Функции цитоплазматической мембраны бактерий (прокариот)
Добавил пользователь Alex Обновлено: 29.01.2026
Структура Какой состав имеет цитоплазматическая мембрана? Строение пленки достаточно разнообразно. В соответствии с химической организацией, она представляет собой комплекс белков и липидов. Цитоплазматическая мембрана клетки включает в себя бислой. Он выступает в качестве основы. Кроме этого, цитоплазматическая мембрана содержит холестерол и гликолипиды. Этим веществам свойственна амфипатричность. Другими словами, в них присутствуют гидрофобные ("боящиеся влаги") и гидрофильные ("любящие воду") концы. Последние (фосфатная группа) направлены наружу от мембраны, вторые (остатки от жирных кислот) ориентированы друг к другу. За счет этого и формируется липидный биполярный слой. Липидные молекулы обладают подвижностью. Они способны перемещаться в собственном монослое либо (что редко) из одного в другой.
Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды – основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Функции Барьерная. Защитная пленка обеспечивает активный, пассивный, избирательный, регулируемый обмен соединений с внешней средой. За счет избирательной проницаемости осуществляется отделение клетки и ее компартментов и снабжение их нужными веществами.
Транспортная. Сквозь пленку осуществляется переход соединений от клетки к клетке. Благодаря этому доставляются питательные соединения, удаляются конечные продукты обмена, происходит секреция разных веществ. Кроме этого, формируются ионные градиенты, на оптимальном уровне поддерживаются ионная концентрация и рН. Они необходимы для активной деятельности ферментов клетки.
Вспомогательные задачи
Матричная. Эта функция обеспечивает определенную ориентацию и взаиморасположение белков мембраны, а также оптимальное их взаимодействие. Механическая. За счет нее обеспечивается автономность клетки, внутренних структур. Также осуществляется соединение элемента с прочими аналогичными. Энергетическая. На фоне фотосинтеза в хлоропластах и при осуществлении клеточного дыхания в мембранах активны системы энергетического переноса. В них также участвуют и белковые соединения.
Рецепторная. Ряд белков, которые присутствуют в мембране, обеспечивает восприятие различных сигналов. К примеру, циркулирующие в крови стероиды оказывают воздействие только на те клетки-мишени, которые обладают соответствующими гормонам рецепторами. Химические соединения, обеспечивающие проведение импульсов (нейромедиаторы), также связываются с помощью особых белков клеток-мишеней.
Дополнительные сведения
Если какие-то частицы по тем или другим причинам не способны пройти сквозь фосфолипидный бислой (к примеру, вследствие гидрофильных свойств, поскольку внутри цитоплазматическая мембрана гидрофобна и такие соединения не пропускает, либо из-за больших размеров самих частиц), но они необходимы, то пройти они могут с помощью специальных белков-переносчиков (транспортеров) и белков-каналов. Либо проникновение их осуществляется посредством эндоцитоза. В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
Цитоплазматическая клеточная мембрана состоит из трех слоев:
Среднего – бимолекулярного слоя липидов;
Толщина мембраны 7,5-10 нм. Бимолекулярный слой липидов является матриксом мембраны. Липидные молекулы его обоих слоев взаимодействуют с белковыми молекулами, погруженными в них. От 60 до 75% липидов мембраны составляют фосфолипиды, 15-30% холестерин. Белки представлены в основном гликопротеинами. Различают интегральные белки, пронизывающие всю мембрану, ипериферические, находящиеся на наружной или внутренней поверхности.
Интегральные белки образуют ионные каналы, обеспечивающие обмен определенных ионов между вне- и внутриклеточной жидкостью. Они также являются ферментами, осуществляющими противоградиентный перенос ионов через мембрану.
Периферическими белками являются хеморецепторы наружной поверхности мембраны, которые могут взаимодействовать с различными физиологически активными веществами.
Функции мембран: 1. Обеспечивает целостность клетки как структурной единицы ткани. 2.Осуществляет обмен ионов между цитоплазмой и внеклеточной жидкостью. 3.Обеспечивает активный транспорт ионов и других веществ в клетку и из нее. 4.Производит восприятие и переработку информации, поступающей к клетке в виде химических и электрических сигналов.
___________________________ Цитоплазматическая мембрана составляет в зависимости от вида бактерий 8–15 % сухой массы клетки. Химический состав ее представлен белково-липидным комплексом, в котором на долю белков приходится 50–75 %, на долю липидов – 15–50 %. Главным липидным компонентом мембраны являются фосфолипиды. Белковая фракция цитоплазматической мембраны представлена структурными белками, обладающими ферментативной активностью. Белковый состав цитоплазматической мембраны разнообразен. Цитоплазматическая мембрана бактерий по химическому составу в целом сходна с мембранами эукариотических клеток, но мембраны бактерий богаче белками, содержат необычные жирные кислоты и в основном не имеют стеринов. К строению цитоплазматической мембраны бактерий приложима жидкостно-мозаичная модель, разработанная для мембран эукариот. Согласно этой модели, мембрана состоит из бислоя липидов. Гидрофобные «концы» молекул фосфолипидов и триглицеридов направлены внутрь, а гидрофильные «головки» – наружу. В двойной слой липидов встроены
белковые молекулы . По расположению и характеру взаимодействия с липидным бислоем белки цитоплазматической мембраны подразделяются на периферические и интегральные.
Цитоплазматическая мембрана выполняет ряд существенных для клетки функций:
• поддержание внутреннего постоянства цитоплазмы клетки. Это достигается за счет уникального свойства цитоплазматической мембраны – ее полупроницаемости. Она проницаема для воды и низкомолекулярных веществ, но не проницаема для ионизированных соединений.Транспорт таких веществ внутрь клетки и выход наружу осуществляется за счет специализированных транспортных систем, которые локализуются в мембране. Такие транспортные системы функционируют за счет механизмов активного транспорта и системы специфических ферментов пермеаз;
• с вышеуказанной особенностью (полупроницаемостью) цитоплазматической мембраны связана и функция транспорта веществ в клетку и вывод их наружу;
• в цитоплазматической мембране локализуются электронтранспортная цепь и ферменты окислительного фосфорилирования;
• цитоплазматическая мембрана связана с синтезом клеточной стенки и капсулы за счет наличия в ней специфических переносчиков для образующих их молекул;
• в цитоплазматической мембране закреплены жгутики. Энергетическое обеспечение работы жгутиков связано с цитоплазматической мембраной.
У прокариот, принадлежащих к разным таксономическим группам, обнаружены мезосомы, которые образуются при впячивании цитоплазматической мембраны в цитоплазму. Существуют разные точки зрения относительно роли мезосом в бактериальной клетке. Согласно одной из них, мезосомы служат для усиления мембранзависимых функциональных активностей клетки, так как в мембранах, образующих мезосомы, находятся ферменты, участвующие в энергетическом метаболизме бактерий. Кроме того, считают, что мезосомы играют роль в репликации ДНК и последующем расхождении ее копий по дочерним клеткам. Мезосомы участвуют в процессе инициациии формирования поперечной перегородки при клеточном делении.
_______________________________________________________ Цитоплазматическая мембрана (плазмалемма) — основная, универсальная для всех клеток часть поверхностного аппарата. Ее толщина составляет около 10 нм. Плазмалемма ограничивает цитоплазму и защищает ее от внешних воздействий, принимает участие в процессах обмена веществ между клеткой и внеклеточной средой.
Основными компонентами мембраны являются липиды и белки. Липиды составляют около 40 % массы мембран. Среди них преобладают фосфолипиды.
Молекулы фосфолипидов располагаются в виде двойного слоя (липидный бислой). Как вы уже знаете, каждая молекула фосфолипида образована полярной гидрофильной головкой и неполярными гидрофобными хвостами. В цитоплазматической мембране гидрофильные головки обращены к наружной и внутренней сторонам мембраны, а гидрофобные хвосты — внутрь мембраны (рис. 30).
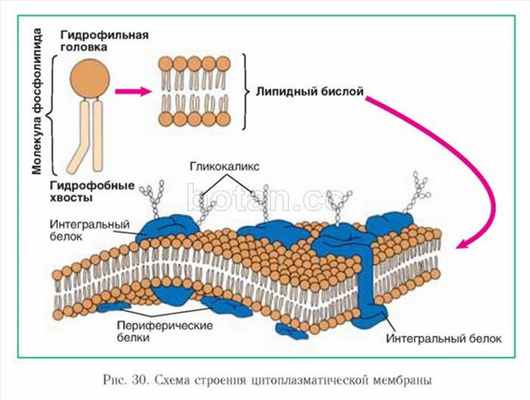
Кроме липидов, в состав мембран входят белки двух типов: интегральные и периферические. Интегральные белки более или менее глубоко погружены в мембрану либо пронизывают ее насквозь. Периферические белки располагаются на внешней и внутренней поверхностях мембраны, причем многие из них обеспечивают взаимодействие плазмалеммы с надмембранными и внутриклеточными структурами.
На внешней поверхности цитоплазматической мембраны могут располагаться молекулы олиго- и полисахаридов. Они ковалентно связываются с мембранными липидами и белками, образуя гликолипиды и гликопротеины. В клетках животных такой углеводный слой покрывает всю поверхность плазмалеммы, образуя надмембранный комплекс. Он называется гликокаликсом (от лат. гликис — сладкий, калюм — толстая кожа).
Функции цитоплазматической мембраны. Плазмалемма выполняет ряд функций, важнейшими из которых являются барьерная, рецепторная и транспортная.
Барьерная функция. Цитоплазматическая мембрана окружает клетку со всех сторон, играя роль барьера — преграды между сложно организованным внутриклеточным содержимым и внеклеточной средой. Барьерную функцию обеспечивает, прежде всего, липидный бислой, не позволяющий содержимому клетки растекаться и препятствующий проникновению в клетку чужеродных веществ.
Рецепторная функция. В цитоплазматическую мембрану встроены белки, способные в ответ на действие различных факторов внешней среды изменять свою пространственную структуру и таким образом передавать сигналы внутрь клетки. Следовательно, цитоплазматическая мембрана обеспечивает раздражимость клеток (способность воспринимать раздражители и определенным образом реагировать на них), осуществляя обмен информацией между клеткой и окружающей средой.
Некоторые рецепторные белки цитоплазматической мембраны способны распознавать определенные вещества и специфически связываться с ними. Такие белки могут участвовать в отборе необходимых молекул, поступающих в клетки.
К рецепторным белкам относятся, например, антигенраспознающие рецепторы лимфоцитов, рецепторы гормонов и нейромедиаторов и т. д. В осуществлении рецепторной функции, кроме мембранных белков, важную роль играют элементы гликокаликса.
Разнообразие и специфичность наборов рецепторов на поверхности клеток приводит к созданию сложной системы маркеров, позволяющих отличать s.свои:/ клетки (той же особи или того же вида) от s.чужих:/. Благодаря этому клетки могут вступать друг с другом во взаимодействия (например, конъюгация у бактерий, образование тканей у животных).
В цитоплазматической мембране могут быть локализованы специфические рецепторы, реагирующие на различные физические факторы. Например, в плазмалемме светочувствительных клеток животных расположена специальная фоторецепторная система, ключевую роль в функционировании которой играет зрительный пигмент родопсин. С помощью фоторецепторов световой сигнал превращается в химический, что, в свою очередь, приводит к возникновению нервного импульса.
Транспортная функция. Одной из основных функций плазмалеммы является обеспечение транспорта веществ как в клетку, так и из нее во внеклеточную среду. Выделяют несколько основных способов транспорта веществ через цитоплазматическую мембрану: простая диффузия, облегченная диффузия, активный транспорт и транспорт в мембранной упаковке (рис. 31).
При простой диффузии наблюдается самопроизвольное перемещение веществ через мембрану из области, где концентрация этих веществ выше, в область, где их концентрация ниже. Путем простой диффузии через плазмалем-му могут проходить небольшие молекулы (например, Н20, 02, С02, мочевина) и ионы. Как правило, неполярные вещества транспортируются непосредственно через липидный бислой, а полярные молекулы и ионы — через каналы, образованные специальными мембранными белками. Простая диффузия происходит относительно медленно. Для ускорения диффузного транспорта существуют мембранные белки-переносчики. Они избирательно связываются с тем или иным ионом или молекулой и переносят их через мембрану. Такой тип транспорта называется облегченной диффузией. Скорость переноса веществ при облегченной диффузии во много раз выше, чем при простой.
Диффузия (простая и облегченная) — разновидности пассивного транспорта. Он характеризуется тем, что вещества транспортируются через мембрану без затрат энергии и только в том направлении, где наблюдается меньшая концентрация данных веществ.
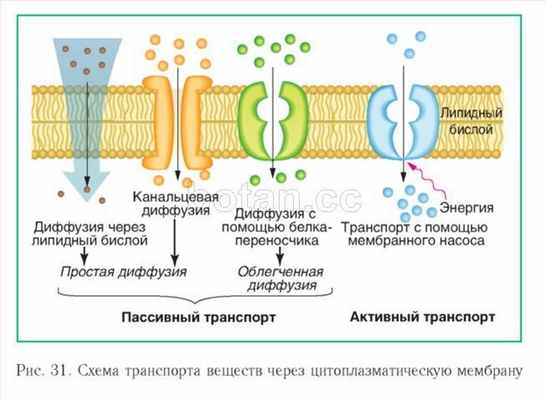
Активный транспорт — перенос веществ через мембрану из области низкой концентрации этих веществ в область более высокой. Для этого в мембране имеются специальные насосы, работающие с использованием энергии (см. рис. 31). Чаще всего для работы мембранных насосов используется энергия АТФ.
Одним из наиболее распространенных мембранных насосов является натрий-калиевая АТ Фаза (Na + /K + - АТ Фаза). Она удаляет из клетки ионы Na + и закачивает в нее ионы К + - Для работы Ыа + /К + -АТФаза использует энергию, выделяемую при гидролизе АТФ. Благодаря этому насосу поддерживается разность концентраций Na + и К + в клетке и внеклеточной среде, что лежит в основе многих биоэлектрических и транспортных процессов.
В результате активного транспорта с помощью мембранных насосов происходит также регуляция содержания Mgr + , Са 2+ и других ионов в клетке.
Путем активного транспорта через цитоплазматическую мембрану могут перемещаться не только ионы, но и моносахариды, аминокислоты, другие низкомолекулярные вещества.
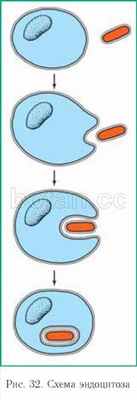
Своеобразной и относительно хорошо изученной разновидностью мембранного транспорта является транспорт в мембранной упаковке. В зависимости от того, в каком направлении переносятся вещества (в клетку или из нее), различают два вида этого транспорта — эндоцитоз и экзоцитоз.
Эндоцитоз (отгреч. эндон — внутри, китос — клетка, ячейка) — поглощение клеткой внешних частиц путем образования мембранных пузырьков. При эндоцитозе определенный участок плазмалеммы обволакивает внеклеточный материал и захватывает его, заключая в мембранную упаковку (рис. 32).
Выделяют такие разновидности эндоцитоза, как фагоцитоз (захват и поглощение твердых частиц) и пиноцитоз (поглощение жидкости).
Путем эндоцитоза осуществляется питание гетеротрофных протистов, защитные реакции организма (поглощение лейкоцитами чужеродных частиц) и др.
Экзоцитоз (от греч. экзо — снаружи) — транспортировка веществ, заключенных в мембранную упаковку, из клетки во внешнюю среду. Например, пузырек комплекса Гольджи перемещается к цитоплазматической мембране и сливается с ней, а содержимое пузырька выделяется во внеклеточную среду. Таким способом клетки выделяют пищеварительные ферменты, гормоны и другие вещества.
Биология и медицина
ЦПМ (цитоплазматическая мембрана) прокариот: функции
ЦПМ (цитоплазматическая мембрана) прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ.
Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза , в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии (дельта мю H+), используемой затем в клетке по разным каналам, в том числе и для образования химической энергии ( АТФ ). ЦПМ является одним из компонентов аппарата генерирования дельта мю H+. В мембране расположены также ферментные комплексы, обеспечивающие превращения дельта мю H+ в АТФ и обратно.
ЦПМ принимает участие в репликации и последующем разделении хромосомы прокариотной клетки.
В последнее время выявляется еще одна функциональная грань клеточных мембран - их интегрирующая роль в организме, вполне сочетающаяся с давно установленной разъединяющей (барьерной) функцией. Клетка - единое целое. В обеспечении этого принципа клеточной организации важная роль принадлежит мембранам. Показан перенос электрохимической энергии и электронов вдоль мембран. Последние рассматриваются так же как возможные пути транспорта жирорастворимых субстратов и молекулярного кислорода.
ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов. (У грамположительных форм ЦПМ является и единственным барьером такого рода, у грамотрицательных эубактерий функции дополнительного барьера (молекулярного "сита") выполняет наружная мембрана клеточной стенки, через которую молекулы транспортируются только по механизму пассивной диффузии). Осуществляется это с использованием разных механизмов мембранного транспорта.
Выделяют 4 типа транспортных систем, с участием которых происходит проникновение молекул в бактериальную клетку: пассивную диффузию, облегченную диффузию, активный транспорт и перенос химически модифицированных молекул.
Молекулы воды, некоторых газов (например, О2, Н2, N2) и углеводородов, концентрации которых во внешней среде выше, чем в клетке, проходят через ЦПМ внутрь клетки посредством пассивной диффузии. Движущей силой этого процесса служит градиент концентрации вещества по обе стороны мембраны. Основным соединением, поступающим в клетку и покидающим ее таким путем, является вода. Движение воды через мембрану, подчиняющееся законам пассивной диффузии, привело к выводу о существовании в мембране пор. Эти поры пока не удалось увидеть в электронный микроскоп, но некоторые данные о них были получены косвенными методами. Расчетным путем установлено, что поры должны быть очень мелкими и занимать небольшую часть поверхности ЦПМ. Высказывается предположение, что они не являются стабильными структурными образованиями, а возникают в результате временных перестроек молекулярной организации мембраны.
Большинство (если не все) гидрофильных веществ поступает в клетку за счет функционирования систем, в состав которых входят специальные переносчики (транслоказы, или пермеазы), так как скорость физической диффузии этих веществ через гидрофобный слой мембраны очень невелика. Переносчики - вещества белковой природы, локализованные в мембране и характеризующиеся высокой субстратной специфичностью, - связываясь с субстратом, подвергаются конформационным изменениям и вследствие этого приобретают способность к перемещению субстрата с одной стороны ЦПМ на другую.
Известен механизм транспорта, получивший название облегченной диффузии, требующий для переноса веществ через мембрану участия транслоказ. Перенос веществ в этом случае происходит по градиенту их концентрации и не требует энергетических затрат. Этот механизм транспорта не получил широкого распространения у прокариот. Основным механизмом избирательного переноса веществ через ЦПМ прокариот является активный транспорт, позволяющий "накачивать" в клетку молекулы и ионы против их концентрационных и электрических градиентов. Этот процесс, так же как и облегченная диффузия, протекает при участии локализованных в ЦПМ переносчиков белковой природы с высокой специфичностью к субстрату, но в отличие от облегченной диффузии для движения против электрохимического градиента требует затрат метаболической энергии. Транспорт такого рода должен быть поэтому сопряжен с реакциями, продуцирующими энергию в химической или электрохимической форме.
Во всех описанных выше путях переноса веществ через ЦПМ они поступают в клетку в химически неизмененном виде. У прокариот известны системы транспорта, с помощью которых осуществляется поступление в клетку ряда сахаров , при этом процесс их переноса через мембрану сопровождается химической модификацией молекул. Так происходит, например, поступление в клетки многих прокариот молекул глюкозы , в процессе которого они фосфорилируются.
Цитоплазматическая мембрана
Цитоплазматическая мембрана (ЦПМ) – обязательная структура любой клетки, расположена под клеточной стенкой бактерии и отделяющая содержимое клетки от клеточной стенки [2] [3] .
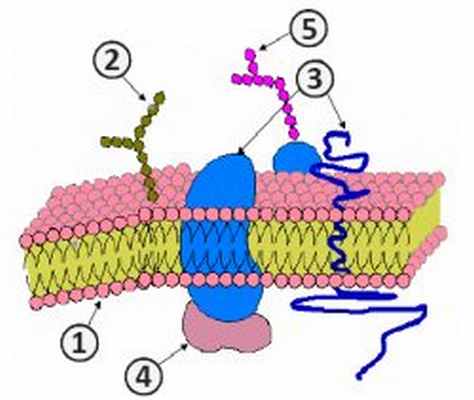
Структура цитоплазматической мембраны
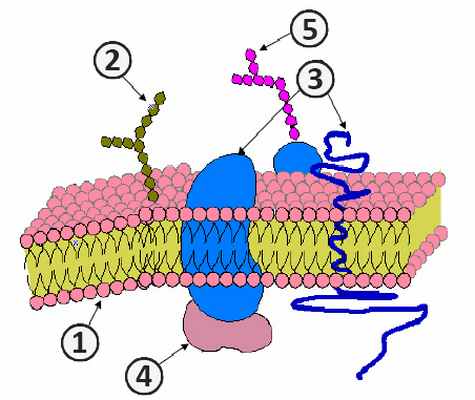
1. Фосфолипиды; 2. Гликолепиды; 3. Интегральные белки; 4. Периферические белки; 5. Олигосахариды [1] .
Бактериальная клетка, как и любая другая клетка прокариот, имеет цитоплазму, окруженную цитоплазмотической мембраной. Цитоплазма и цитоплазматическая мембрана составляют протопласт. Снаружи от него располагаются поверхностные структуры. К ним относятся: клеточная стенка, капсулы, чехлы, слизистые слои, жгутики, ворсинки и прочие структуры [1] .
Состав ЦПМ
Толщина цитоплазматической мембраны бактериальной клетки обычно составляет около 6–8 нм. На ее долю приходится до 15% сухой массы клетки [3] .
Состоит ЦПМ из липидов (15–45%), белков (45–60%) и незначительного количества углеводов (около 10%) [3] .
Липиды представлены фосфолипидами – до 30% сухой массы самой мембраны. Преобладают фосфатидилглицерин и дифосфатидилглицерин. В меньшем количестве представлены фосфатидилинозит и фосфатидилэтаноламин. Кроме того, обнаружены гликолипиды, каротиноиды, хиноны [3] .
В составе липидов присутствуют нетипичные жирные кислоты (ненасыщенные и мононасыщенные), циклопропановые и разветвленные жирные кислоты. Набор жирных кислот и состоящих из них липидов для прокариот является видоспецифичным признаком [3] .
Белки составляют половину и более сухой массы мембран. Их насчитывается более 20 типов. Они подразделяются на интегральные (погружены в гидрофобную область мембраны) и периферические (локализованы на поверхности гидрофильного слоя и часто прикреплены к интегральным белкам) [3] .
Углеводы в мембране взаимосвязаны с белками и липидами. Они обычно локализованы только на наружной поверхности и выполняют функции рецепторов опознавания факторов внешней среды [3] .
Структура ЦПМ
Цитоплазматическая мембрана бактерий, как и все прочие биологические мембраны, является асимметричной жидкокристаллической структурой. Ее асимметрия обусловлена химическим строением молекул белка и их расположением в липидном слое. Одни белки располагаются на поверхности, другие – погружены в него, третьи проходят насквозь от внутренней до внешней поверхности бислоя. Строго определенная ориентация мембранных белков обусловлена их синтезом и асимметричным включением в мембрану [3] .
Наружная и внутренняя поверхности ЦПМ различаются по ферментативной активности [3] .
В зависимости от условий окружающей среды, в частности от температуры, ЦПМ находится в различных фазовых состояниях: разжиженном или кристаллическом. При переходе из одной фазы в другую меняется подвижность компонентов мембраны, плотность ее упаковки. Это может приводить к нарушениям в функциональной активности ЦПМ [3] .
Функции ЦПМ
Цитоплазматическая мембрана выполняет ряд существенных для клетки функций:
Функции цитоплазматической мембраны бактерий (прокариот)
• Различные молекулы проходят через цитоплазматическую мембрану за счет пассивной диффузии или активного перемещения
• Большинство растворенных веществ перемещается через мембрану с участием специфических транспортных белков
• Цитоплазматическая мембрана поддерживает протонный градиент между цитоплазмой и внеклеточной средой
Клетки всех организмов обладают цитоплазматической (или плазматической) мембраной, которая предотвращает выход и поступление в клетку растворимых соединений. У прокариот цитоплазматическая мембрана, толщиной примерно 8 нм, служит барьером между содержимым клетки и окружающей средой. Цитоплазматические мембраны содержат липиды и белки. Так же как и для других биологических мембран, в основе их структуры лежит фосфолипидный бислой. Фосфолипиды содержат фосфатную группу, присоединенную к основной трехуглеродной структуре глицерина.
Гидрофобные цепи жирных кислот связаны со свободными атомами углерода этой структуры, причем они ориентированы по направлению друг к другу и в противоположную сторону от внешней среды, содержащей воду. Напротив, гидрофильные фосфатные группы ориентированы в сторону водного окружения. Цитоплазматическая мембрана не дает возможность большинству биологических молекул и ионов пассивно диффундировать через нее в обоих направлениях. Исключение составляет вода, которая способна к медленной диффузии через цитоплазматическую мембрану; небольшой размер молекул и отсутствие заряда позволяют ей свободно проходить через фосфолипидный бислой.
С цитоплазматической мембраной связано много типов белков. Часто мембранные белки содержат участки гидрофобных аминокислот, которые взаимодействуют с гидрофобными цепями жирных кислот, находящихся в мембране. Белки, гидрофобные участки которых пронизывают мембрану, называются интегральными мембранными белками. Многие белки, находящиеся в цитоплазматической мембране, играют определенную роль в транспорте молекул в клетку и в выходе из нее. Такой транспорт через плазматическую мембрану может представлять собой активный или пассивный процесс. При пассивном транспорте молекулы движутся по концентрационному градиенту, т. е. из области их высокой концентрации в область с низкой концентрацией.
Таким образом, пассивный транспорт не требует затрат энергии. В противоположность пассивной диффузии, в результате активного транспорта, внутри клетки и вне ее устанавливаются различные концентрации растворенных метаболитов. Транспортные системы состоят из белков, которые связаны или с мембраной в виде ее стабильных интегральных компонентов, или с расположенными на периферии липидами через модифицированные концевые аминогруппы, за счет которых белок прикрепляется к поверхности мембраны. Такие большие молекулы, как белки, которые не могут свободно диффундировать через мембраны, транспортируются в процессе активного транспорта. Часто транспортные системы проявляют высокую специфичность, транспортируя только молекулы одного вещества или группы веществ.
Комплекс транспортных белков, связывающий АТФ (ABC), представляет собой самое обширное семейство транспортных белков прокариот. Только у Е. coli оно насчитывает свыше 200 белков. ABC-переносчики могут перемещать субстраты в клетку и из нее. Они проявляют активность по отношению к разнообразным субстратам, от ионов до молекул белков. ABC-переносчики, транспортирующие субстраты в клетки грамотрицательных бактерий, обычно состоят из трех компонентов: транспортный белок, пронизывающий мембрану, периплазматический белок, связывающий субстрат, и белок, гидролизующий АТФ, локализованный в цитоплазме. Периплазматические связывающие белки проявляют крайне высокое сродство к специфическим субстратам.
Это позволяет им осуществлять транспорт субстратов, даже если они присутствуют в крайне низких концентрациях. Белки связывающие АТФ, расположенные на цитоплазматической стороне мембраны, обеспечивают энергией процесс транспорта. Эти белки стабильно связаны с мембраной через липидную часть со стороны N-концевого участка.
Цитоплазматическая мембрана играет важную роль в энергетических процессах у прокариот. Электроны, которые генерируются в процессе дыхания, сопрягаются со своими рецепторами в мембране. Протоны транспортируются к клеточной поверхности посредством мембранных транспортных белков; при этом наружная мембрана приобретает слабый положительный заряд.
Внутренняя часть мембраны заряжена отрицательно. Таким образом, создается трансмембранный протонный градиент. Энергетически выгодное движение протонов по градиенту, от наружной поверхности вглубь клетки, обеспечивает протекание многих внутриклеточных реакций. Иными словами, энергия, запасенная в цитоплазматической мембране может использоваться клеткой для разных целей. Например, ферменты используют протонный градиент для образования АТФ из АДФ. Некоторые мембраносвязанные ферментные комплексы способствуют генерации протонного градиента в ходе окислительного фосфорилирования.
При окислительном фосфорилировании терминальным акцептором электронов служит кислород. Однако в анаэробных условиях многие прокариоты могут использовать другие акцепторы электронов, например серу, азот, железо и марганец. Энергия, которая запасается в цитоплазматической мембране за счет функционирования ферментов, обеспечивает большую часть потребностей растущей клетки, т. е. такие процессы, как синтез молекул, транспорт белков и субстратов, а также ее подвижность. Изучение молекулярных механизмов генерации энергии в бактериальных клетках представляет собой активно развивающуюся область современных исследований.
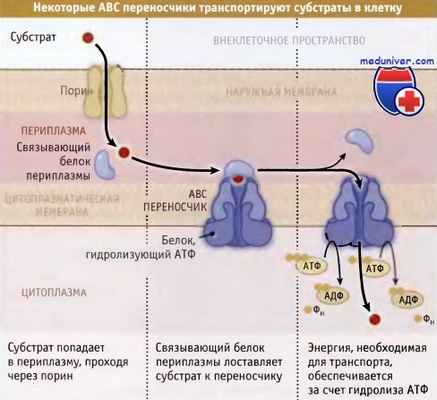
У грамотрицательных бакерий растворимые субстраты поступают в периплазму при участии поринов,
представляющих собой белки, которые образуют в наружной мембране определенных размеров поры.
Субстраты связываются специфическими белками, присутствующими в периплазме, которые, в свою очередь,
в цитоплазматической мембране связываются с ABC переносчиками.
Для транспортировки субстратов в цитоплазму, эти переносчики используют энергию гидролиза АТФ.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Функции цитоплазматической мембраны бактерий (прокариот)
Структура бактериальной клетки
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
Обозначения:
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40—90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Липополисахарид наружной мембраны состоит из трех фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (лат. core — ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельнои О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима,
пенициллина, защитных факторов организма образуются клетки с измененной (часто шаровидной) формой: протопласты — бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами.
Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку.
Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактомазы) и компоненты транспортных систем.
Цитоплазматическая мембрана
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым - промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами.
Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Рибосомные РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными веществами для питания и энергетических потребностей бактерий. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды, представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слиэь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима в воде.
Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Кроме синтеза
экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны.
Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий
с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
Читайте также: