Фабрики репликации ДНК бактерий
Добавил пользователь Владимир З. Обновлено: 29.01.2026
Основные результаты по репликации получены на модельной системе с ДНК вируса SV40, в которой процесс репликации исследовали в зараженных клетках человека, культивируемых in vitro. В этой системе вирусный белок, называемый Т-антигеном, выполняет многие функции, необходимые для репликации вирусной ДНК. Во-первых, он является белком-инициатором, необходимым для инициации репликации; во-вторых, он обладает ДНК-геликазной активностью, т.е. расплетает цепи реплицируемой ДНК перед работающей ДНК-полимеразой, и, в-третьих, Т-антиген необходим для правильного взаимодействия с ДНК ферментного комплекса, синтезирующего праймеры (праймосомы). Тем не менее, вирус SV40 использует для репликации ДНК своей небольшой хромосомы и многие белки клетки-хозяина, что позволяет исследовать функционирование репликативного комплекса клеток человека в такой относительно простой системе.
У эукариот обнаружены шесть ДНК-полимераз, три из которых – α, δ и ε –непосредственно участвуют в репликации хромосомной ДНК. Аминокислотные последовательности этих трех ферментов гомологичны друг другу и последовательности продукта гена 43 бактериофага Т4.
Эукариотическая ДНК-праймаза в отличие от аналогичного белка прокариот образует постоянный комплекс с ДНК-полимеразой α, роль которого, по-видимому, ограничивается синтезом праймеров при репликации обеих цепей ДНК.
Белок PCNA и фактор репликации C (RFС) также образуют стабильный комплекс с ДНК-полимеразой δ, а в определенных условиях стимулируют и активность ДНК-полимеразы ε. Во многих отношениях PCNA и RFС являются функциональными аналогами соответственно β-белка и белков γ-комплекса E. coli, и их роль в синтезе ведущей и отстающей цепей ДНК вируса SV40 хорошо известна.
Механизмы репликации ДНК прокариот и эукариот существенно различаются в том отношении, что во втором случае синтез ведущей и отстающей цепей ДНК осуществляют разные ДНК-полимеразы (α и δ соответственно), тогда как у E. coli обе цепи ДНК синтезируются димером ДНК-полимеразы III. ДНК-полимераза α проводит инициацию синтеза ведущей цепи в точках начала репликации, а ДНК-полимераза δ осуществляет циклические реинициации синтеза фрагментов Оказаки, по-видимому, распознавая наличие 5’-концевого нуклеотида очередного праймера с последующей диссоциацией от матричной ДНК и присоединением к ней для реинициации синтеза следующего фрагмента Оказаки. Созревание фрагментов Оказаки у эукариот требует удаления РНК-затравок с помощью 5’→3’-экзонуклеазы (белковые факторы FEN-1 или MF-1) и РНКазы H1, а также ковалентного соединения фрагментов друг с другом под действием ДНК-лигазы I. Роль ДНК-полимеразы ε в настоящее время не ясна. Возможно, этот фермент непосредственно участвует в репликации или в сопряженной с репликацией репарации повреждений ДНК, а также в регуляции клеточного цикла. ДНК-полимераза ζ обнаружена в 1996 г. у дрожжей S. cerevisiae. При исследовании белков Rev3 и Rev7, которые необходимы для мутагенеза, индуцируемого в ответ на повреждения ДНК, оказалось, что их комплекс обладает ДНК-полимеразной активностью. Эта полимераза способна эффективно использовать в качестве матрицы ДНК, содержащую циклобутановые димеры. В таких условиях активность ДНК-полимеразы α составляет лишь 1% от активности ДНК-полимеразы ζ. ДНК-полимераза η, так же как и предыдущий фермент, участвует в SOS-ответе дрожжей на генотоксические воздействия. В присутствии всех четырех дезоксирибонуклеозидтрифосфатов она осуществляет включение в строящуюся цепь ДНК напротив тиминовых димеров только правильных нуклеотидов (А).
Поскольку ДНК-полимеразы млекопитающих лишены 3'-5'- и 5'-3'-экзонуклеазных активностей, присущих ферментам Е. coli, остается неясно, как в процессе репликации ДНК у этих организмов удаляются случайно включенные ошибочные нуклеотиды и РНК-затравки фрагментов Оказаки.
Синтез лидирующей и отстающей цепи осуществляют разные дНК-полимеразы (альфа, дельта), тогда как у E. coli обе цепи ДНК синтезируются димером ДНК-полимеразы III.
ДНК-полимераза ? проводит инициацию синтеза ведущей цепи в точках начала репликации, а ДНК-полимераза ? осуществляет циклические реинициации синтеза фрагментов Оказаки, по-видимому, распознавая наличие 5’-концевого нуклеотида очередного праймера с последующей диссоциацией от матричной ДНК и присоединением к ней для реинициации синтеза следующего фрагмента
Оказаки. Созревание фрагментов Оказаки у эукариот требует удаления РНК-затравок с помощью 5’>3’-экзонуклеазы (белковые факторы FEN-1 или MF-1) и РНКазы H1, а также ковалентного соединения фрагментов друг с другом под действием ДНК-лигазы I.
ДНК-полимераза ? обнаружена в 1996 г. у дрожжей S. cerevisiae. При исследовании белков Rev3 и Rev7, которые необходимы для мутагенеза, индуцируемого в ответ на повреждения ДНК, оказалось, что их комплекс обладает ДНК-полимеразной активностью. Эта полимераза способна эффективно использовать в качестве матрицы ДНК, содержащую циклобутановые димеры. В таких условиях активность ДНК-полимеразы ? составляет лишь 1% от активности ДНК-полимеразы ?. ДНК-полимераза ?, так же как и предыдущий фермент, участвует в SOS-ответе дрожжей на генотоксические воздействия. В присутствии всех четырех дезоксирибонуклеозидтрифосфатов она осуществляет включение в строящуюся цепь ДНК напротив тиминовых димеров только правильных нуклеотидов (А).
Точки начала репликации репликации
Ori - AT-богатые, легкоплавкие участки длинной ~200 пн, расположенные, как правило, между генами в промоторных областях.
Не все ori реплицируются в одном клеточном цикле.
Ориджины репликации могут располагаться внутри кодирующей части генов, напримр в гене (betta-глобине ципленка находится по крайней мере четыре ориджина репликации, причем было показано что активность генов не связана со степенью репликации. На выбор ориджинов репликации влияет ацетилирование гистонов, хотя данные противоречивы. Например, репликация в бетта-глобиновом локусе ципленка не чувствительна ни к ацетилированию ни к деацетилированию гистонов. В мышином HoxB локусе, ацетилирование гистонов имеет зависимость с молчанием ориджинов скорее чем их активация [Norio, 2006]
Инициация репликации
Инициация репликации у эукариот происходит на специфических множественных последовательностях нуклеотидов – репликаторах. Наиболее изученными являются репликаторы дрожжей S. cerevisiae, впервые идентифицированные как автономно реплицирующиеся последовательности (ARS – autonomously replicating sequence), способные поддерживать внехромосомную репликацию плазмид в дрожжевых клетках. Исследование структуры ARS1 показало, что этот хромосомный элемент состоит из нескольких коротких регуляторных последовательностей. Аналогичная организация характерна и для других ARS дрожжей. В частности, ARS307 в дополнение к канонической последовательности ACS, общей для всех ARS, содержат еще два элемента – B1 и B2, которые необходимы для выполнения репликатором своих функций in vivo. Несмотря на то что эти последовательности в разных репликаторах не строго консервативны, внутри групп (B1, B2 и т.п.) они функционально взаимозаменяемы. Изменение положения по отношению к ACS предотвращает их функционирование.
Первым этапом инициации репликации у дрожжей является взаимодействие регуляторных последовательностей репликатора, по крайней мере, с шестью различными белками, которые образуют комплекс, распознающий область начала репликации ORС (origin-recognition complex). ARS определяет место инициации репликации в клетках дрожжей. Элемент B3 ARS1 взаимодействует с белком Abf1, который стимулирует репликацию доменом, характерным для белков-активаторов транскрипции, тогда как B1 взаимодействует с ORC. Остающиеся регуляторные последовательности области начала репликации дрожжей образуют ранее неизвестный элемент,
названный ДНК-расплетающим элементом DUE (DNA-unwinding element), который, как полагают, облегчает раскручивание цепей ДНК при инициации репликации. Точковые мутации в элементе B2 не влияют на функции репликатора, что является общим свойством структурных элементов, тогда как мутации в ACS, B1 и B3 нарушают инициацию репликации, как и следовало ожидать от регуляторных элементов нуклеиновых кислот, взаимодействующих с белками.
Элонгация репликации
Терминация репликации
Репликация теломерных районов хромосом
Синтез теломерных последовательностей ДНК осуществляется специальными ферментами – теломеразами. Особенностью этих ферментов является присутствие у них в качестве составной части короткого фрагмента РНК – компонента, служащего матрицей при синтезе теломерных последовательностей хромосом. Комплементарное взаимодействие внутренней РНК теломеразы с 3’-концевым выступающим одноцепочечным сегментом ДНК хромосомы инициирует синтез теломерных последовательностей. При этом 3’-концевой фрагмент ДНК служит затравкой для удлинения этой ДНК на РНК-матрице. После удлинения (элонгации) выступающей цепи ДНК до конца матрицы происходит транслокация фермента на один теломерный повтор вперед относительно матрицы с освобождением последовательности матричных нуклеотидов, после чего он готов для вступления в следующий цикл элонгации только что добавленной 3'-концевой последовательности хромосомы. После завершения удлинения одноцепочечной 3'-концевой теломерной последовательности вторая цепь ДНК достраивается обычным способом. Таким образом происходит решение "проблемы отстающей цепи ДНК" при репликации ДНК у эукариот. Наличие у животных тканеспецифичности в распределении теломер по размерам, а также изменение размеров этих последовательностей в онтогенезе предполагают существование механизмов, регулирующих данный процесс. Создается впечатление, что для активной пролиферации клеток теломерные последовательности не должны становиться короче определенного порогового размера. Недавние исследования обнаружили резкое повышение активности теломераз, характерное для опухолевых клеток, что служит в настоящее время чувствительным физиологическим маркером их злокачественного перерождения. В этой связи сегодня в качестве одного из подходов к терапии опухолей рассматривают подавление активности теломераз, функционирование которых, как полагают, необходимо для иммортализации клеток и роста опухолей. Именно благодаря таким свойствам теломеразы в последнее время вызывают особый интерес, что сопровождается расширением исследований в данной области молекулярной генетики.
Регуляция репликации
Пространственная организация репликации
В ядрах млекопитающих при репликации обнаруживается около 150 центров репликации - “репликативные фабрики” или реплисомы, которые приблизительно равномерно удалены друг от друга. Во время инициации синтеза ДНК размер этих центров мал, и они обнаруживаются в виде небольших четко очерченных "точек", которые со временем становятся более диффузными. В данных центрах репликации происходит аккумуляция белков, участвующих в синтезе ДНК: ДНК-полимеразы, PCNA и RP-A, а также регуляторных молекул типа циклина А, cdk2 и RPA70.
Иммунохимическими методами с использованием частиц коллоидного золота было показано, что растущие цепи ДНК выходят из центров репликации. Это позволяет предполагать, что во время репликативного синтеза цепи ДНК перемещаются через фиксированные внутри ядра структуры аппарата репликации. Такая внутриядерная компартментализация синтеза ДНК позволяет концентрировать регуляторные, структурные и ферментативные компоненты, участвующие в репликации и поддержании пространственной структуры хромосом. Ступенчатая сборка функционально активных элонгирующих комплексов в микрокомпартментах ядра предоставляет большие возможности для регуляции инициации репликативного синтеза ДНК.
Роль ядерных пространственных структур в обеспечении функциональных свойств ДНК можно рассмотреть на примере репликации
хромосом в ооцитах Xenopus laevis. Инъекция прокариотической ДНК в ооциты или ее добавление к экстрактам ооцитов сопровождается образованием псевдоядер, компетентных в отношении репликации ДНК. Репликация в таких системах пространственно упорядочена: она происходит в дискретных участках ДНК, содержащих кластеры репликативных вилок. При этом наблюдается замечательная корреляция между числом и пространственным распределением центров репликации в искусственных псевдоядрах и ядрах культивируемых клеток, находящихся в S-фазе. Следовательно, сборка функционирующих комплексов, способных инициировать репликацию, не находится в строгой зависимости от последовательностей нуклеотидов ДНК хромосом, но в большой степени зависит от внутренней пространственной структуры ядра и может эффективно происходить даже на прокариотических ДНК. Это означает, что пространственная структура ядра может непосредственно контролировать его функции, в данном случае – инициацию репликации хромосом. Следует, однако, иметь в виду, что на ранних эмбриональных стадиях развития Xenopus контроль репликации ослаблен, и число зон начала репликации значительно больше, чем в соматических клетках. Нормальный контроль репликации устанавливается после увеличения продолжительности клеточного цикла на стадии средней бластулы. В нормальных соматических клетках не все центры репликации одновременно начинают синтез ДНК. Некоторые из них функционируют в ранней, а некоторые в поздней S-фазе клеточного цикла. Такая дифференциальная репликация хромосом является важным регуляторным механизмом, обеспечивающим локальную организацию хроматина и активность генов. Аналогичное явление обнаружено в клетках дрожжей, где функционирование во времени центров репликации зависит от положения хромосом в ядре.
P. Norio DNA replication: the unbearable lightness of origins EMBO reports 7, 8, 779–781 (2006)
Преподаватели и учебные курсы
Преподаватели и учебные курсы | Институт молекулярной генетики
ЛЕКЦИОННЫЕ КУРСЫ В 2013-2015 ГОДАХ
Программа обновленного курса лекций «Геномика, молекулярная биотехнология и медицина»
Молекулярная биология ДНК – основа биотехнологии (2 часа)
Преподаватель: академик РАН В.А. Гвоздев
Молекула ДНК. ДНК как основа генетической информации. Экспериментальные доказательства генетической функции ДНК. Конформации ДНК (А, В и Z-формы). Нуклеотидный состав ДНК и конформации ДНК. Большая и малые бороздки ДНК. Узнавание ДНК белками в малой и большой бороздке. Подвижность структуры ДНК. Свехспирализация. Неканонические структуры ДНК. Изгибы в ДНК (упаковка ДНК и регуляция транскрипции). Топоизомеры. Топоизомеразы. Полуконсервативная репликация ДНК. ДНК-полимеразы. Вилка репликации ДНК. Регуляция репликации ДНК у бактерий. Понятие о репликоне и репликаторе. Репликация у эукариот. Полирепликонное строение хромосомы. "Фабрики" репликации ДНК в ядре. Клеточный цикл эукариотической клетки. Теломераза и репликация ДНК у эукариот.
Механизмы транскрипции у прокариот (2 часа)
Преподаватель: д.б.н. А.В. Кульбачинский
Структура РНК-полимеразы и механизм синтеза РНК. Инициация синтеза РНК у прокариот, промоторы и опероны. Структура промоторов. Роль сигма-субединицы РНК-полимеразы в узнавании промотора. Разные классы сигма-субъединиц и их роль в переключении активности генов. Регуляция инициации транскрипции с участием белков–активаторов и репрессоров. Регуляция элонгации и терминации транскрипции. Регуляторные белки клетки и бактериофагов, изменяющие активность РНК-полимеразы в процессе элонгации транскрипции. Рибопереключатели и механизмы их действия. Основные методы исследования механизмов работы РНК-полимеразы и транскрипционных факторов. Сравнение основных принципов инициации транскрипции у бактерий и эукариот. Сходство структуры РНК-полимераз бактерий и эукариот, основные типы эукариотических РНК-полимераз. Главные транскрипционные факторы и коактиваторы транскрипции. Принцип действия энхансеров.
Механизмы регуляции транскрипции у эукариот (2 часа)
Преподаватель: к.б.н. Ю.Я. Шевелев
Структура корового промотора РНК-полимеразы II. Базальные факторы транскрипции. Активаторы и репрессоры. Энхансеры и сайленсеры. Хроматин. Нуклеосомная организация. Структура нуклеосомы. Хроматин-ремоделирующие комплексы. N-концевые модификации гистонов. Гистоновый код. Бромо- и хромодомены. Структурные и функциональные хроматиновые домены. Механизмы активации и репрессии транскрипции в домене. Механизм распространения гетерохроматина. Наследование эпигенетических состояний при репликации. Компартментализация генов в ядре как новый уровень регуляции генов.
Механизмы репликации ДНК (2 часа)
Преподаватель: д.б.н., проф. К.В. Северинов
Репликация ДНК хромосом – основа процесса передачи родительских признаков потомству. Регуляция репликации путем синтеза комплементарной нити на каждой из двух цепочек молекулы ДНК. Основные ферментативные и топологические проблемы этого процесса. Структурно–функциональный анализ фермента репликации – ДНК-зависимой ДНК-полимеразы. Минимальный набор белков для процесса репликации. Механизм репликации линейных концов ДНК в теломерной области хромосом.
Химическая коммуникация у бактерий: фундаментальные и прикладные аспекты (2 часа)
Преподаватель: д.б.н. И.А. Хмель
Основные принципы коммуникации бактерий - Quorum Sensing (QS) регуляции экспрессии генов. Типы QS систем и молекулярные механизмы коммуникации бактерий. Сигнальные молекулы, участвующие в функционировании QS систем регуляции. QS системы регуляции как глобальные регуляторы экспрессии генов. Роль QS систем в регуляции клеточных процессов: вирулентности патогенных и фитопатогенных бактерий, синтезе антибиотиков, ферментов, биолюминесценции, антагонистической активности, образовании биопленок. Новая стратегия антимикробной терапии, основанная на использовании ингибиторов
QS регуляции. Методы борьбы с биопленками бактерий – источником трудно излечиваемых хронических инфекций.
Трансгенные животные в биотехнологии (2 часа)
Преподаватель: д.б.н., проф. В.З. Тарантул
Общие понятия о трансгенах и трансгенных организмах. Методы получения трансгенных животных. Структура трансгенов. Механизм трансгеноза. Фундаментальные задачи, решаемые с использованием трансгенных организмов: изучение регуляции экспрессии и функции генов, механизмы эмбрионального развития, получение продуцентов. Инсерционный мутагенез. Токсикогенетика. Эмбриональные стволовые клетки. Химеры и их использование. Генный таргетинг: нокаут генов и генный нокин. Трансгеноз и клонирование животных. Трансгенные животные как биореакторы. Сельскохозяйственные трансгенные животные. Криоконсервация и пересадка ооцитов - новая технология размножения сельскохозяйственных, лабораторных и диких животных и сохранения видов и пород.
Внутриклеточная сигнализация (4 часа)
Преподаватель: д.б.н., проф. И.А. Гривенников
Пути передачи информации в эукариотических клетках. Рецепторы на поверхности эукариотических клеток. Краткая характеристика различных типов рецепторов. G-белки. Вторичные мессенджеры. Система протеинкиназ. Регуляция экспрессии генов. Иерархия регуляции. Факторы транскрипции. Протоонкогены (мембранные, ядерные и цитоплазматические). Роль протоонкогенов в развитии. Факторы роста, краткая характеристика. Молекулярная биология и функции фактора роста нервов в качестве примера. Регуляторные пептиды в качестве регуляторов функций эукариотических клеток.
Медицинская и этническая геномика (2 часа)
Преподаватель: д.б.н., проф. С.А. Лимборская
Геном человека, основные черты организации. Полиморфные маркеры ДНК. Принципы картирования генов наследственных болезней. Прогрессирующая мышечная дистрофия как пример локализации гена на хромосоме. Типы мутаций при наследственных заболеваниях. Динамические мутации, понятие антиципации. Молекулярная диагностика наследственных заболеваний. Полиморфизм генов как инструмент изучения генома. Мультифакториальные заболевания.Болезнь Паркинсона как пример мультифакториального заболевания. Фармкогеномика и противоопухолевая терапия. Транскриптомные исследования в изучении мультифакториальных заболеваний и в анализе механизма действия лекарственных средств. Этногеномика. Изменение генофонда народонаселения во времени и пространстве.
Молекулярная диагностика (2 часа)
Преподаватель: к.м.н. В.В. Демкин
Технологии, основанные на индикации нуклеиновых кислот: методы амплификации нуклеиновых кислот, компоненты и условия проведения полимеразой цепной реакции, методы анализа продуктов амплификации, микрочипы. Примеры решения конкретных диагностических задач. Технологии, основанные на индикации белков и других биомолекул. Иммуноферментый анализ.
Изотопно-меченные биологически активные соединения и биотехнология (2 часа)
Преподаватель: д.х.н. Г.В. Сидоров
Основные преимущества метода меченых атомов перед традиционными химическими и физико-химическими методами детектирования. Биогенные элементы (азот, кислород, водород, углерод, сера, фосфор), их изотопы. Наиболее распространенные изотопы для получения меченых биологически важных соединений, их основные характеристики. Основные методы синтеза изотопно-меченых соединений и используемое для этого исходное изотопное сырье. Радиоактивные изотопы и основные характеристики меченого соединения (молярная радиоактивность, химическая и радиохимическая чистота). Соединения, меченные углеродом-14 и тритием. Соединения, меченные тритием и основные способы их синтеза. Анализ и устойчивость изотопно-меченых соединений. Особенности работы с радиоактивно меченными соединениями, их радиометрия, дозиметрия и основные меры безопасности.
Антибиотики и устойчивость к ним бактерий (2 часа)
Преподаватель: д.б.н. М.А. Петрова
Антибиотики: понятие, история создания, классификация. Основные механизмы устойчивости к антибиотикам: энзиматическая инактивация (ацетилирование, фосфорилирование, гидролиз и др.), модификация молекулы-мишени, ограничение доступа антибиотика к мишени (выведение антибиотика из микробной клетки, нарушение проницаемости внешних мембран микробной клетки), образование метаболического «шунта». Роль горизонтального переноса генов и спонтанных мутаций в возникновении штаммов, обладающих множественной лекарственной устойчивостью.
Фабрики репликации ДНК бактерий
• Инициация репликации ДНК представляет собой ключевую контрольную точку клеточного цикла бактерий
• Репликация происходит в двух направлениях, начиная от фиксированной точки, называемой oriC
• Процесс реликации организован в специальные «фабрики»
• Белки рестарта репликации обеспечивают прохождение вилок от точки начала до точки терминации
• На кольцевых хромосомах обычно присутствует ловушка, обеспечивающая схождение репликативных вилок в точке терминации
• Для кольцевых хромосом необходимы специальные механизмы, координирующие терминацию репликации с декатенацией, разделением димера хромосомы, сегрегацией и клеточным делением
• Белок SpoIIIE завершает процесс сегрегации хромосомы, транспортируя захваченные участки ДНК из области, ограниченной септой деления
Репликация хромосомы у бактерий начинается в некоторой фиксированной точке, называемой oriС (точке начала репликации) и имеет двунаправленный характер, т. е. репликативные вилки одновременно движутся по часовой стрелке и в противоположном направлении. Процесс репликации характеризуется высокой процессивностью, и репликативные вилки встречаются в точке, расположенной диаметрально противоположно oriC, которая называется terC (точка окончания репликации).
Репликация происходит в дискретных сайтах, называемых фабриками репликации, в которых накапливаются субъединицы аппарата репликации. Исследование функционирования фабрик репликации в клеточном цикле B. subtilis показало, что в одной фабрике происходит один раунд репликации. Таким образом, образование примерно одного участка меченого белка репликации соответствует одному раунду репликации ДНК. Эти данные противоречат традиционной точке зрения о том, что репликативные вилки отходят от точки начала, oriC, в двух противоположных направлениях. Вместо этого предполагается, что обе вилки расположены рядом. При такой локализации возможно, что обе вилки могут иметь общий пул субъединиц белков репликации, и даже общий пул предшественников ДНК — нуклеотидтрифосатов.
С учетом такой локализации репликативных вилок, предложена модель общей организации репликации ДНК в клетке. На начальном этапе репликации хромосомы репликативная фабрика связывает область, содержащую oriC. С противоположных сторон репликативного комплекса появляются новореплицированные копии oriC, и обе репликативные вилки располагаются вместе. По мере прохождения репликации, нереплицированный участок хромосомы поступает на фабрику репликации в середине клетки, а новореплицированная ДНК выходит к противоположным полюсам. Области, содержащие oriС, сегрегируют у противоположных полюсов клетки. Область terC локализуется в середине клетки, в плотной массе нереплицированной ДНК. В конце концов происходит репликация области, содержащей terC, завершается раунд репликации хромосомы, и фабрика репликации прекращает свое существование.
Флуоресцентная микрофотография, показывающие положение фабрик репликации в делящихся клетках.
Клетки экспрессируют конъюгат зеленого флуоресцирующего белка и DnaE, субъединицы холофермента ДНК-полимеразы.
При данных условиях роста в каждой клетке находятся два реплицирующихся нуклеоида,
в каждом из которых находится центральная фабрика репликации.
Модель, описывающая функционирование фабрики репликации, помогает объяснить удивительную процессивность репликации ДНК in vivo. Репликативные вилки редко проходят путь от начала репликации до ее окончания без перерывов. Остановка репликации может объясняться повреждениями в ДНК или задержкой функционирования РНК-полимеразы. В бактериальной клетки существуют много механизмов, которые позволяют возобновить движение репликативных вилок до или после репарации поврежденной цепи ДНК. Сборка всех этих белков в непрочный комплекс вместе с новосинтезированными цепями ДНК могла бы способствовать восстановлению продвижения репликативных вилок и обеспечить их надлежащее функционирование до момента завершения репликации.
Прекращение репликации ДНК выдвигает ряд топологических проблем для клеток бактерий, имеющих кольцевые хромосомы.
• Во-первых, репликативные вилки, сходящиеся в одной точке, генерируют много положительных супервитков, для снятия которых необходимы топоизомеразы.
• Во-вторых, завершенные хромосомы должны быть скручены (катенированы), что также требует участия различных топоизомераз.
• В-третьих, при рекомбинации между сестринскими хромосомами могут образоваться димеры, которые до момента сегрегации должны быть превращены в мономеры.
Эти проблемы решаются с помощью специальных белков, присутствующих в клетках бактерий.
Когда происходит задержка репликативной вилки, и она не достигает точки окончания, возникает опасность того, что вторая вилка, продвигающаяся по другой хромосоме, пройдет через эту точку и будет продолжать движение по другой половине хромосомы. Белки прекращения репликации контролируют сайт terC, блокируя прохождение вилок в неправильном направлении. В результате репликативные вилки вынуждены встретиться в определенной области хромосомы, где клетка может управлять последующими процессами разделения димера и декатенации. Декатенация, по-видимому, осуществляется при действии специальной топоизомеразы, Торо IV. В разделении димера участвует система сайт-специфической рекомбинации, которая включает гетеродимерную каталитическую резольвазу (XerCD) и сайт мишени dif локализованный в терминальном домене.
При образовании димера хромосомы происходит спаривание двух dif сайтов, и XerCD разрезает и обменивает две нити хромосом, образуя два отдельных кольца. Очевидно, что эта система нуждается в каком-то способе разграничения димерных и мономерных хромосом, и она должна действовать только на димеры (поскольку при рекомбинации мономеров образуется димер!) Пространственное разделение молекул ДНК у Е. coli, по-видимому, находится под контролем белка FtsK, который выполняет несколько функций на поздних стадиях клеточного деления. Во-первых, он необходим для образования септы деления. Во-вторых, FtsK способен транслоцировать ДНК. За счет этого, при попадании ДНК в область, ограниченную септой деления, она может выходить оттуда. При этом может достигаться более близкое расположение двух сайтов dif в димерной хромосоме, и возможно, что смысл переноса ДНК состоит в том, чтобы обеспечить такое расположение этих сайтов. В-третьих, при необходимости FtsK активирует XerCD рекомбиназу. Это приводит к разделению димера и обеспечивает распределение по клеткам новых сестринских мономерных хромосом. Наконец, FtsK взаимодействует также с топоизомеразой IV, которая принимает участие во всех прочих процессах, связанных с декатенацией хромосом.
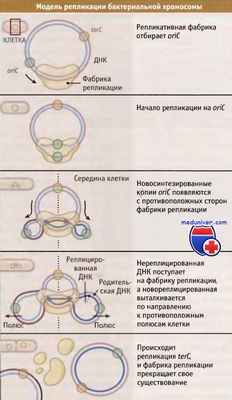
Модель репликации кольцевой хромосомы, которая учитывает близкое расположение двух репликативных вилок в фабрике репликации.
Одна цепь ДНК обозначена синим цветом; комплементарная ей цепь — красным.
Новореплицированные цепи ДНК обозначены более темным оттенком.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Фабрики репликации ДНК бактерий
ДНК-полимеразы, синтезируемые другими бактериями и многими бактериофагами, различаются по своим физической структуре и свойствам. Тем не менее, катализируемые ими реакции практически идентичны реакциям, изученным у Е. соli. У всех ДНК-полимераз есть корректирующая 3'-5'-экзонуклеаза, однако 5'-3'-экзонуклеаза у большинства ферментов отсутствует. Например, ДНК-полимераза фага Т4 может осуществлять 3'-5'-экзонуклеазную реакцию и корректировать ошибки репликации, но не способна катализировать 5'– З'-экзонуклеазную реакцию и поэтому не может обеспечить ник-трансляцию. При репликации ДНК фага Т4 5'-3'-экзонуклеазную реакцию удаления РНК-праймеров перед объединением фрагментов Оказаки катализирует другой кодируемый фагом белок. В процессе прерывистого синтеза отстающих нитей и репарации повреждений ДНК фага Т4 этот фермент работает согласованно с фаговой ДНК-полимеразой. Некоторые вирусы животных (например, герпесвирус, вирус коровьей оспы и вирус гепатита) индуцируют синтез особых полимераз для репликации своих геномов.
Другие вирусы образуют белки, которые стимулируют системы репликации клеточной ДНК или участвуют в репликации вирусной ДНК. Например, паповавирусы синтезируют белки, необходимые для инициации репликации. Аденовирусы человека кодируют белки, «запускающие» инициацию синтеза обеих цепей линейной вирусной ДНК. Они продуцируют также особые ДНК-связывающие белки, облегчающие репликацию.
4.3.1.2. ДНК-полимеразы эукариот
В эукариотических клетках идентифицировано множество ДНК-полимераз, но их физические и функциональные свойства изучены менее детально, чем у соответствующих ферментов прокариот.
Таблица 3
ДНК-полимеразы эукариот
Точная пространственная структура, определенная с помощью рентгеноструктурного анализа, известна лишь для одной ДНК-полимеразы эукариот – полимеразы β-типа, которая заметно отличается по строению от других эукариотических ДНК-полимераз. Сводные данные об основных полимеразах эукариот приведены в таблице 3.
4.3.1.3. ДНК-полимераза α – праймаза
В клетках эукариот синтез ДНК происходит, в основном, в специфических плотных структурах ("репликативных фабриках"), присоединенных к диффузному ядерному матриксу. Предполагается, что в молекулярной организации ядерного матрикса играют некоторую роль фосфолипиды и что ДНК связана с ядерным скелетом гидрофобными взаимодействиями. "Репликативные фабрики" или 21S репликативные комплексы, включают в себя группу ферментов, состоящую не менее чем из 30 белков с молекулярной массой от 15 до 300 кДа, и содержат помимо ДНК-полимеразы α – праймазы еще и 3'-5'-экзонуклеазу, ДНК-лигазу I, РНКазу Н, ДНК-топоизомеразу I, ДНК-геликазу, РСNA и ряд других факторов. Также этот комплекс содержит RРА, специфически взаимодействующий с субъединицей р48 комплекса полимераза-праймаза. Значительный запас ДНК-полимеразы α накапливается в яйцеклетках иглокожих, амфибий, костистых рыб и дрозофилы для обеспечения интенсивной репликации ядерной ДНК в ходе раннего развития.
Как правило, ДНК-полимеразы α не обладают корректорской 3'—5'-экзонуклеазной активностью. Однако в каталитической субъединице 182 кДа ДНК-полимеразы α дрозофилы обнаружена 3'—5'-экзонуклеаза, проявляющая активность только при диссоциации субъединицы 73 кДа.
Мультибелковая форма ДНК-полимеразы α содержит также белок, который связывает динуклеотид диаденозинтетрафосфат (Ар4А). Предполагается, что Ар4А участвует в репликации ДНК и клеточном делении. Имеются данные о способности ДНК-полимеразы α использовать Ар4А в качестве праймера. Однако участие Ар4А в качестве праймера in vivo маловероятно, скорее он используется как эффектор. Интересно, что триптофанил-тРНК-синтетаза, синтезирующая этот динуклеотид, находится в том же мултибелковом комплексе.
Обычно комплекс праймаза-полимераза α состоит из четырех субъединиц: большой субъединицы с молекулярной массой 180 кДа, или семейства полипептидов с размерами от 140 до 160 кДа; субъединицы с молекулярной массой около 68–70 кДа и двух малых субъединиц с молекулярными массами 54–58 и 46–50 кДа. Субъединица р180 отвечает за полимеразную функцию. С двумя малыми субъединицами связана праймазная активность. При этом субъединица 48 кДа является каталитической и непосредственно осуществляет праймирование ДНК, а субъединица 58 кДа участвует в связывании инициирующего пуринового нуклеотида и присоединении субъединицы р48 к ДНК-полимеразе α. Она также влияет на скорость полимеризации и стабильность продукта, синтезируемого субъединицей 48 кДа. р58 также облегчает проникновение р48 из цитоплазмы в ядро. Субъединица р180 непосредственно взаимодействует с р58.
С каталитической субъединицей связана субъединица 68–70 кДа, которая необходима для транспорта каталитического полипептида в клеточное ядро. Субъединица 68–70 кДа участвует также в регуляции уровня ДНК-полимеразы α в клетке, она стимулирует синтез каталитического полипептида. Хотя комплекс ДНК-полимераза α-праймаза состоит из четырех субъединиц, количественный состав этого комплекса может быть различным. Вероятно, полимераза α и праймаза находятся в «коре» в соотношении 1: 3.
4.3.1.4. Реакция праймирования
Инициация репликации и прерывистый синтез ДНК на отстающей цепи происходит по РНК-праймерному механизму и является универсальным свойством репликации ДНК у про– и эукариот.
ДНК-праймаза отличается от других РНК-полимераз целым рядом присущих только ей свойств. Во-первых, матричной и субстратной специфичностью. Во-вторых, необычной процессивностью – синтезом мультимеров, кратных 10-нуклеотидным звеньям. В-третьих, низкой точностью и устойчивостью к некоторым ингибиторам РНК– и ДНК-полимераз. Здесь необходимо вернуться к проблеме синтеза РНК-праймеров. Синтез РНК-праймеров на природных матрицах начинается во множественных, но не случайных участках, его инициация происходит с АТР или GТР даже при высоких концентрациях СТР и UTР. Показано, что, например, ДНК вируса SV40 содержат предпочтительные участки инициации – 3'-dСТТТ или 3'-dССС, расположенные внутри участков из 7-25 пиримидиновых нуклеотидов. Высокое соотношение АТР/GТР повышает вероятность инициации в участках 3'-dСТТТ, а низкое – в участках 3'-dССС. Таким образом, концентрация NТР и нуклеотидная последовательность матрицы определяют участки инициации. Впрочем, участки инициации in vivo заметно отличаются от участков инициации, используемых во время репликации ДНК SV40. Во многих случаях обнаружена последовательность 5'-YYYYYYYYСТТТYYYY-3', где Y = С или Т, которая является стартовой площадкой для инициации синтеза ДНК-праймазой в составе комплекса с ДНК-полимеразой α. Минимальная длина пиримидинового кластера должна быть не менее 7 н. Замена одного из пиримидинов на 3'-конце кластера значительно понижает частоту инициации, а замены внутри и вне кластера приводят к смещению точки старта. Известно, что матрицу распознает сама ДНК-праймаза. Стартовый нуклеотид вновь синтезированного праймера всегда является пурином (чаще всего – аденином).
Этот комплекс имеет еще одну специфическую функцию – синтез теломерной отстающей цепи ДНК млекопитающих осуществляет ДНК-полимераза α-праймаза.
ДНК-праймаза – сравнительно медленный фермент. Средняя скорость включения NTP этим ферментом примерно на два порядка меньше, чем скорость включения dNТР ДНК-полимеразой α. ДНК-полимераза α способна удлинять праймеры длиной более 7-10 н. Продукты длиной 2–6 н. не являются субстратами ДНК-полимеразы α и называются абортивными, До синтеза РНК-праймера ДНК-полимераза α и ДНК-праймаза действуют независимо, а после этого их активности координируются. Синтезированный РНК-праймер перемещается в полимеразный активный центр без диссоциации в раствор. Это внутримолекулярное перемещение праймера в дуплексе с матрицей является быстрым и сравнимо по скорости с синтезом праймера. После того как ДНК-полимераза α удлинит праймер, праймаза начинает синтез нового праймера, и цикл повторяется. ДНК-праймаза эукариот отличается от других РНК-полимераз своей способностью к включению дезоксирибонуклеотида в праймер, таким же свойством обладает и праймаза прокариот. Одним из возможных объяснений необходимости включения dNТР в 3'-конец праймера является необходимость перехода от А-формы к В-форме ДНК. Поэтому понимание механизма "переключения" комплекса ДНК-полимеразы α-праймазы от синтеза РНК к ДНК имеет очень большое значение. Способность праймазы узнавать одновременно и рибо-, и дезоксириботрифосфаты представляет серьезный научный интерес.
Выбор РНК-праймера определяется гидрофобным характером белково-нуклеиновых взаимодействий. В случае гибрида РНК-ДНК дуплекс находится в А-форме, в которой обеспечивается оптимальный баланс между гидрофобными и комплементарными взаимодействиями оснований матрицы и праймера.
После инициации рост праймера сопровождается извлечением оснований матрицы из гидрофобной полости белка для спаривания с основаниями растущей цепи РНК. В условиях такой конкуренции короткие ди– и тринуклеотиды легко диссоциируют, образуя абортивные продукты. С ростом длины праймера прочность дуплекса увеличивается, ослабевает влияние гидрофобности активного центра, и пары оснований приближаются к оси спирали. При длине праймера 7 н создаются условия для включения дезоксинуклеотида и перехода в энергетически более выгодную В-форму. Здесь нужно учитывать и большее сродство к ферменту dNТР по сравнению с NТР. Предложенная концепция, по-видимому, носит универсальный характер, поскольку подобное происходит и при инициации транскрипции.
ДНК-полимераза α связывает сначала матрицу, затем праймер и субстрат. ДНК-полимераза α наиболее активна на двунитевой ДНК, содержащей бреши длиной не менее 20–30 н. Область связывания матрицы с ДНК-полимеразой α является достаточно протяженной. Она строго защищает от гидролиза 9 н праймерной цепи, 13 н двухцепочечного участка и 14 н одноцепочечной матрицы и слабо защищает несколько оснований вне этого района. Фермент связывается с 19–20 н матрицы посредством ионных и гидрофобных взаимодействий, эффективность связывания при этом коррелирует с гидрофобностью оснований матрицы.
Отличительной особенностью ДНК-полимеразы α является ее способность удлинять РНК-праймеры и прочная ассоциация с ДНК-праймазой, которая синтезирует эти праймеры.
Средняя процессивность ДНК-полимеразы α составляет 20–50 н. Праймаза редко ошибается в момент синтеза динуклеотида, но затем легко использует неправильные NТР. Хотя скорость включения ошибочных нуклеотидов зависит от нуклеотидной последовательности матрицы и самого неправильного нуклеотида, в принципе, ДНК-праймаза является самым неточным нуклеотид-полимеризующим ферментом. В некоторых случаях один неправильный нуклеотид приходится менее чем на 100 правильных. Встраивание неправильного нуклеотида не препятствует включению следующего правильного нуклеотида. Праймаза может полимеризовать сходные нуклеотиды и генерировать праймеры с множественными ошибками, котрые не ингибируют дальнейший синтез и после внутримолекулярного переноса в ДНК-полимеразный активный центр удлиняются ДНК-полимеразой в присутствии dNTP.
Существуют две модели механизма синтеза праймеров, некомплементарных матрице. Согласно первой модели, фермент просто включает некомплементарные матрице нуклеотиды. Вторая модель предполагает включение нуклеотида, комплементарного матрице, с последующим скольжением праймера относительно матрицы. Низкая точность ДНК-праймазы послужила основой гипотезы о том, почему именно РНК является затравкой при репликации ДНК. Поскольку первые нуклеотиды могут ошибочно включаться в новую растущую цепь, предполагается, что РНК-праймер отмечает "высокоошибочный" участок для последующего вырезания и более точной застройки. Эта точка зрения выглядит убедительной, но, вероятно, главная причина появления РНК-праймера связана все-таки с более эффективной инициацией синтеза ДНК при наличии А-формы РНК-ДНК дуплекса.
От атомов к дереву: вирусы и клетки
Латинское слово virus вообще-то значит «яд». Например, змеиный яд и яд, которым смазывают наконечник отравленной стрелы, обозначаются в классической латыни именно этим словом. Ну а в конце XIX века биологи назвали вирусами болезнетворные агенты, обладающие тремя очень характерными особенностями:
- они невидимы под световым микроскопом;
- они свободно проходят сквозь фильтры, предназначенные для задержки бактерий (отсюда устойчивое, хотя и совсем уж сейчас устаревшее выражение «фильтрующийся вирус»);
- они не поддаются выращиванию ни на каких искусственных питательных средах.
Не приходится удивляться, что при таких вводных природа вирусов первое время была совершенно загадочна. Вирус мог оказаться мельчайшим живым организмом, а мог и просто ядовитой молекулой. Лет 100 назад биологи вполне допускали и то и другое.
В 1935 году американский биохимик Уэнделл Стэнли ухитрился не только выделить из табачного сока вирус табачной мозаики, но и кристаллизовать его — так, как если бы это была обычная молекула. В то время это произвело сенсацию, тем более что под микроскопом вирусы на тот момент еще никто не видел. Неудивительно, что, получив этот результат, Стэнли посчитал вирус просто белковой молекулой, пусть и имеющей кое-какие особые свойства…
…Постепенно биологам становилось ясно, что вирус…— это мелкий (не больше 200 нанометров) инфекционный агент, не обладающий автономностью и не способный размножаться делением, в отличие от живых клеток… Любой вирус обязательно содержит помимо белка нуклеиновую кислоту, но только одного типа — или ДНК, или РНК. Последнее резко отличает вирусы от живых клеток, в состав которых… всегда входят оба типа нуклеиновых кислот…
В любом случае такие взгляды подразумевали, что вирусы — это по сути своей нечто более простое, чем клетки. Гораздо, качественно более простое. Иная ступень устройства природы, если угодно.

Мимивирус
Вот это мнение и оказалось неверным. Скажем аккуратнее: не вполне верным. Потому что в 2003 году был открыт мимивирус… — гигантский вирус, который паразитирует в пресноводных амебах и создает в них «вирусные фабрики». Его вирион приближается к невероятному для вирусов размеру в 500 нанометров, то есть 0,5 микрона. Этот размер больше, чем у некоторых бактерий (среди последних есть внутриклеточные паразиты со средним размером 200–300 нанометров). Такой вирус вполне можно увидеть в световой микроскоп, а не только в электронный. И бактериальные фильтры его задерживают. Собственно говоря, поначалу мимивирусы за бактерий и приняли. Неудивительно, учитывая, что средний размер обычных вирусов — всего-то около 100 нанометров.
Все гигантские вирусы оказались ДНК-содержащими, причем ДНК у них двуцепочечная. Прочтение генома мимивируса показало, что этот геном состоит из 1,18 миллиона пар нуклеотидов и включает 979 генов, кодирующих белки… Он приближается по величине к геномам клеточных организмов — и не просто приближается, а вполне достигает их размеров…
Но ведь и мимивирус не уникален. Исследования быстро показали, что гигантских ДНК-содержащих вирусов в природе не так уж мало.
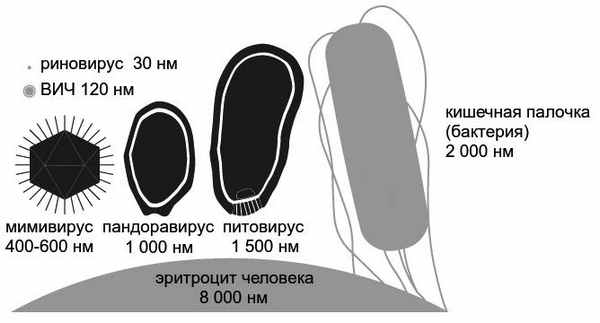
Форма и размеры некоторых гигантских ДНК-содержащих вирусов в сравнении с «обычными» вирусами и с клетками
За прошедшие годы было открыто еще несколько близких друг к другу родов гигантских ДНК-содержащих вирусов. Они получили разнообразные названия: марсельвирусы, мамавирусы, мегавирусы, пандоравирусы, молливирусы, питовирусы и даже моумоувирусы…
В 2017 году был описан… клоснойвирус, названный по имени города Клостернойбург в Нижней Австрии… Его геном содержит необычайно большой (для вируса) набор генов, служащих для трансляции: 25 генов транспортных РНК и 19 генов ферментов-кодаз, которые «пришивают» к транспортным РНК транспортируемые ими аминокислоты. Судя по всему, вирус получил эти гены из геномов своих хозяев — одноклеточных эукариот ; в мире вирусов подобные захваты чужих генов вообще не редкость. Но вот уж чего-чего, а трансляции у вирусов быть не должно.
Клоснойвирус
Впрочем, полного аппарата трансляции, способного к самостоятельной работе, нет и у клоснойвируса. У него полностью отсутствуют гены, кодирующие рибосомную РНК и рибосомные белки… — это, как мы уже говорили, одно из самых главных различий между вирусами и клетками. По этому признаку никаких переходных форм между ними пока не видно.
Так или иначе, открытие гигантских вирусов смело можно назвать одним из важнейших в современной биологии. Оно целиком датируется XXI веком. Открытие гигантских вирусов заметно изменило наши представления о структуре живой природы в целом. Конечно, никто не утверждает, что пандоравирус устроен сложнее кита-полосатика. Верхний предел сложности (даже чисто генетической) у клеточных организмов явно гораздо выше; это видно хотя бы по размерам их геномов. Но вот сказать, что любая клетка устроена сложнее любого вируса, теперь нельзя никак.
Итак, главный вывод: вирусы и клеточные организмы существенно перекрываются друг с другом как по размеру, так и по сложности. Это не две эволюционные ступени, а две ветви…
…Бросается в глаза, что есть как минимум две важные группы генов, свойственных клеткам, но не свойственных вирусам. Это гены энергетического обмена и гены, ответственные за создание рибосом. У подавляющего большинства вирусов никаких генов энергетического обмена нет вообще, а если даже они есть, то кодируют только отдельные ферменты, но не полную систему синтеза АТФ … Судя по биоинформатическим данным, последний общий предок всех клеточных организмов имел как минимум 34 рибосомных белка, и эти белки (вместе с кодирующими их генами) сохранились у всех бактерий, архей и эукариот. У вирусов же нет ни одного из них.
Бактерий, архей и эукариот можно с полным основанием назвать рибосомокодирующими организмами (ribosome-encoding organisms, REO). Этот признак четко отделяет всех их, вместе взятых, от вирусов.
…Обладают ли вирусы чем-то таким, что ни у каких клеток не встречается? Да. В геноме любого вируса, заслуживающего этого названия, есть гены, кодирующие белки капсида — белкового «футляра», заключающего в себе ДНК или РНК. Слово «футляр» тут, пожалуй, вводит в заблуждение, на самом деле вирусные капсиды частенько бывают довольно сложными структурами, построенными из разнородных частей и способными изменять свою геометрию (как, например, капсид бактериофага, который будто бы шприцем впрыскивает вирусный генетический материал в клетку бактерии). Вирус без капсида — это не вирус, а cубвирусная частица. Итак, особенность, которая есть у всех вирусов, и только у них, — это экспрессия генов капсида.
Вирусы можно с полным основанием назвать капсид-кодирующими организмами (capsid-encoding organisms, CEO). Этот признак четко отделяет их от всех, кто состоит из клеток.
…Деление организмов на рибосомокодирующие и капсид-кодирующие — это первичная дихотомия, на которой может основываться вся система живой природы. Она соответствует двум огромным эволюционным ветвям, разошедшимся на заре жизни, — двум ветвям, каждая из которых породила свой собственный биологический мир. Одни организмы «изобрели» рибосому и получили автономный аппарат трансляции — эффективный, но требующий постоянного снабжения энергией. А другие организмы сделали ставку на манипулятивное использование в своих интересах чужих аппаратов трансляции и на очень быстрое размножение. Сложностью при этом пришлось по большей части пожертвовать, но зато выживание генетического материала, рассеянного в чуждой и часто агрессивной среде, потребовало «изобретения» капсида.
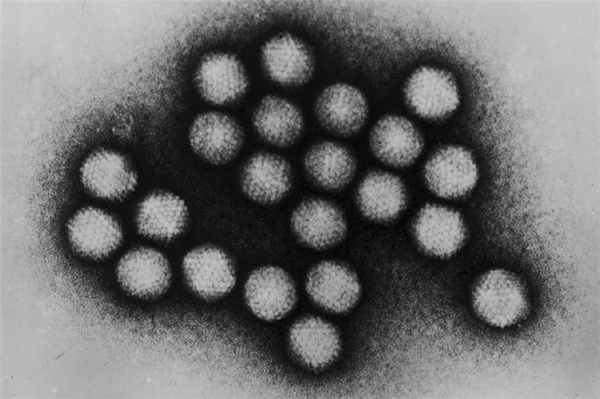
Икосаэдрические капсиды аденовируса
Более того, не исключено, что заодно некоторые древние вирусы «изобрели» и ДНК. Ведь приспособительный смысл особенностей ДНК, отличающих ее от РНК, в том, что она чисто химически более устойчива. А в условиях, когда вирионы переносились на большие расстояния водой или ветром, это могло быть особенно важно для сохранности их генетического материала. Между тем, есть независимо обоснованная гипотеза, что первые клетки были РНК-содержащими — аналогично тому, как бывают РНК-содержащие вирусы. Это хорошо согласуется с тем фактом, что рибосомы со всем прилагающимся к ним набором РНК и белков, появились раньше, чем клеточные механизмы репликации ДНК (во всяком случае, к таким выводам приводят данные сравнительной геномики). А если первые ДНК-содержащие вирусы уже существовали в эпоху первых клеток, то. Почему бы этим клеткам не получить «ноу-хау» хранения генетической информации на ДНК прямо от вирусов? Такой обмен генами вполне возможен. Тогда получается, что взаимодействие (или даже слияние) ДНК-содержащего вируса и РНК-содержащей клетки произвело на свет ДНК-содержащую клетку — ту самую форму жизни, которая сейчас доминирует на Земле.
Из чего состоят живые тела и при чем тут углерод? Что такое генетический код, кто такие вирусы, как устроено эволюционное древо и почему произошел кембрийский взрыв? Книга Сергея Ястребова «От атомов к дереву. Введение в современную биологию» дает актуальные ответы на эти и многие другие вопросы. «Фокусом» рассказа служит эволюция жизни на Земле: автор считает, что только под этим углом зрения самые разные биологические проблемы обретают единый смысл. Книга состоит из четырех частей, темы которых последовательно расширяются: «Химия жизни», «Механизм жизни», «Древо жизни» и «История жизни».
Издательство: «Альпина нон-фикшн»
На сайте могут быть использованы материалы интернет-ресурсов Facebook и Instagram, владельцем которых является компания Meta Platforms Inc., запрещённая на территории Российской Федерации
Читайте также: