоксиновый шок. Эндотоксинемия. Экзоферменты. Суперантигены.
Эндотоксины. Эндотоксины микроорганизмов. Эндотоксиновый шок. Эндотоксинемия. Экзоферменты. Суперантигены.
В определённой степени токсигенным микроорганизмам (активно секретирующими токсины) противопоставлены патогенные бактерии, обладающие токсическими субстанциями, слабо диффундирующими в окружающую среду и названные (по предложению Р. Пфайффера) эндотоксинами.
Эндотоксины — интегральные компоненты клеточной стенки грамотрицателъных бактерий; большая их часть высвобождается только после гибели бактериальной клетки. Представлены комплексом протеинов, липидных и полисахаридных остатков. За проявление биологического эффекта ответственны все группировки молекулы эндотоксина.
Биологическая активность напоминает таковую у некоторых медиаторов воспаления; эндотоксинемия обычно сопровождается лихорадкой, обусловленной выбросом эндогенных пирогенов из гранулоцитов и моноцитов. При попадании значительного количества эндотоксина в кровоток возможен эндотоксиновый шок, обычно заканчивающийся смертью больного.
Бактериальные эндотоксины проявляют сравнительно слабое иммуногенное действие, и иммунные сыворотки не способны полностью блокировать их токсические эффекты. Некоторые бактерии могут одновременно синтезировать экзотоксины и выделять (при гибели) эндотоксины (например, токсигенные Escherichia coli и холерные вибрионы).
Экзоферменты
Важными факторами патогенности следует считать экзоферменты (например, лецитиназа, гиалуронидаза, коллагеназа и др.), нарушающие гомеостаз клеток и тканей, что приводит к их повреждению. Способность к образованию экзоферментов во многом определяет инвазивность бактерий — возможность проникать через слизистые оболочки, соединительнотканные и другие барьеры. Например, гиалуронидаза расщепляет гиалуроновую кислоту, входящую в состав межклеточного вещества, что повышает проницаемость различных тканей. Этот фермент синтезируют бактерии родов Clostridium, Streptococcus, Staphylococcus и др, Нейраминидаза облегчает преодоление слоя слизи, проникновение внутрь клеток и распространение в межклеточных пространствах. Нейраминидазу секретируют холерные вибрионы, дифтерийная палочка; он также входит в состав вируса гриппа. К этой же группе следует отнести и бактериальные ферменты, разлагающие антибиотики.
Суперантигены
Некоторые токсины (например, токсин Дика стрептококков или энтеротоксин стафилококков) способны действовать как суперантигены, вызывая поликлональную активацию различных клонов лимфоцитов. Поликлональная активация сопровождается гиперсекрецией лимфокинов с развитием цитокинопосредованной интоксикации.
- Вернуться в оглавление раздела "Микробиология."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Шпаргалка по "Микробиологии"
51.
Инфе́кция — заражение живых организмов бактериями или вирусами, или грибами, или простейшими. В медицине термин инфекция означает различные виды взаимодействия чужеродных микроорганизмов с организмом человека и животных.
Работа содержит 1 файл
ответы на вопросы с 51 (мои).docx
• Наиболее распространённая классификация экзотоксинов основана на характере мишеней для их эффектов: нейротоксины поражают клетки нервной ткани, гемолизины разрушают эритроциты, энтеротоксины поражают эпителий тонкого кишечника, дерматонекротоксины вызывают некротические поражения кожных покровов, лейкоцидины повреждают фагоциты (лейкоциты) и т.д.
• По механизму действия среди экзотоксинов выделяют цитотоксины (например, энтеротоксины или дерматонекротоксины), мембранотоксины (например, гемолизины и лейкоцидины), функциональные блокаторы (например, холероген), эксфолиатины и эритрогенины. Нередко патогенные бактерии синтезируют несколько экзотоксинов, проявляющих различное действие (летальное, гемолитическое, цитотоксическое и т.д.).
2)Эндотоксины — интегральные компоненты клеточной стенки грамотрицателъных бактерий; большая их часть высвобождается только после гибели бактериальной клетки. Представлены комплексом протеинов, липидных и полисахаридных остатков. За проявление биологического эффекта ответственны все группировки молекулы эндотоксина.
Биологическая активность напоминает таковую у некоторых медиаторов воспаления; эндотоксинемия обы чно сопровождается лихорадкой, обусловленной выбросом эндогенных пирогенов из гранулоцитов и моноцитов. При попадании значительного количества эндотоксина в кровоток возможен эндотоксиновый шок, обычно заканчивающийся смертью больного.
Бактериальные эндотоксины проявляют сравнительно слабое иммуногенное действие, и иммунные сыворотки не способны полностью блокировать их токсические эффекты. Некоторые бактерии могут одновременно синтезировать экзотоксины и выделять (при гибели) эндотоксины (например, токсигенные Escherichia coli и холерные вибрионы).
Экзоферменты
Важными факторами патогенности следует считать экзоферменты ( например, лецитиназа, гиалуронидаза, коллагеназа и др.), нарушающие гомеостаз клеток и тканей, что приводит к их повреждению. Способность к образованию экзоферментов во многом определяет инвазивность бактерий — возможность проникать через слизистые оболочки, соединительнотканные и другие барьеры. Например, гиалуронидаза расщепляет гиалуроновую кислоту, входящую в состав межклеточного вещества, что повышает проницаемость различных тканей. Этот фермент синтезируют бактерии родов Clostridium, Streptococcus, Staphylococcus и др, Нейраминидаза облегчает преодоление слоя слизи, проникновение внутрь клеток и распространение в межклеточных пространствах. Нейраминидазу секретируют холерные вибрионы, дифтерийная палочка; он также входит в состав вируса гриппа. К этой же группе следует отнести и бактериальные ферменты, разлагающие антибиотики.
Суперантигены
Некоторые токсины (например, токсин Дика стрептококков или энтеротоксин стафилококков) способны действовать как суперантигены, вызывая поликлональную активацию различных клонов лимфоцитов. Поликлональная активация сопровождается гиперсекрецией лимфокинов с развитием цитокинопосредованной интоксикации.
Ферменты [от лат. fermentum, закваска], или энзимы [от греч. enzyme, дрожжи или закваска], — специфичные и эффективные белковые катализаторы, присутствующие во всех живых клетках. За каждое превращение одного соединения в другое ответственен особый фермент.
• Ферменты снижают энергию активации, обеспечивая протекание таких химических реакций, которые без них могли бы проходить только при высокой температуре, избыточном давлении и при других нефизиологических условиях, неприемлемых для живой клетки.
• Ферменты увеличивают скорость реакции примерно на 10 порядков, что сокращает полупериод какой-либо реакции с 300 лет до одной секунды.
• Ферменты «узнают» субстрат по пространственному расположению его молекулы и распределению зарядов в ней. За связывание с субстратом отвечает определённый участок молекулы ферментативного белка — его каталитический центр. При этом образуется промежуточный фермент-субстратный комплекс, который затем распадается с образованием продукта реакции и свободного фермента.
• Регуляторные (аллостерические) ферменты воспринимают различные метаболические сигналы и в соответствии с ними изменяют свою каталитическую активность.
• Эффекторные ферменты. Известно шесть основных классов ферментов, катализирующих следующие реакции: оксидоредуктазы — перенос электронов; трансферазы — перенос различных химических групп; гидролазы — перенос функциональных групп на молекулу воды; лиазы — присоединение групп по двойным связям и обратные реакции; изомера-зы — перенос групп внутри молекулы с образованием изомерных форм; лигазы — образование связей С-С, C-S, С-О, C-N за счёт реакций конденсации, сопряжённых с распадом аденозинтрифосфата (АТФ).
• Бактерии способны синтезировать все ферменты, необходимые для утилизации широкого спектра питательных субстратов. Определённый субстрат в среде вызывает синтез ферментов, обеспечивающих его катаболизм. В этом случае говорят об индукции катаболических ферментов индуцирующим субстратом (иидуцибельные ферменты). Образование анаболических ферментов в процессах биосинтеза регулируется путём репрессии конечным продуктом (репрессибельные ферменты). Если в среде имеются одновременно два субстрата, то бактерия использует субстрат, обеспечивающий более быстрый рост. Синтез ферментов для расщепления второго субстрата репрессируется; такой вариант известен как катаболитная репрессия. Ферменты, синтезируемые вне зависимости от условий среды, — конститутивные ферменты.
Определение ферментативной активности бактерий
• Определение ферментативной активности бактерий играет огромную роль в их идентификации. Например, все аэробы или факультативные анаэробы обладают супероксид дисмутазой и каталазой — ферментами, защищающими клетку от токсичных продуктов кислородного метаболизма. Практически все облигатные анаэробы не синтезируют эти ферменты. Только одна группа аэробных бактерий — молочнокислые бактерии каталазонегативны, но аккумулируют пероксидазу — фермент, катализирующий окисление органических соединений под действием Н202 (восстанавливается до воды). Наличие аргининдигидролазы — диагностический признак, позволяющий различить сапрофитические виды Pseudomonas от фитопатогенных. Среди пяти основных групп семейства Enterobacteriaceae только две — Escherichiae и Erwiniae— не синтезируют уреазу. Часто вирулентность штамма связана с повышенной активностью ферментов, ответственных за синтез токсинов.
• Получение микробных ферментов — важнейшая отрасль промышленной микробиологии. Например, для улучшения пищеварения применяют готовые препараты ферментов — амилазы, целлюлазы, протеазы, липазы, облегчающих соответственно гидролиз крахмала, целлюлозы, белка и липидов. При изготовлении сладостей для предупреждения кристаллизации сахарозы применяют инвертазу дрожжей, для осветления фруктовых соков — пектиназу. Коллагеназа клостридий и стрептокиназа стрептококков, гидролизующие белки, способствуют заживлению ран и ожогов. Литические ферменты бактерий, секретируемые в окружающую среду, действуют на клеточные стенки патогенных микроорганизмов и служат эффективным средством в борьбе с последними, даже если они обладают множественной устойчивостью к антибиотикам. В качестве инструментария в биоорганической химии, генной инженерии и генотерапии используют выделенные из бактерий рибонуклеазы, дезоксирибонуклеазы, полимеразы, ДНК-лигазы и прочие ферменты, направленно модифицирующие нуклеиновые кислоты.
Ферменты агрессии и защиты.
Для осуществления колонизации и инвазии многие бактерии выделяют ферменты агрессии и защиты:
-протеазы, действие которых, в первую очередь, направлено на разрушение антител;
-лецитовителлаза - лецитиназа, разрушает клеточные мембраны;
-плазмокоагулаза - способствует образованию фибриновых барьеров;
-антифагин - липополисахарид, оказывающий токсическое действие на фагоциты;
-фибринолизин - протеолитический фермент, который растворяет сгустки фибрина;
-гиалуронидаза -фермент, гидролизующий гиалуроновую кислоту - основной компонент соединительной ткани;
-нейраминидаза - отщепляет от различных гликопротеидов, гликолипидов, полисахаридов сиаловую (нейраминовую) кислоту, повышая проницаемость различных тканей.
Три последних фермента облегчают распространение микроорганизмов в тканях организма.
1)Капсулы «экранируют» бактериальные структуры, активирующие систему комплемента, а также структуры, распознаваемые иммунокомпетентными клетками. Например, слой капсульно-го вещества защищает тейхоевые кислоты стафилококков от связывания опсонинами.
Разнообразные микробы хорошо выдерживают воздействие защитных факторов и колонизируют нестерильные полости различных организмов. К длительному выживанию в составе микробных сообществ способны не только сапрофитические и условно-патогенные (формирующие нормальную микрофлору), но и некоторые паразитические виды.
• В ряде случаев развивающиеся иммунные реакции лишают возбудитель возможности полностью реализовать свои патогенные свойства, но оказываются бессильными удалить его из организма. Таким путём формируется бактерионосительст во.
• Патогенез ряда инфекционных болезней в качестве ведущего фактора включает длительную циркуляцию возбудителя (например, возбудителей сифилиса, туберкулёза, бруцеллёза и др.). Длительное выживание в организме хозяина во многом определяет способность патогенов «уходить» от действия защитных факторов (например, за счёт изменения антигенной структуры либо путём антигенной мимикрии).
Антигены — вещества различного происхождения, несущие признаки генетической чужеродное™ и вызывающие развитие иммунных реакций (гуморальных, клеточных, состояние иммунной толерантности, индуцирование иммунной памяти). Свойства Аг определяются комплексом признаков: иммуногенность, антигенность, специфичность, чужеродность.
• Иммуногенность — способность индуцировать иммунный ответ.
• Антигенность — способность Аг избирательно реагировать со специфичными к нему AT или Аг-распознающими рецепторами лимфоцитов. С понятием «антигенность» связан другой термин «чужеродность»: без чужеродности нет антигенности применительно к конкретному организму. Например, альбумины мыши не проявляют антигенные свойства по отношению к другим мышам, но являются Аг для морской свинки.
• Специфичность — структурные особенности, отличающие один Аг от другого.
Способностью вызывать развитие иммунного ответа и определять его специфичность обладает фрагмент молекулы Аг — антигенная детерминанта(эпитоп), избирательно реагирующая с Аг-распознающими рецепторами и AT. Антигенные детерминанты располагаются в областях Аг, обращенных к его микроокружению.
Эпитоп — наименьшая распознаваемая единица Аг; молекула Аг может иметь несколько эпитопов, то есть быть поливалентной. Чем сложнее молекула Аг и чем больше у неё эпитопов, тем больше вероятность развития иммунного ответа. Структура многих антигенных детерминант известна. Например, в полипептидной последовательности эпитопом может быть фрагмент из 7-8 аминокислотных остатков; свойства антигенности и специфичности определяются также пространственной конфигурацией фрагмента.
Моноклональные AT специфически распознают только одну Аг-детерминанту и связываются с ней. Поликлональные AT, как правило, распознают несколько антигенных детерминант в составе Аг.
Валентность Аг. Белки содержат несколько Аг-детерминант. Количество молекул AT, связывающих все эпитопы, определяет валентность Аг (возрастает пропорционально увеличению молекулярной массы белковой молекулы).
Антигены ( Аг ) разделяют на иммуногены, гаптены и толерогены.
Иммуногены. Большая часть Аг способна запускать иммунные реакции, выступая в последующем в качестве мишени, в отношении которой эти реакции реализуются. Иначе иммуногены известны как полные Аг. Часть Аг имеют малые размеры и простое строение, тогда как другие представляют крупные и сложные молекулы, содержат множество эпитопов, каждый из которых распознают различные рецепторы лимфоцитов и/или AT.
Гаптены [от греч. hapto, прикрепляться] обладают антигенностью (то есть взаимодействуют со специфическими AT), но не иммуногенны - (то есть не способны запускать иммунные реакции). Иначе гаптены известны как неполные Аг. Как правило, они имеют небольшую молекулярную массу и не распознаются имму некомпетентным и клетками.
Гаптены могут быть простыми и сложными; простые гаптены взаимодействуют с AT в организме, но не способны реагировать с ними in vitro; сложные гаптены взаимодействуют с AT in vivo и in vitro. Гаптены могут стать иммуногенными при связывании с высокомолекулярным носителем, обладающим собственной иммуногенностью. Например, хром и никель, связываясь с белками кожи, способны вызвать аллергический контактный дерматит, развивающийся при повторных соприкосновениях кожи с хромированными или никелированными предметами. При этом антигенные детерминантыгаптена полностью маскируют аналогичные структуры носителя.
Непреципитирующие гаптены взаимодействуют с AT, блокируют их, но не образуют видимых преципитатов. AT, связавшиеся с такими гаптенами, не реагируют с полными Аг, вызывающими образование AT.
Преципитирующие гаптены образуют видимые преципитаты при взаимодействии со специфическими AT. Свойствами преципитирующих гаптеновобладают полисахариды энтеробактерии и пневмококков.
Полугаптены — неорганические вещества (например, йод или хром), присоединение которых к молекуле белка меняет его иммуногенные свойства. Образующиеся AT специфичны к йоду или хрому, то есть к детерминантам на поверхности полного Аг, но не к белку-носителю.
Проантигены — гаптены, способные присоединяться к белкам организма и сенсибилизировать его как аутоантигены. Например, метаболиты грибов пенициллов или продукты распада пенициллинов могут связывать белки и вызывать развитие к ним иммунных реакций.
Генетический контроль патогенности и вирулентности
Все вышеназванные факторы и параметры патогенности и вирулентности подвержены фенотипическим и генотипическим изменениям. Причины таких изменений — эффектыразличных физических и химических факторов. В первую очередь патогенные свойства бактерии находятся под контролем хромосомных и плазмидийных генов. Способность к образованию экзотоксинов детерминируют внехромосомные tox-гены конвертирующих бактериофагов и плазмид (например, синтез дифтерийного гистотоксина, ботулинического нейротоксина и др.). Образование эндотоксинов кодируют хромосомные гены.
Генотипическое снижение вирулентностивозможнопри мутациях, рекомбинациях, утере внехромосомных наследственных факторов (плазмид, транспозонов, вставочных (IS-) последовательностей).
Фенотипическое снижение вирулентностивозможно при попадании возбудителя в неблагоприятные условия. In vitro оно возникает в результате неблагоприятного режима культивирования и состава питательной среды, воздействия селективных неблагоприятных факторов либо обработки популяции гомологичной антисывороткой. In vivo снижение вирулентности возникает вследствие селекции маловирулентных штаммов в гетерогенной популяции возбудителя под действием защитных факторов, антимикробных препаратов и др. Выжившая популяция приобретает устойчивость к этим воздействиям, но «платит» своими патогенными свойствами (например, за счёт утери плазмидных или хромосомных генов патогенности).
Со времён Пастёра искусственное снижение вирулентности — аттенуация [от лат. attenuo, ослаблять] — положено в основу производства ряда вакцин. Таким образом, патогенность — качественный, признак болезнетворного микроба, а вирулентность — количественное проявление патогенности.
Основные факторы патогенности
К основным факторам патогенности (вирулентности) относят способность микроорганизмов к колонизации, их устойчивость к разным микробицидным факторам организма, свойства инвазивности и токсигенности, а также способность к длительному персистированию.
Способность к колонизации
Адгезия
Размножению бактерий в первичном очаге инфицирования предшествует адгезия [от лат. adhaesio, прикрепляться к чему-либо], то есть закрепление бактерий на поверхности клеток, что, собственно, и служит началом инфекционного процесса. Прикрепление к поверхности клеток (например, к эпителию слизистых оболочек) обеспечивают адгезины, или факторы колонизации— различные микробные продукты — молекулы адгезии (белки, ЛПС, липо-тейхоевые кислоты). Молекулы адгезии могут располагаться непосредственно на поверхности бактериальной клетки либо входить в состав микроворсинок или капсул. Взаимодействие инфекционного агента с эпителиальными клетками происходит в результате нескольких типов связей, различных по природе и специфичности. Выделяют связи, основанные на взаимодействии электростатических сил, обусловленные гидрофобными свойствами поверхности, лиганд-рецепторные взаимодействия.
Заряд. Бактериальные и эукариотических клетки заряжены отрицательно, но поверхностные микроворсинки грамотрицательных бактерий снижают заряд бактерий и уменьшают электростатические силы отталкивания.
Гидрофобность. Бескапсульные бактерии обладают высокой гидрофобностью, усиливающей адгезивность; гидрофобные участки обладают сродством к лигандам на поверхности эукариотических клеток, что и приводит к прочности связи.
Специфические взаимодействия.На поверхности бактерий имеются молекулы, способные к стереоспецифичному связыванию с комплементарными молекулами на мембранах эукариотических клеток (например, гемагглютининыили тейхоевые кислоты).
Другие механизмы колонизации.Некоторые бактерии способны «заранее подготавливать» место для дальнейшего размножения; например, нейраминидаза облегчает проникновение холерного вибриона через слой слизи и контакт с сиалосодержащими рецепторами эпителия кишечника. Микроорганизмы также способны сорбироваться на бактериях, уже колонизировавших поверхность слизистых оболочек, либо связывать белки (например, фибронектин), рецепторы к которому имеются на многих клетках макроорганизма. У капсулированных бактерий в прикреплении активно участвуют полисахариды капсулы.
Для успешной колонизации очага первичного инфицирования бактерии должны выдержать действие многочисленных и разнообразных микробицидных факторов хозяина. Для защиты от них микроорганизмы активно используют ряд структур (например, капсулы) и синтезируемых веществ (например, ферменты).
Капсула
Капсула (или её менее выраженный аналог — слизистый слой) ингибирует начальные этапы защитных реакций — распознавание и поглощение.
• Капсулы «экранируют» бактериальные структуры, активирующие систему комплемента, а также структуры, распознаваемые иммунокомпетентными клетками. Например, слой капсульного вещества защищает тейхоевые кислоты стафилококков от связывания опсонинами.
• Гидрофильность капсул затрудняет их поглощение фагоцитами, а само капсульное вещество защищает бактерию от действия лизосомальных ферментов и токсичных оксидантов, выделяемых фагоцитирующими клетками.
• Большое значение имеет лёгкая отделяемость капсул или слизистого слоя от поверхности бактерий. В частности, при поглощении капсулированных бактерий (например, синегнойной палочки), последние легко «снимают с себя» капсулы и избегают прямого контакта с фагоцитом.
Инактивирующие ферменты
Микроорганизмы синтезируют различные ферменты, обезвреживающие многие гуморальные защитные факторы. Например, многие возбудители, особенно паразитирующие на слизистых оболочках, выделяют протеазы, расщепляющие в том числе и молекулы IgA. В инактивировании токсических кислородных продуктов фагоцитов задействованы каталазаи супероксид-дисмутаза. Бактериальные ферменты также способны изменять рН окружающей среды, делая её пригодной для размножения. Например, Helicobacter pylori выделяет уреазу, нейтрализующую кислую среду в желудке.
Инвазивность
Патогенность многих микроорганизмов (например, шигелл) связана с проникновением в эпителиальные клетки, где они размножаются, вызывая нарушение целостности пласта эпителия. Основные факторы, обеспечивающие инвазивность бактерий, — подвижность(обеспечивает проникновение как в клетки, так и межклеточные пространства) и особые клеточные факторы — инвазины[от лат. invasio, проникать, атаковать], способствующие проникновению в эпителиоциты посредством эндоцитоза (например, поверхностные белки грамотрицательных бактерий). Некоторые микроорганизмы проникают за пределы эпителия либо посредством активной инвазии, либо в результате имплантации через различные повреждения кожных покровов. Как разновидность инвазии можно рассматривать способность микроорганизмов диссеминировать из первичного очага инфекции и циркулировать в крови.
Токсигенность
Токсины
Токсины [от трем, toxikon, яд] — важнейшие факторы патогенности, вырабатываемые микроорганизмами и реализующие основные механизмы инфекционного процесса. Роль микробных токсинов в патогенезе инфекционных болезней впервые доказали Э. Ру и А. Иерсён (1888), отделившие «ядовитое начало» возбудителя дифтерии от бактериальных клеток и сумевшие воспроизвести с его помощью клиническую картину болезни у морских свинок.
Экзотоксины
Группа А — токсины, секретируемые во внешнюю среду (например, токсин дифтерийной палочки).
| Экзотоксины | Эндотоксины | |
| Продуцент | Грамположительные и грамотрицательные бактерии | Грамотрицательные бактерии |
| Локализация | Внутри- и внеклеточная | Внутриклеточная |
| Химическая природа | пептиды | Комплексы «белок-ЛПС» |
| Стабильность при 100 0 С | Лабильны | Стабильны |
| Инактивация формальдегидом | Инактивируются | Не инактивируются |
| Нейтрализация гомологичными АТ | Полная | Частичная |
| Биологическая активность | Индивидуальная для каждого токсина | Общая для все токсинов |
| Токсичность | 100-1 000 000 | 0,1 |
Группа В — токсины, частично секретируемые во внешнюю среду и частично ассоциированные с бактериальной клеткой (например, тетаноспазмин столбнячной палочки).
Группа С — токсины, связанные с бактериальной клеткой и высвобождающиеся после её гибели (например, экзотоксины энтеробактерий).
Свойства экзотоксинов
• Экзотоксины обычно содержат бифункциональные (лигандныс и эффскторные) структуры. Первые распознают и связывают комплементарный рецептор (ганглиозиды, белки, гликопротеиды) на мембране клетки, вторые обеспечивают эффекторное действие, наиболее часто — гидролиз НАД до АДФ-рибозы и никотинамида с последующим переносом АДФ-рибозильного остатка на мишени.
• Связывание и проникновение экзотоксинов в определённой степени напоминает механизм действия пептидных и гликопротеиновых гормонов, что обусловлено родством их молекулярных структур. Внутриклеточная мишень для эффекторной части молекулы токсина — обычно жизненно важная система, например биосинтеза белка (для А-токсина синегнойной палочки и шигелл) либо адснилатциклазная система (для холерогена, термолабилыюго токсина кишечной палочки или экзотоксина Bordetella pertussis).
• Наиболее распространённая классификация экзотоксинов основана на характере мишеней для их эффектов: нейротоксиныпоражают клетки нервной ткани, гемолизиныразрушают эритроциты, энтеротоксиныпоражают эпителий тонкого кишечника, дерматонекро-токсинывызывают некротические поражения кожных покровов, лейкоцидиныповреждают фагоциты (лейкоциты) и т.д.
• По механизму действия выделяют цитотоксины(например, энтеротоксины или дерматонекротоксины), мембранотоксины(например, гемолизины и лейкоцидины), функциональные блокаторы(например, холероген), эксфолиатиныи эритрогенины. Нередко патогенные бактерии синтезируют несколько экзотоксинов, проявляющих различное действие (летальное, гемолитическое, цитотоксическое и т.д.).
Эндотоксины
В определённой степени токсигенным микроорганизмам (активно секретирующими токсины) противопоставлены патогенные бактерии, обладающие токсическими субстанциями, слабо диффундирующими в окружающую среду и названные (по предложению Р. Пфайффера) эндотоксинами. Эндотоксины — интегральные компоненты клеточной стенки грамотрицателъных бактерий; большая их часть высвобождается только после гибели бактериальной клетки. Представлены комплексомпротеинов, липидных и полисахаридных остатков. За проявление биологического эффекта ответственны все группировки молекулы эндотоксина. Биологическая активность напоминает таковую у некоторых медиаторов воспаления; эндотоксинемия обычно сопровождается лихорадкой, обусловленной выбросом эндогенных пирогенов из гранулоцитов и моноцитов. При попадании значительного количества эндотоксина в кровоток возможен эндотоксиновый шок, обычно заканчивающийся смертью больного. Бактериальные эндотоксины проявляют сравнительно слабое иммуногенное действие, и иммунные сыворотки не способны полностью блокировать их токсические эффекты. Некоторые бактерии могут одновременно синтезировать экзотоксины и выделять (при гибели) эндотоксины (например, токсигенные Escherichia coli и холерные вибрионы).
Экзоферменты
Важными факторами патогенности следует считать экзоферменты (например, лецитиназа, гиалуронидаза, коллагеназа и др.), нарушающие гомеостаз клеток и тканей, что приводит к их повреждению. Способность к образованию экзоферментов во многом определяет инвазивность бактерий — возможность проникать через слизистые оболочки, соединительнотканные и другие барьеры. Например, гиалуронидаза расщепляет гиалуроновую кислоту, входящую в состав межклеточного вещества, что повышает проницаемость различных тканей. Этот фермент синтезируют бактерии родов Clostridium, Streptococcus, Staphylococcus и др, Нейраминидаза облегчает преодоление слоя слизи, проникновение внутрь клеток и распространение в межклеточных пространствах. Нейраминидазу секретируют холерные вибрионы, дифтерийная палочка; он также входит в состав вируса гриппа. К этой же группе следует отнести и бактериальные ферменты, разлагающие антибиотики.
Суперантигены
Некоторые токсины (например, токсин Дика стрептококков или энтеротоксин стафилококков) способны действовать как суперантигены, вызывая поликлональную активацию различных клонов лимфоцитов. Поликлональная активация сопровождается гиперсекрецией лимфокинов с развитием цитокинопосредованной интоксикации.
Распространение условно патогенных микроорганизмов среди людей. Условно-патогенные микроорганизмы. Факторы патогенности бактерий. Внутрибольничные эковары. Оппортунистические инфекции. Вопросы для самопроверки
Несостоятельность определения "" очевидна, поскольку совершенно ясно, что для возникновения заболевания заражение любым микроорганизмом, в том числе и безусловно-патогенным, должно происходить при определенных условиях. Известно, например, что безусловно-патогенный для грызунов Yersinia pestis даже в больших дозах не вызывает заболевания чумой, если животное находится в состоянии зимней спячки. Нужны определенные условия для возникновения брюшного тифа, дизентерии, холеры и других болезней, вызываемых безусловно-патогенными для человека возбудителями. Такими условиями являются инфицирующая доза возбудителя, пути его проникновения в организм, состояние восприимчивости микроорганизма и др.
Кроме того, следует указать, сколько людей в контактном круге также получает конкретный антибиотик. Например, если дать многим пациентам в том же больничном отделении, то будет сделан решительный отбор для резистентных бактериальных штаммов, поскольку чувствительные также будут уничтожены или уничтожены. Влияние на микробную экосистему станции было бы более благоприятным при использовании разных препаратов.
Поэтому врачи должны не только стремиться эффективно лечить каждую отдельную инфекцию; скорее, использование подобных антибиотиков во всей популяции пациентов всегда должно оставаться ниже тех порогов, при которых устойчивые штаммы уже процветают или чувствительны, что на самом деле являются их конкурентами, разрушаются в массовом масштабе. Таким образом, первоначальная безвредная микрофлора каждого пациента и его среды может быстро восстановиться после окончания лечения.
С другой стороны, непатогенные обитатели кожи и слизистых оболочек человека и животных в экстремальных условиях (при радиоактивном облучении, аглобулинемии, у животных-гнотобионтов, при лечении кортикостероидами, антилимфоцитарной сывороткой, антибиотиками) вызывают тажелые заболевания, т. е. ведут себя как патогенные микроорганизмы.
Трудность, однако, в том, что до сих пор никто не знает, как найти такой предел - кроме того, что в большинстве случаев точных чисел по составу бактериальных популяций нет. С определенными усилиями ученые должны уметь решать оба. С другой стороны, решение проблемы антибиотиков на международном уровне требует глобального сотрудничества между странами, а также совместных усилий по просвещению и просвещению людей о последствиях ненадлежащего поведения. Сейчас есть шаги в этом направлении. Несколько групп теперь пытаются отслеживать появление устойчивых бактериальных штаммов.
Поиски факторов патогенности , которые позволили бы с достаточной достоверностью отличать патогенный микроорганизм от условно-патогенного, также не увенчались успехом. Так, условно-патогенный по общепринятым понятиям Staphylococcus aureus обладает относительно большим набором факторов патогенности (лейкоцидин, коагулаза, экзотоксин, гиалуронидаза, различные гемолизины, фибринолизин, ДПКаза и др.), тогда как у безусловнопатогенной для человека S. typhi этот набор ограничивается эндотоксином.
Она также готовит информационные бюллетени для общественности и работников здравоохранения. Пришло время, чтобы наше глобальное сообщество приняло бактерии как обычные и обычно полезные соседи по дому вместо того, чтобы пытаться их искоренить, - если они не вызывают болезни. Необходимо новое понимание широких последствий использования антибиотиков. Только тогда будет отменено угрожающее развитие сопротивления. В дополнение к лечению нынешних бактериальных заболеваний в центре внимания находится долгосрочное сохранение микробных сообществ.
Тяжесть вызываемого микробом заболевания также не может лежать в основе определения его патогенности. Так, Clostridium tetani - условно-патогенный микроорганизм - выделяет один из самых биологически активных экзотоксинов и вызывает заболевание, которое в досывороточный период давало летальность в 90-100% случаев; при современных методах лечения летальность также достаточно высока. Гнойные менингиты, пневмонии, пиелонефриты, перитониты, сепсис и много других воспалительных болезней, вызываемых условно-патогенными микроорганизмами S. aureus, Streptococcus pneumoniae, Escherichia coli и др., протекают, как правило, тяжело даже при интенсивной специфической химиотерапии.
Очень важно защитить непротивляемые микробы до сих пор, чтобы они смогли выдержать резистентные штаммы. Подобные концепции желательны для контроля над паразитами, грибами и вирусами. Поскольку использование таких препаратов в последнее время резко возросло, проблема резистентности также усиливается в этих патогенах.
Парадокс антибиотика: как чудесные наркотики разрушают чудо. Сопротивление лекарственным средствам: новый апокалипсис. Специальный выпуск к: Тенденции в микробиологии, том 2, выпуск 10, с 341 по 425. Сопротивление антибиотикам: происхождение, эволюция, выбор и распространение.
Л. А. Фаворова, II. Б. Мордвинова (1974) указывают, что особенностью условно-патогенных микроорганизмов является их способность в течение длительного времени, а иногда пожизненно, сохраняться в организме. Однако известно, что и безусловно-патогенные микробы способны длительно сохраняться в организме биологического хозяина. При этом хроническое носительство S. typhi, Vibrio cholerae - далеко не редкое исключение. Многие годы, часто пожизненно способны персистировать в организме человека возбудители туберкулеза, сифилиса, грибковых заболеваний, малярийные плазмодии, гельминты, риккетсии, вирусы кори, гриппа, гепатита В, возбудители так называемых медленных вирусных инфекций. Напротив, некоторые условно-патогенные микроорганизмы быстро выводятся, например, при пищевых токсикоинфекциях.
Медицинские последствия использования антибиотиков в сельском хозяйстве. Вольфганг Витте: Наука, том 279, с 996 по 997. Сдерживание антибиотикорезистентности. Поддерживается смягчение легионеллы в сети распределения питьевой воды. . Факторы, способствующие загрязнению систем водоснабжения легионеллы.
Практически и технически невозможно управлять распределением питьевой воды с помощью простых легионелл или других нежелательных микроорганизмов. Избегать контакта с водой является абсурдным, по крайней мере в течение жизни, при определенных условиях это неизбежно. В то же время необходимо ограничить или полностью устранить контакт с аэрозолями, что также является тупым. Или устройство для медицинских, бытовых или других аэрозольных производств и использование стерильной воды в своей функции с использованием воды из системы.
Для определения понятий «патогенные » и «условно-патогенные » микроорганизмы возможно и целесообразно использовать экологический и филогенетический подходы.
Согласно современной концепции , некоторых возбудителей антропонозов человек получил от своих человекообразных предков, других-от промысловых, синантропных и домашних животных в результате адаптации возбудителей к организму человека (исторические зоонозы), наконец, еще часть-в результате адаптации к организму человека свободноживущих микроорганизмов (сапронозы).
Оценка риска заражения легионеллой возможна только для уменьшения количества легионелл в питьевой воде. Удаление системного загрязнения является необходимым условием ликвидации локального загрязнения. Поэтому предотвращение легионеллеза является ограничением образования аэрозолей, минимизацией появления легионеллы в холодном состоянии и особенно горячей водой до приемлемого уровня.
Характер загрязнения легионеллы. Мы делим загрязнение питьевой воды на местную и систему. Застой воды приводит к температурному ощущению, и это позволяет свергнуть легионеллы. Просто краны, душевые и батарейки показывают гораздо большее количество легионелл, но также и других микроорганизмов, чем циркулирующая горячая вода.
Процесс формирования патогенных для человека (животного) микробов можно представить в виде следующей упрощенной схемы.
Свободноживущие микроорганизмы , автотрофы или гетеротрофы по характеру питания, при случайном попадании в макроорганизм находят здесь благоприятную среду обитания и становятся комменсалами. Этот симбиоз какое-то время является факультативным, но при постоянной циркуляции комменсализм становится облигатным. Характер питания микроба изменяется за счет утраты некоторых и появления новых ферментных систем.
Симметричное загрязнение влияет на центральные секции системы распределения, то есть на всю систему, за исключением выходов и их соединений. Это загрязнение не может быть устранено путем промывки от выходов, дезинфекции, химической и термической дезинфекции, лучшего сочетания обоих. Кроме того, должны быть изменения в эксплуатации, техническом обслуживании и санитарии трубопроводов, возможно, также ряд технических и строительных мер. В противном случае дезинфекция скоро вернется к первоначальному уровню, и долгосрочное поддержание приемлемого загрязнения не может быть гарантировано.
Результатом этого процесса является сложная система биоценозов , «выгодных» для макро- и микроорганизмов. Иногда этот симбиоз принимает форму мутуализма. На коже и слизистых оболочках человека обитают сотни видов микроорганизмов (только в кишечнике-более 260). Нарушение стабильности биоценозов приводит к заболеваниям (дисбактериозам различного характера), отсутствие микробов - к серьезным нарушениям жизнедеятельности (животные-гнотобионты в естественных условиях нежизнеспособны).
То есть, при наличии системного заражения наблюдается тенденция к средней высокой легионелле в воде. Крупные комплексы зданий, гостиниц, больниц, жилых домов, учреждений, библиотек, домов престарелых, санаториев, спортивных и промышленных объектов представляют большой риск с точки зрения легионеллы.
Эпидемиологические аспекты легионеллы Количество изолятов легионеллы с места питьевой воды, выраженное в процентах от положительности вспышек, соответствует заболеваемости. Если положительность больше 30%, случаи легионеллы встречаются в больницах, естественно, ниже этого предела.
Оглавление темы "Патогенность микроорганизмов. Вирулентность.":
1. Патогенность микроорганизмов. Патогенные микроорганизмы. Патогенные микробы.
3. Облигатные паразиты. Факультативные паразиты. Случайные паразиты. Патогенность. Что такое патогенность?
4. Вирулентность. Что такое вирулентность? Критерии вирулентности. Летальная доза (DL, LD). Инфицирующая доза (ID).
5. Генетический контроль патогенности и вирулентности. Генотипическое снижение вирулентности. Фенотипическое снижение вирулентности. Аттенуация.
6. Факторы патогенности микроорганизмов. Факторы патогенности микробов. Способность к колонизации. Адгезия. Факторы колонизации.
7. Капсула как фактор патогенности микроорганизмов. Ингибирующие ферменты микробов как фактор патогенности. Инвазивность микроорганизмов.
8. Токсигенность микроорганизмов. Токсины. Парциальные токсины. Цитолизины. Протоксины.
9. Экзотоксины. Экзотоксины микроорганизмов. Классификация экзотоксинов. Группы экзотоксинов.
10. Эндотоксины. Эндотоксины микроорганизмов. Эндотоксиновый шок. Эндотоксинемия. Экзоферменты. Суперантигены.
Условно-патогенные микроорганизмы , как правило, лишены болезнетворных свойств и не вызывают инфекционных заболеваний у здорового человека. Они нередко колонизируют кожу и слизистые оболочки, но способны и к длительному существованию во внешней среде.
Условно-патогенные микробы вызывают поражения после пассивного переноса во внутреннюю среду организма (например, при нарушении целостности анатомических барьеров). Поскольку эти микроорганизмы лишены тропности к тем или иным тканям, то заболевания не имеют выраженной специфичности и больше зависят от степени поражения органа, чем от патогенных свойств возбудителя. Важные условия их развития - массивность инфицирования и нарушения сопротивляемости организма. Чем более выражены эти нарушения, тем более широкий спектр микроорганизмов способен вызывать инфекционные поражения.
Эта взаимосвязь не показывает кластеризацию явлений, то есть кластеризацию в определенное пространство, местоположение, но случаи не имеют никакого отношения в соответствии с общим распределением легионеллы на сайте объекта. Таким образом, процент позитивных участков участка распределения питьевой воды является лучшим критерием для риска образования легионеллы, чем простая плотность легионелл.
Аэраторы повышают вероятность передачи чувствительным и ослабленным людям. Меры по ликвидации, и в частности химическая, термическая и комбинированная дезинфекция, как правило, имеют лишь краткосрочный эффект и нуждаются в поддержке системы оперативных, и строительных мер.
Обычно даже непатогенные (точнее, не способные вызывать поражения у здорового человека) микроорганизмы находят «возможность» инициировать инфекционный процесс. Их так и называют - оппортунистические патогены [от англ. opportunity, возможность, удобный случай].
Читайте также:
- Кокаин и кокаиновая зависимость. Амфетамины, никотин, кофеин и зависимость от них
- Реоэнцефалография. Реограммы и реография
- Дефицит хрома
- Эволюция непатогенных микроорганизмов. Патогенность в эпидемиологии.
- Классификация рецепторов. Мономодальные и полимодальные рецепторы. Ноцицепторы ( болевые рецепторы ). Экстерорецепторы. Интерорецепторы.