Классический путь активации комплемента
Добавил пользователь Morpheus Обновлено: 27.01.2026
Компоненты системы комплемента выполняют множество биологических функций (например, стимуляция хемотаксиса, запуск дегрануляции тучных клеток независимо от иммуноглобулина E [IgE]).
Активация комплемента
Пути активации комплемента
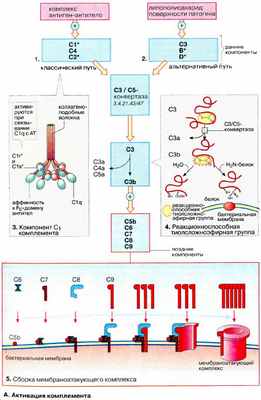
Классический, лектиновый и альтернативный пути сходятся в 1, в котором СЗ-конвертаза расщепляет СЗ на СЗа и СЗb. АТ = антитело; АГ = антиген; C1-INH = С1- ингибитор; MAC = комплекс мембранной атаки; MASP = MBL-ассоциированная сериновая протеаза; MBL = маннозосвязывающий лектин. Верхняя черта означает активацию.
Компоненты классического пути обозначаются буквой С и цифрой (например, С1, С3), обозначающей очередность их идентификации. Компоненты альтернативного пути часто обозначаются буквами (например, фактор B, фактор D) или имеют название (например, пропердин).
Активация классического пути может быть
антитело-зависимая, возникающая при взаимодействии C1 с антиген-IgM или агрегированными антиген-IgG комплексами
АТ-независимая, возникающая когда полианионы (например, гепарин, протамин, ДНК и РНК из апоптотических клеток), грамотрицательные бактерии или связанный С-реактивный белок реагируют непосредственно с С1
Этот путь регулируется С1-ингибитором (C1-INM). Наследственная ангиоэдема Наследственный ангионевротический отек И наследственный, и приобретенный ангионевротический отек (приобретенный дефицит ингибитора С1-белка) вызваны дефицитом или дисфункцией ингибитора комплемента 1 (С1), белка, участвующего в регуляции. Прочитайте дополнительные сведенияАктивация лектинового пути – АТ-независимый процесс; он возникает, когда маннозосвязывающий лектин (МСЛ), протеин сыворотки, связывается с маннозой, фруктозой или N-ацетилглюкозаминовыми группами на клеточных стенках бактерий и дрожжей или на вирусах. В остальном этот путь структурно и функционально напоминает классический путь.
Активация альтернативного пути осуществляется тогда, когда компоненты поверхностей клеток микроорганизмов (например, клеточных стенок дрожжей, липополисахариды клеточной стенки бактерий [эндотоксин]) или иммуноглобулин (например, нефротический фактор, агрегированный IgA) расщепляют небольшое количество С3. Этот путь регулируется пропердином, фактором Н, и фактором, ускоряющим некроз (CD55).
Эти три пути в итоге сходятся в один, где СЗ-конвертаза расщепляет СЗ на СЗа и СЗb (см. рисунок Пути активации комплемента [Complement activation pathways] Пути активации комплемента ). Расщепление С3 может привести к образованию комплекса атаки на мембрану (МАС), цитотоксическому компоненту системы комплемента. МАС является причиной лизиса чужеродных клеток.
Фактор I с кофакторами, включая мембранный кофакторный белок (CD46), инактивирует C3b и C4b.
Дефекты и дефицит системы комплемента
Недостатки или дефекты в конкретных компонентах комплемента связаны с конкретными нарушениями; ниже приведены примеры:
Дефицит С1, С2, С3, MBL, MBL-ассоциированной серин-протеаза (MASP-2), фактора Н, фактора I или рецептора комплемента 2 типа (CR2): повышение риска рецидивирующих бактериальных инфекций
Дефицит C5, C9, фактора B, фактора D или пропердина: восприимчивость к нейссериальным инфекциям Менингококковая инфекция Менингококки (Neisseria meningitidis) – это грамотрицательные кокки, которые вызывают менингит и менингококкемию. Симптомы обычно тяжелые, включают головную боль, тошноту, рвоту, светобоязнь. Прочитайте дополнительные сведения Дефекты С1, С4 и С5: системная красная волчанка Системная красная волчанка (СКВ) Системная красная волчанка – хроническое мультисистемное воспалительное заболевание аутоиммунной природы; поражает преимущественно молодых женщин. Наиболее часто заболевание проявляется артралгиями. Прочитайте дополнительные сведения Мутации генов фактора В, фактора Н, фактора I, кофактора мембранных белков (CD46) или С3: развитие атипичного варианта гемолитико-уремического синдрома Гемолитико-уремический синдром (ГУС) Гемолитико-уремический синдром (ГУС) является острым скоротечным заболеванием, для которого характерна тромбоцитопения, микроангиопатическая гемолитическая анемия и острое поражение почек. ГУС. Прочитайте дополнительные сведенияБиологическая активность системы комплемента
Компоненты системы комплемента выполняют и другие биологические функции, которые реализуются рецепторами комплемента (CR) на различных типах клеток. Некоторые РК (CR) используют молекулы, которым был назначен номер CD.
CR1 (CD35) способствует фагоцитозу и участвует в выведении иммунных комплексов.
CR2 (CD21) регулирует продукцию АТ В-лимфоцитами и является рецептором вируса Эпштейна – Барр.
CR3 (CD11b/CD18), SR4 (CD11c/CD18) и рецепторы C1q играют роль в фагоцитозе.
С3а, С5а и С4а (слабо) проявляют анафилатическую активность: они вызывают дегрануляцию тучных клеток, ведущую к повышению проницаемости сосудов и сокращению гладкой мускулатуры.
С3b работает в качестве опсонина, покрывая микрооорганизмы и, тем самым, усиливая их фагоцитоз.
С3d усиливает продукцию АТ В-лимфоцитами.
С5а является хемоаттрактантом нейтрофилов. Он контролирует активность нейтрофилов и моноцитов и может стать причиной повышенного слипания клеток, дегрануляции и высвобождения внутриклеточных ферментов из гранулоцитов, продукции токсических метаболитов кислорода и других действий, связанных с клеточным метаболизмом.
Авторское право © 2022 Merck & Co., Inc., Rahway, NJ, США и ее аффилированные лица. Все права сохранены.
Классический путь активации комплемента
Ткани и органы. Иммунная система
Система комплемента является частью иммунной системы, она осуществляет неспецифическую защиту от бактерий и других проникающих в организм возбудителей болезней. Система комплемента состоит примерно из 20 различных белков — «факторов (компонентов) комплемента», которые находятся в плазме крови и составляют около 4% от всех белков плазмы.
А. Активация комплемента
Система комплемента может действовать тремя различными способами:
• через хемотаксис: различные компоненты (факторы) комплемента могут привлекать иммунные клетки, которые атакуют бактерии и пожирают их (фагоцитируют);
• через лизис: компоненты комплемента присоединяются к бактериальным мембранам, в результате чего образуется отверстие в мембране и бактерия лизируется;
• через опсонизацию: компоненты комплемента присоединяются к бактерии, в результате чего образуется метка для узнавания фагоцитирующими клетками (например, макрофагами и лейкоцитами). имеющими рецепторы к компонентам комплемента.
Реакции системы комплемента осуществляются, как правило, молекулами на поверхности микроорганизма. Факторы (компоненты) с С1 по С9 (С от англ. complement) формируют так называемый «классический путь» ( 1 ) активации комплемента, факторы В и D участвуют в активации «альтернативного пути» ( 2 ). Другие компоненты системы комплемента, здесь не показанные, выполняют регуляторные функции.
Ранние компоненты системы комплемента являются сериновыми протеиназами (см. с. 178). Они создают амплифицирующий ферментативный каскад реакций. Классический путь инициируется связыванием компонента C1 ( 3 ) с несколькими молекулами IgG или с пентамерным IgM на поверхности микроорганизма (см. ниже). Альтернативный путь инициируется связыванием фактора В , например, с бактериальным липополисахаридом (эндотоксином). Оба пути ведут к расщеплению компонента С3 комплемента на два фрагмента, обладающих различными функциями. Меньший фрагмент С3а принимает участие в развитии воспалительного процесса, индуцируя хемотаксис лейкоцитов к очагу воспаления (хемотаксис, воспалительные процессы). Более крупный фрагмент С3b связывается ковалентно на поверхности бактериальной клетки и инициирует цепь реакций, приводящих к образованию мембраноатакующего комплекса (см. ниже) поздними компонентами системы комплемента.
Компонент C1 комплемента представляет собой сложный молекулярный комплекс, состоящий из трех различных компонентов C1q, C1r и C1s ( 3 ). Гексамерный C1q по форме напоминает букет нераскрытых тюльпанов, «бутоны» которого могут связываться с F с -фрагментом антител. При связывании нескольких C1q с антителами активируется серин-протеиназа C1r, с которой начинается протеолитический каскад классического пути.
Компонент СЗ комплемента стоит в центре активации системы. СЗ подвергается протеолизу СЗ-конвертазой с расщеплением на СЗа и СЗb фрагменты и участвует в формировании С5-конвертазы. СЗ-конвертаза представляет собой комплекс из C4b и С2а (в случае классического пути) или из СЗb и Bb (в случае альтернативного пути). При гидролизе СЗ в СЗb становится доступной очень реакционноспособная тиолслож-ноэфирная группа, которая реагирует с гидроксильной или аминогруппой ( 4 ). Вследствие этого СЗb ковалентно связывается с молекулами бактериальной мембраны (опсонизация).
Мембраноатакующий комплекс — это ионный канал (пора), в плазматической мембране бактериальной клетки, в формировании которого участвуют компоненты СЗb, С5b, С6, С7, С8 и главным образом С9 ( 5 ). При этом молекулы С9 последовательно присоединяются к агрегату, формируя кольцевую структуру, через центр которой могут диффундировать небольшие молекулы, такие, как вода и ионы. Осмотические силы способствуют «накачиванию» воды внутрь бактериальной клетки, которая набухает и лопается (лизирует).
Активность системы комплемента контролируется ингибиторами в плазме крови, блокирующими избыточную реакцию.
26.2. Пути активации комплемента
Существуют три пути активации комплемента: классический, лектиновый и альтернативный.
А. Классический путь активации комплемента является основным. Участие в этом пути активации комплемента – главная функция антител.
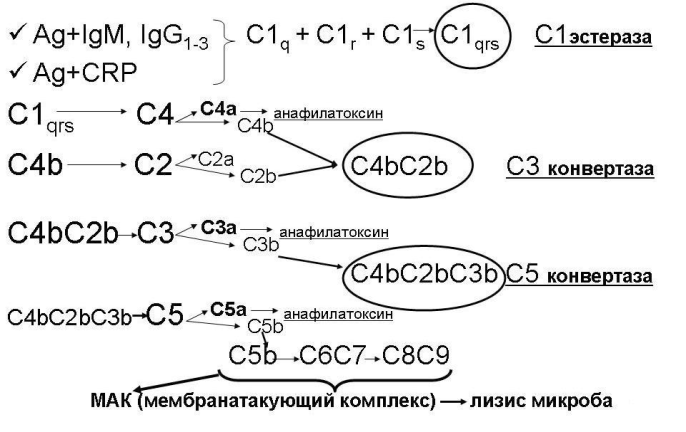
Рис 26.2-2. Схема классического пути активации комплемента
1. Активацию комплемента по классическому пути запускает иммунный комплекс: комплекс антигена с иммуноглобулином (класса G – первых трех подклассов – или М). Место антитела может «занять» С-реактивный белок – такой комплекс также активирует комплемент по классическому пути.
2. Классический путь активации комплемента осуществляется следующим образом (рис 26.2-1).
а. Сначала активируется фракция С1: она собирается из трех субфракций (C1q, C1r, C1s) и превращается в фермент С1-эстеразу (С1qrs).
б. С1-эстераза расщепляет фракцию С4.
в. Активная фракция С4b ковалентно связывается с поверхностью микробных клеток (но не с собственными эукариотическими клетками макроорганизма) с здесь присоединяет к себе фракцию С2.
г. Фракция С2 в комплексе с фракцией С4b расщепляется С1-эстеразой с образованием активной фракции С2b.
д. Активные фракции С4b и С2b в один комплекс – С4bС2b – обладающий ферментативной активностью. Это так называемая С3-конвертаза классического пути.
е. С3-конвертаза расщепляет фракцию С3, нарабатываю большие количества активной фракции С3b.
ж. Активная фракция С3b присоединяется к комплексу С4bС2b и превращает его в С5-конвертазу (С4bС2bС3b).
з. С5-конвертаза расщепляет фракцию С5.
и. Появившаяся в результате этого активная фракция С5b присоединяет фракцию С6.
к. Комплекс С5bС6 присоединяет фракцию С7.
л. Комплекс С5bС6С7 встраивается в фосфолипидный бислой мембраны микробной клетки.
м. К этому комплексу присоединяется белок С8.
н. Будучи вместе со всем комплексом в фосфолипидный бислой мембраны микробной клетки, белок С8 катализирует полимеризацию 10 – 16 молекул белка С9. Данный полимер формирует в мембране микробной клетки неспадающую пору диаметром около 10 нм (рис 26.2-2)., что приводит к лизису микроба (так как на его поверхности образуется множество таких пор – «деятельность» одной единицы С3-конвертазы приводит к появлению около 1000 пор). Комплекс С5bС6С7С8С9, образующийся в результате активации комплемента, называется мемранатакующим комплексом (МАК).
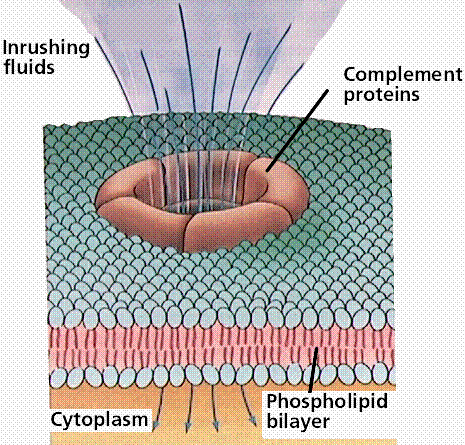
Рис. 26.2-2. Схема образования МАК (слева) и результат активации комплемента – формирование поры в фосфолипидном бислое микробной мембраны, приводящей к осмотическому лизису микробной клетки (справа)
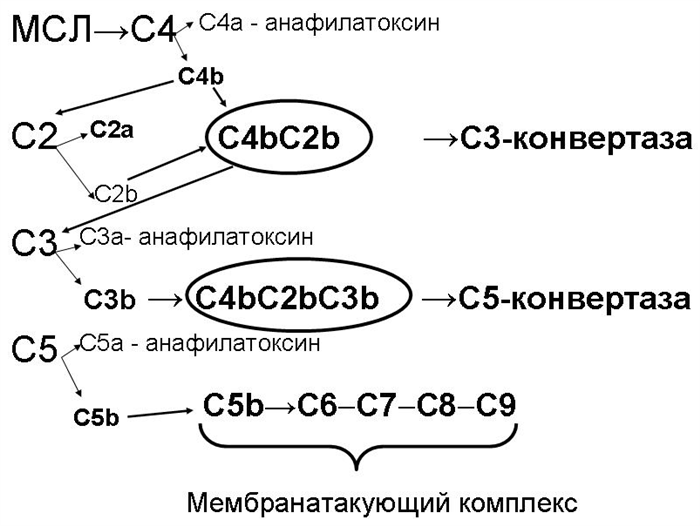
Рис 26.2-3. Схема лектинового пути активации комплемента
Б. Лектиновый путь активации комплемента запускается комплексом нормального белка сыворотки крови – маннансвязывающего лектина (МСЛ) – с углеводами поверхностных структур микробных клеток (с остатками маннозы). Активизирующаяся в результате этого процесса МСЛ-ассоциированная сериновая протеаза действует аналогично С1-эстеразе классического пути, по которому, собственно, и развиваются дальнейшие события, заканчивающиеся формированием МАК (рис. 26.2-3).
В. Альтернативный путь активации комплемента (рис. 26.2-4) начинается с ковалентного связывания активной фракции С3b – которая всегда присутствует в сыворотке крови в результате постоянно протекающего здесь спонтанного расщепления фракции С3 – с поверхностными молекулами не всех, но некоторых микроорганизмов.
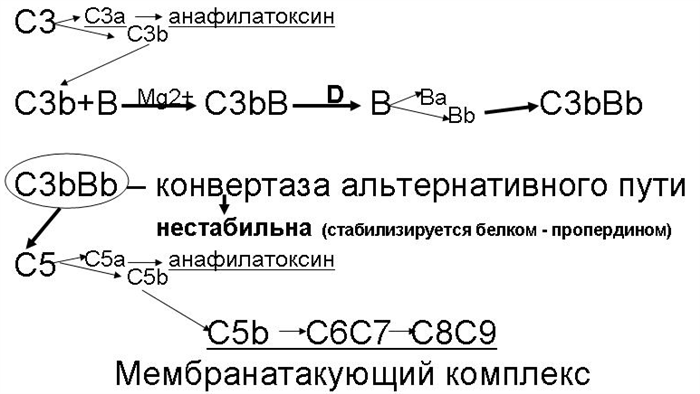
Рис. 26.2-4. Схема альтернативного пути активации комплемента
1. Дальнейшие события развиваются следующим образом.
а. С3b связывает фактор В (который структурно и функционально гомологичен фактору С2), образуя комплекс С3bВ.
б. В связанном с С3b виде фактор В выступает в качестве сусбтрата для фактора D (сывороточной сериновой протеазы), которая расщепляет его с образованием активного комплекса С3bВb. Этот комплекс обладает ферментативной активностью, структурно и функционально гомологичен С3-конвертазе классического пути (С4bС2b) и называется С3-конвертазой альтернативного пути.
в. Сама по себе С3-конвертаза альтернативного пути нестабильна. Чтобы альтернативный путь активации комплемента успешно продолжался этот фермент стабилизируется фактором Р (пропердином).
г. То, что происходит дальше, аналогично классическому пути активации комплемента.
1. Нарабатывается много С3b и образуется комплекс С3bВbС3b, являющийся С5-конвертазой.
2. Активация С5 дает начало образованию мембранатакующего комплекса (см. разделы 26.2.А.2.и – 26.2.А.2.н).
2. Основное функциональное отличие альтернативного пути активации комплемента, по сравнению с классическим, заключается в быстроте ответа на патоген: так как не требуется время для накопления специфических антител и образования иммунных комплексов.
Г. Важно понимать, что и классический и альтернативный пути активации комплемента действуют параллельно, еще и амплифицируя (т.е. усиливая) друг друга. Другими словами комплемент активируется не «или по классическому или по альтернативному», а «и по классическому и по альтернативному» путям активации. Это, еще и с добавлением лектинового пути активации, – единый процесс (см. рис. 26.2-5), разные составляющие которого могут просто проявляться в разной степени.
Альтернативный путь активации системы комплемента. Понятие об инициирующем сигнале
Система комплемента – тонкосбалансированная система, для активации которой необходим сигнал. Конечный эффект действия системы комплемента – цитолитическая реакция+образование широкого спектра БАВ, активирующих всю систему резистентности. Существует 3 основных пути активации системы комплемента: 1. Классический путь 2. Альтернативный путь 3. Лектиновый путь. Пути отличаются: 1. Инициирующим сигналом 2. Молекулярным составом главной С3 конвертазы.
Альтернативный путь активации.
В обычных условиях происходит незначительная неспецифическая фрагментация С3 компонента комплемента (синтез-ся в гепатоцитах) на С3а(БАВ) и С3b. Инициирующий сигнал-появление МКО, при этом C3b образует ковалентную связь с МКО (это пусковой механизм) С3b изменяет конформацию и к комплексу МКО-С3b присоединяется белок В, который также меняет конформацию и у него появляются сайты для специфического протеолиза белком D образуются Ва (БАВ) и Вb. Образуется комплекс МКО-С3b- Вb – это С3-конвертаза альтернативного пути (стабилизируется белком плазмы-пропердином).
После образования С3-конвертазы происходит осаживание МКО-ма молекулами С3b (до 1000 молекул). Ва и С3а активируют белоксинтезирующую функцию гепатоцитов.
С3-конвертаза является и С5-конвертазой к комплексу последовательно присоедин-ся С5b, С6, С7 (образуется МКО-С3b- Вb-C5b-C6-C7). При присоединении белок С7 меняет конформацию, увеличивается его гидрофобность и он проникает в стенку МКО. Весь комплекс на поверхности МКО присоединяет С8 и С9 (до 16-20 молекул С9). Образуется мембрано-атакующий комплекс, который образует ниспадающие поры в мембране, происходит лизис МКО, образуются фрагменты МКО-мусор, который накапливается в организме и является мощным активатором клеточных элементов неспецифической системы резистентности.
7. Мембрано-атакующий комплекс – молекулярные механизмы формирования, эффекты, реализуемые с его участием.
В результате активации компонентов комплимента по любому из путей образуется МАК. Одна молекула С3-конвертазы классического (С4b2a), лектинового (С4b2a) или альтернативного пути (С3bBb) расщепляет сотни молекул С3 на С3а и С3b. Функции С3b: 1) при связывании с С4b2a образуется комплекс С4b2a3b который называется С5-конвертазой классического (лектинового) пути; 2) при связывании с фактором В образуется комплекс С3bBb, который называется С3-конвертазой альтернативного пути; 3) при связывании с С3bBb образуется комплекс С3bBb3b, который называется С5-конвертазой альтернативного пути
1) С5-конвертазы классического (С4b2a3b), лектинового (С4b2a3b) и альтернативного (С3bBb3b) путей расщепляют С5-компонент комплимента на С5а (БАВ) и С5b;
2) к С5b присоединяется С6, образуется С5b6;
3) к С5b6 присоединяется С7, образуется С5b67, что приводит к изменению конформации, образованию гидрофобного центра, в результате чего вся структура заякоривается на поверхности микроорганизма;
4) к С5b67 присоединяется С8, образуется С5b678 – мембрана частично повреждается, клетка медленно разрушается;
5) к С5b678 присоединяется С9, образуется С5b6789 – МАК (мембрано-атакующий комплекс).
МАК - ниспадающая пора, диаметром до 30 нм, Мr до 2 кДа. Количество таких пор образуется на поверхности клетки мко около 1000. Через поры устремляются вода и электролиты, клетка становится нежизнеспособной и погибает.
8. Классический путь активации системы комплемента – молекулярные особенности прохождения. Инициирующий сигнал.
Система комплемента – тонкосбалансированная система, для активации которой необходим сигнал. Существует 3 основных пути активации системы комплемента: 1. Классический путь 2. Альтернативный путь 3. Лектиновый путь. Пути отличаются: 1. Инициирующим сигналом 2. Молекулярным составом главной С3 конвертазы.
Классический путь: инициируется комплексом АГ-АТ (IgG, IgM). Стадии процесса:
1) Активация компонента С1. Состоит из 3 компонентов: C1q (6 молекул), C1r, C1s. После связывания С1q с иммунным комплексом (с Fc-областями IgG или IgM) происходит активация C1r и C1s (протеазы).
2) Расщепление активированным компонентом С1 компонента С4 с образованием С4а и С4b. С4b может затем связаться либо с комплексом антитело-С1, либо с поверхностью микроба, либо с компонентом С2, в результате чего получится комплекс С4b2 – новый субстрат для С1s.
3) Образование из С4b2 комплекса С4b2а, который является С3-конвертазой классического пути.
4) Расщепление ферментом С3-конвертазой классического пути компонента С3 с образованием С3b, который затем активирует остальные компоненты комплимента. Далее пути прохождения одинаковые с лектиновым и альтернативным (образование С5-конвертазы, образование мембраноатакующего комплекса, гибель клетки) (см вопрос 7).
9. Лектиновый путь активации комплемента – молекулярные особенности прохождения. Инициирующий сигнал.
Лектиновый путь активации комплемента почти идентичен классическому, но запускается независимо от антител. Лектиновый путь инициируется связывающим маннозу лектином (МСЛ), который связывает углеводные структуры (остатки маннозы) микробной поверхности.
СМЛ относится к семейству кальций-зависимых лектинов, названных коллектинами (коллагеновые лектины). СМЛ может связываться с концевыми маннановыми группами на поверхности клеток бактерий, приобретая за счет этого способность к взаимодействию с двумя маннан-связывающими лектин- ассоциированными сериновыми протеиназами.
СМЛ - ассоциированная сериновая протеаза (МASP1 (гомолог С1r), МASP2 (гомолог С1s)) катализирует активационное расщепление С4 и С2. Взаимодействие СМЛ - МASP1- МASP2 аналогичнообразованию комплекса C1q—C1r—C1s в классическом пути. В дальнейшем активация комплемента происходит так же, как и по классическому пути.
Таким образом, лектиновый пути инициируют первичный иммунный ответ даже в той ситуации, когда антиген еще не распознан антителами или иммунокомпетентными клетками.
Классический путь активации системы комплемента
Схема классического и альтернативного путей активации комплемента. Первые два этапа — инициация, следующие два — формирование C3-конвертазы, последние два — формирование C5-конвертазы и мембраноатакующего комплекса
Класси́ческий пу́ть актива́ции систе́мы комплеме́нта — один из трёх путей активации системы комплемента, наряду с альтернативным путём и лектиновым путём. Классический путь запускают комплексы антиген-антитело ( иммунные комплексы [en] ), причём антитела должны принадлежать к классу IgG или IgM. После активации происходит сборка белкового комплекса C3-конвертазы [en] (C4b2b), который разрезает белок комплемента C3 [en] . Один из фрагментов C3, C3b [en] , связывается с C3-конвертазой, в результате чего образуется C5-конвертаза [en] (C4b2b3b). C5-конвертаза разрезает компонент комплемента 5, после чего образовавшиеся белковые фрагменты привлекают фагоциты в очаг инфекции и способствуют поглощению клеток патогенов. C5-конвертаза также запускает финальные стадии каскада комплемента, которые завершаются формированием мембраноатакующего комплекса. Он формирует поры в мембране клетки микроорганизма, вызывая её лизис и гибель. Помимо микробных клеток, классический путь комплемента могут запускать апоптотические тельца и некротизированные клетки [1] [2] [3] [4] .
Классический путь активации комплемента был открыт в 1888 году Жюлем Борде и стал первым описанным механизмом активации системы комплемента [5] .
Содержание
Механизм
Инициация
Классический путь инициируется при связывании белка комплемента C1 [en] с доменом CH2 молекулы иммуноглобулина G (IgG) или доменом CH3 молекулы иммуноглобулина M (IgM), которые уже связались с антигеном. Среди антител IgG классический путь наиболее эффективно активируют IgG3 и IgG1 (у человека). Белок C1 состоит из субъединиц C1q [en] , C1r [en] и C1s [en] , причём C1q связывается с антителом, а C1r и C1s являются протеазами. C1q представляет собой гексамер, который специфически связывается с участками Fc тяжёлых цепей типа μ и некоторыми тяжёлыми цепями типа γ. Классический путь комплемента не может быть активирован свободными антителами, но только антителами, связанными с соответствующим антигеном, причём для активации необходимо, чтобы C1 связался с двумя и более участками Fc. Поскольку каждая молекула IgG имеет только один участок Fc, для связывания с C1 необходимо, чтобы две или более молекулы IgG находились рядом. Хотя свободные IgM в плазме крови являются пентамерами [en] , путь комплемента не может быть активирован при связывании лишь с одной молекулой IgM, поскольку участки Fc каждого мономера располагаются так, что они не могут быть связаны одной молекулой C1. Одна молекула IgM, будучи пентамером, может связать две молекулы C1, поэтому IgM активирует комплемент эффективнее, чем IgG. C1r и C1s являются сериновыми протеазами и формируют тетрамер, в котором C1r и C1s имеются в числе двух молекул [6] .
Формирование C3-конвертазы
Когда C1q связывается с IgG или IgM, связанный C1r активируется и вносит разрез в C1s, активируя его. Активированный C1s разрезает следующий белок каскада, C4 [en] , с образованием C4b. Как и C3b, C4b содержит внутреннюю тиоэфирную связь, которая обеспечивает ковалентное сшивание C4b с комплексом антиген-антитело на поверхности микробной клетки или непосредственно с поверхностью клетки. Следующий участник каскада, C2 [en] , связывается с C4b, ковалентно прикреплённым к поверхности клетки, и разрушается C1s с образованием фрагмента C2b, функции которого неизвестны. При этом C2a остаётся связанным с C4b на поверхности клетки патогена (в отличие от остальных компонентов комплемента, у C2 больший фрагмент называется C2a, а меньший фрагмент C2b, высвобождаемый при расщеплении С2, остаётся несвязанным). Комплекс C4b2a является C3-конвертазой и может связывать C3 и ферментативно расщеплять его. Связывание C3 с C3-конвертазой обеспечивается C4b, а C2a катализирует протеолиз C3. При расщеплении C3 образуются два фрагмента, меньший из которых, C3a, удаляется, а C3b может ковалентно связываться с белками клеточной поверхности или антителами, связанными с клеткой, на поверхности которой был активирован каскад комплемента. C3b также может взаимодействовать с фактором B и образовывать больше C3-конвертаз по альтернативному пути активации комплемента. Одна C3-конвертаза в итоге может давать начало сотням и тысячам молекул C3b на поверхности клетки, где был активирован комплемент. Ранние этапы классического и альтернативного путей комплемента имеют много похожего: C3 в альтернативном пути гомологичен C4 классического пути, а фактор B гомологичен C2. Некоторые молекулы C3b связываются с C3-конвертазой с образованием комплекса C4b2a3b, который является C5-конвертазой. C5-конвертаза расщепляет C5 [en] и запускает поздние стадии каскада комплемента [6] .
При инфекциях, вызванных пневмококками, запускается независимый от антител, но зависимый от C1 вариант классического пути, который активируется при связывании углеводов с лектинами на поверхности клетки. Некоторые макрофаги экспрессируют лектин C-типа, известный как SIGN-R1, который распознаёт полисахариды пневмококков и связывается с C1q. Благодаря этому активируется классический путь комплемента, в результате работы которого клетка пневмококка покрывается C3b [7] .
Формирование C5-конвертазы и мембраноатакующего комплекса
Клиническое значение
Недостаточность по белку C1q может стать причиной развития системной красной волчанки [4] [9] . Помимо других функций, C1q запускает удаление из тканей и сосудов апоптотических телец и иммунных комплексов [1] [10] . При его недостаточной активности иммунные комплексы и апоптотические тельца накапливаются, вызывая воспаление и аутоиммунные процессы, при которых образуются аутоантитела [3] . Изучается возможность использования аутоантител к C1q в качестве молекулярного маркера [en] системной красной волчанки [11] [12] .
Чрезмерная активность классического пути комплемента при недостаточной работе C1-ингибитора [en] может приводить к эпизодической ангиоэдеме [1] . Недостаточность по C1-ингибитору может быть наследственной или приобретённой [13] . В норме C1-ингибитор инактивирует C1r и C1s, мешая запуску классического пути активации комплемента. Кроме того, C1-ингибитор контролирует проницаемость сосудов. Концентрация C1-ингибитора, составляющая 50 % и менее от нормальной, приводит к повышению проницаемости сосудов, из-за чего развивается ангиоэдема [13] . В 2008 году для предотвращения приступов наследственной ангиоэдемы был одобрен препарат цинриз (англ. Cinryze ), представляющий собой производное C1-ингибитора из плазмы крови человека [14] [15] .
Изучается возможность уничтожения вирионов ВИЧ с помощью классического пути активации комплемента [16] . Показана эффективность методов иммунотерапии рака, использующих активацию классического пути [17] . Классический путь комплемента особенно важен для уничтожения клеток метициллинрезистентных штаммов золотистого стафилококка, так как с ними связываются некоторые варианты IgM [18] .
Читайте также: