Нейруляция яйцеклетки. Сегментация мезодермы.
Добавил пользователь Alex Обновлено: 27.01.2026
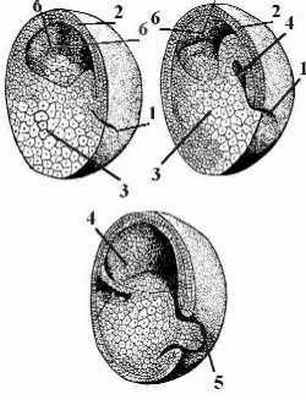
Ранняя гаструла лягушки имеет шарообразную форму, поздняя несколько вытягиваясь в длину, приобретает форму овала благодаря образованию нервных (медуллярных) валиков. Это парные утолщенные гребневидные складки эктодермы, возникающие по бокам дорсальной стороны тела зародыша. Нервные валики ограничивают широкую дорсальную полосу эктодермы, именуемую нервной пластинкой.
Нервная пластинка является зачатком спинного и головного мозга, представляя собой нейроэктодерму. Остальная часть эктодермы становится кожной (эпидермальной) эктодермой.
Приподнимаясь все выше, нервные валики начинают сближаться друг с другом, срастаются и образуют непарную ганглиозную пластинку или нервный гребень. Одновременно нервная пластинка сворачивается в трубку.
Нервная трубка и ганглиозная пластинка погружаются под кожную эктодерму, последняя смыкается над ними в непрерывный слой и дифференцируется в эпидермис. После срастания кожной эктодермы ганглиозная пластинка оказывается зажатой между нею и нервной трубкой (рис. 9).
Мезодерма, образующая вначале однородный слой клеток между экто и энтодермой, затем дифференцируется на дорсальные участки –спинные сегменты (сомиты), сегментные ножки (нефротомы), расположенные вентрально боковые пластинки (спланхнотом). Сомиты и нефротомы сегментируются, подразделяясь на все большее количество парных метамерных участков. Спланхнотомы правой и левой стороны тела срастаются друг с другом под кишечной трубкой, так что целомическая полость становится общей для всех сегментов и обеих сторон тела.
Висцеральный листок прирастает к кишке и другим внутренним органам и образует серозную оболочку. Париетальный прирастает изнутри к стенке тела, образуя пристеночную брюшину. Часть клеток мезодермы, выселяясь в промежутки между эмбриональными зачатками образует рыхлую клеточную массу –мезенхиму (рис. 10).
Сегментированная мезодерма делится на склеротом, прилегающий к хорде и нервной трубке, дерматом, прилегающий к эпидермальной эктодерме и находящийся между ними миотом. Из склеротома развивается осевой скелет и все виды соединительной ткани, из миотома – мышечная ткань, из дерматома – собственно кожа. Нефротомы –это материал мочеполовой системы. Из мезодермы происходит выселение клеток, которые формируют мезенхиму, заполняющую пространство между всеми закладками.
Ганглиозная пластинка разделяется на две части, которые лежат по бокам нервной трубки, образующие их клетки мигрируют в разных направлениях, а затем две части опять срастаются в одну общую пластинку.
Из ганглиозной пластинки образуются спинальные ганглии (спинно-мозговые узлы), ганглии вегетативной нервной системы, хромаффинная ткань (мозговое вещество надпочечников). В головном отделе потомки клеток нервного гребня обнаруживаются в составе костей, хрящей и гладкой мускулатуры лицевой части, косточек среднего уха, одонтобластов зубных зачатков, щитовидной и слюнных желез, дифференцируются в периферические нейроны и глию. В туловищном отделе они образуют пигментные клетки кожи (меланоциты).
На заднем конце тела зародыша кожная эктодерма нарастает как на задний невропор, так и на сильно сузившийся бластопор и прикрывает оба эти отверстия. Под сросшейся над ними кожной эктодермой невропор и бластопор оказываются соединенными друг с другом нервно-кишечным каналом. Он существует недолго и вскоре зарастает. На заднем конце зародыша образуется плотный вырост из малодифференцированных клеток ‑ хвостовая почка, представляющая собой зачаток хвоста.
На переднем конце тела слепо замкнутая энтодермальная кишечная трубка прилегает к кожной эктодерме. В этом месте эктодерма и передний конец кишечной трубки истончаются, прорываются и образуется ротовое отверстие. Таким же образом у основания хвостовой почки происходит прорыв и образование анального отверстия. Зародыш приступает к формированию отдельных органов и систем организма (органогенезу).
Развитие птиц. Особенности строения яйцеклетки птиц. Оплодотворение. Дробление.
Нейруляция яйцеклетки. Сегментация мезодермы.
Содержание темы
1. а) В прошлой теме мы рассматривали начальные этапы эмбриогенеза - оплодотворение и дробление.
б) Второй из них завершается образованием бластулы .
2. В данной теме продолжим знакомство с ранними этапами эмбрионального развития.
6.1. Гаструляция
1. Как было определено (п. 5.2.1), гаструляция приводит к появлению в зародыше 3-х листков -
наружного - эктодермы ,
среднего - мезодермы и
внутреннего - энтодермы .
2. а) Далее из этих листков развиваются осевые зачатки органов.
б) Образование некоторых осевых зачатков (напр., хорды) идёт почти одновременно с гаструляцией.
Но для удобства изложения образование всех осевых зачатков выделено нами в следующий этап.
6.1.1. Способы гаструляции
1. Гаструляция, в зависимости от вида животных, проходит различными способами.
2. Способ гаструляции во многом определяется типом бластулы, который, в свою очередь, обусловлен типом исходной яйцеклетки (п. 5.1.3.2).
6.1.1.1. Ланцетник: гаструляция путём инвагинации
одна половина бластулы постепенно впячивается во вторую половину.
первичную э к тодерму (1) - наружный слой клеток;
первичную э н тодерму (2) - внутренний слой клеток;
бластопор (3), или первичный рот, - щелевидное отверстие,
а в его составе - 4 губы -

гастроцель (6), или полость первичного кишечника, - полость, в которую ведёт бластопор.
выпячивания части первичной энтодермы в виде двух карманов,
6.1.1.2. Амфибии: гаструляция путём эпиболии
I. Общее описание
а клетки вегетативного полюса перегружены желтком и делятся медленно.
совершается за счёт быстро делящихся клеток анимального полюса
и происходит
отчасти - путём инвагинации ,
а главным образом, путём эпиболии ( обрастания ).
а) Инвагинация идёт на границе между анимальной и вегетативной частями бластулы.
Здесь образуется небольшое впячивание - серповидная бороздка (1).
б) Затем клетки анимального полюса (2) начинают энергично обрастать вегетативную часть (3) зародыша -
наползая на неё снаружи и проникая затем внутрь через край бороздки.
в) При этом серповидная бороздка всё углубляется:
внутреннее её пространство превращается в гастроцель (4) ,
а входное отверстие - в бластопор (5) ).
А в стенке последней содержатся все три зародышевых листка, включая средний.
II. Препарат
быстро делящиеся клетки (1) а ним ального полюса обрастают крупные клетки (2) вегетативного полюса, содержащие желток.
6.1.1.3. Птицы: гаструляция путём деламинации и иммиграции
деламинацию (расщепление слоя) и
иммиграцию (перемещение) клеток.
I. Первый этап - деламинация
верхний - эпибласт и
нижний - гипобласт.
образован плоскими клетками,
покрывает желток и,
по современным представлениям, в формировании тканей зародыша не участвует.
б) Из него-то (точнее, из его центральной части - зародышевого щитка) и образуются далее все 3 зародышевые листка -
эктодерма, мезодерма и энтодерма.
II. Второй этап - иммиграция
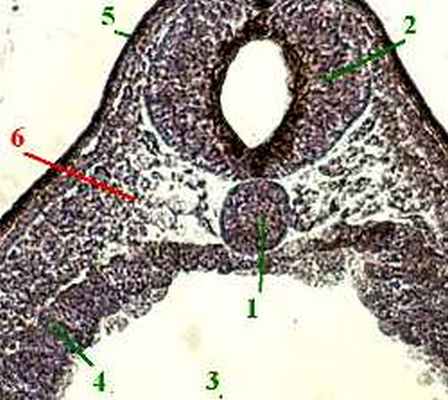
первичная полоска (1) - утолщение эпибласта с первичной бороздкой (2) посередине,
первичный (гензеновский) бугорок (3) - ещё большее возвышение в передней части первичной полоски,
в центре которого находится первичная ямка (4).
в) Эти структуры - эквивалент бластопора амфибий,
т.к. представляют собой
место активной миграции клеток внутрь с внешней поверхности зародыша.
III. Препарат
2. а) Первичная полоска (1) выглядит на поперечном срезе как утолщение в средней части зародыша;
Эктодерма (3) - наиболее толстый слой с многорядным расположением клеточных ядер.
Мезодерма (4) представлена отдельными, рыхло расположенными клетками.
Энтодерма (5) имеет вид тонкого листка, образованного уплощёнными клетками.
6.1.1.4. Млекопитающие: гаструляция путём деламинации и иммиграции
пузырёк, к стенке которого ( трофобласту ) изнутри в одном месте прилегает группа зародышевых клеток ( эмбриобласт ).
поэтому последующая гаструляция во многом подобна таковой у птиц.
I. Деламинация у млекопитающих
б) Но, кроме того, у высших животных и человека эпибласт расщепляется на два листка -
зародышевый эпибласт (2 ) и
амниотическую эктодерму (3) , -
между которыми образуется полость амниона (4).
образует зародышевый щиток (как у птиц) и
служит источником всех трёх зародышевых листков.
II. Иммиграция у млекопитающих
первичной полоски (1) и
первичного (гензеновского) узелка (2).
2. а) В этом месте внутрь проникают клетки (3), образующие энтодерму и мезодерму.
б) Остающиеся на поверхности клетки формируют эктодерму (4 ) .
6.1.2. Некоторые обобщения
6.1.2.1. Сводная таблица
Сведём воедино информацию о том, как тип яйцеклетки у животных определяет особенности их раннего эмбриогенеза.
однослойная, с полостью внутри
Амфибластула:
многослойная, полость смещена к крыше,
у вегетативного полюса клетки крупнее
6.1.2.2. Дифференцировка - составная часть гаструляции
перемещениям клеток и
их продолжающейся дифференцировке.
2. а) Дифференцировка проявляется в том, что
клетки приобретают всё большие биохимические и морфологические различия друг от друга,
а возможности их дальнейшего развития всё сужаются.
б) Например, клетки энтодермы далее могут превращаться только в эпителиальные клетки органов желудочно-кишечного тракта.
6.1.2.3. Факторы, вызывающие гаструляцию
6.2. Образование осевых зачатков органов
6.2.1. Производные зародышевых листков
1. Из материала трёх зародышевых листков (эктодермы, мезодермы, энтодермы) на следующем этапе эмбриогенеза формируются осевые зачатки органов.
2. Рассмотрим этот процесс на примере птиц.
У других животных, в т.ч. млекопитающих, данная стадия эмбриогенеза проходит сходным образом.
6.2.1.1. Производные мезодермы
б) Формирующие её клетки мигрируют из эпибласта через первичный бугорок.
в ) А. Хорда - непарная осевая структура.
Б. Одна из её функций - установление оси тела .
б) Их ключевая особенность состоит в том, что они представляют собой сегменты - компактные скопления мезодермальных клеток.
в) А. Всего вдоль осевой линии образуется 10-11 пар сомитов (у человека - 44 пары) .
Б. Но появляются все пары сомитов не одновременно, а последовательно - в направлении от передней части зародыша к задней.
миотом (4) (среднюю часть) - зачаток поперечно-полосатых мышц,
б) Расслаивается на 2 листка:
париетальный листок (7,А), прилегающий к эктодерме,
висцеральный листок (7,Б), прилегающий к энтодерме .
Б. Кроме того, висцеральный листок - зачаток миокарда и эпикарда.
б) Находится как внутри зародыша, так и вне его (внезародышевая мезенхима).
в) Из мезенхимы образуются
сосуды (в т.ч. аорта (10) ),
ткани внутренней среды организма -все виды соединительных тканей (в т.ч. скелетные и хрящевые),
кроветворная ткань и сама кровь,
6.2.1.2. Производные эктодермы
I. Нервная трубка и ганглиозные пластинки
желобок смыкается в нервную трубку (11) (непарный зачаток),
б) Является зачатком
эпидермиса кожи и его производных ,
эпителия начального и конечного отделов желудочно-кишечного тракта,
эпителия некоторых других органов.
6.2.1.3. Энтодерма
Б. Поэтому энтодерма называется кишечной .
а) Рассмотренная стадия эмбриогенеза часто в литературе называется нейруляцией , а сам зародыш - нейрулой .
Эти названия берут за основу один из важнейших процессов - образование нервной трубки.
б) Тем не менее, правильней использовать более общее название - " стадия образования осевых зачатков органов ".
6.2.2. Просмотр препаратов
6.2.2.1. Препараты зародыша курицы
I. Вид сверху
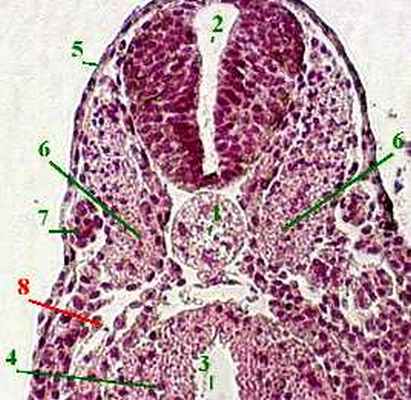
1. По оси зародыша - нервный желобок (1).
2. а) По краям от него зародыш значительно уплотнён -
благодаря наличию под эктодермой "мезодермальных крыльев" (2 ) .
б) А. Передняя часть мезодермы сегментирована:
II. Поперечный срез
1. Эктодерма (1 ) и её производная - нервная трубка (2) :
состоят из высоких цилиндрических клеток, расположенных в один слой.
в) Также видны нефрогонотомы, или сегментные ножки (8) .
г) Хорошо развит спланхнотом , расщеплённый на
п ариетальный (5,А) и
висцеральный (5,Б) листки ,
между которыми находится целомическая полость (6) .
6.2.2.2. Препараты зародышей амфибий
1. Гаструла амфибий (п. 6.1.1.2) заметно отличается от гаструлы птиц (п. 6.1.1.3) и млекопитающих (п. 6.1.1.4).
2. Тем не менее, и здесь образуются те же самые осевые зачатки органов.
I. Относительно ранняя стадия гаструляции
б) Как видно, в месте расположения нервной трубки поверхность зародыша сильно приподнята.
2. а) В центре - обширная полость первичной кишки (3 ) .
В этом - отличие от птиц и млекопитающих , у которых энтодерма - однослойная и ещё на замкнута на данной стадии в первичную кишку.
II. Более поздняя стадия гаструляции
3. а) Остальные структуры имелись и прежде.
б) Это
хорда (1),
нервная трубка (2),
полость первичной кишки (3),
многослойная энтодерма (4),
кожная энтодерма (5).
6.3. Образование внезародышевых органов
и отделение от них тела зародыша
Внезародышевые органы иначе называются провизорными (временными).
Ещё одно название - зародышевые оболочки .
6.3.1. Внезародышевые органы у рыб и птиц:
схема образования
У рыб из внезародышевых органов имеется только желточный мешок,
а у пресмыкающихся и птиц - 4 образования:
желточный мешок,
амнион,
серозная оболочка и
аллантоис.
6.3.1.1. Желточный мешок
б) Затем внезародышевые части энтодермы (1) и висцерального листка мезодермы (2) начинают
обрастать желток, формир уя желточный мешок (13) .
б) Поэтому четыре листка :
эктодерма,
энтодерма,
париетальный и
висцеральный листки мезодермы -
В его стенке впервые образуются клетки крови ( из мезодермального зачатка).
Здесь впервые обнаруживаются предшественники половых клеток (которые затем перемещаются в закладки гонад).
Содержимое желточного мешка - источник строительного материала для зародыша ( трофическая функция).
6.3.1.2. Амнион и серозная оболочка
эктодерма (4) и
париетальный листок (3) мезодермы -
б) Одновременно происходит обрастание теми же листками внутренней поверхности скорлупы яйца . - Формируется серозная оболочка.
эктодермальный листок обращён внутрь (к зародышу),
а париетальный листок мезодермы - кнаружи.
в) А. Данная оболочка отграничивает амниотическую полость,
в которой пространство между оболочкой и зародышем заполнено жидкостью.
б) В ней расположение листков обратное:
эктодермальный листок обращён кнаружи ,
а париетальный листок мезодермы - внутри.
6.3.1.3. Аллантоис
б) Он образован теми же двумя листками ( внезародышевой локализации), что и желточный мешок -
висцеральным листком мезодермы и
энтодермой
6.3.2. Внезародышевые органы на препаратах
6.3.2.1. Зародыш рыб
хорда (1),
нервная трубка (2),
с омиты (3),
первичная кишка (4).
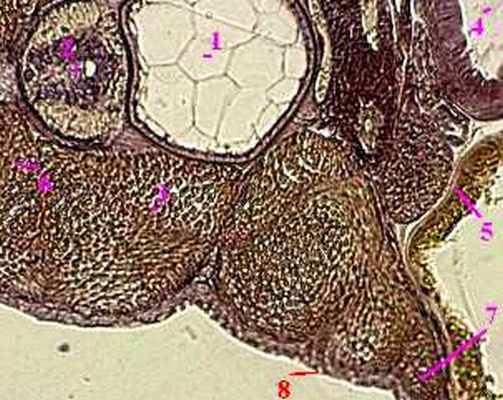
энтодермой (5 ) и
висцеральным листком мезодермы (6) .
б) Н а периферии зародыша к этим листкам присоединяются
париетальный листок (7) мезодермы и
эктодерма (8).
6.3.2.2. Зародыши птиц
I. Образование складок
а) производные эктодермы -
кожная эктодерма (1) и
нервная трубка (2);
б) производные мезодермы -
сомиты , которые уже разделяются на плотный дерматом (4), срединно расположенный миотом (5) и прилегающий к хорде склеротом (6),
спланхнотомы с висцеральным (8) и париетальным (9) листками и целомической полостью (10) между ними,
мезенхима , из которой, в частности, образованы аорта (11) и клетки крови (12).
зародыш приподнимается над желтком (последний на препарате отсутствует),
и внезародышевые части 4-х листков образуют туловищные складки (14).
в) А. Двое из этих листков -
внезародышевая энтодерма (15 ) и
висцеральный листок внезародышевой мезодермы (16) -
формируют стенку желточного мешка.
париетальный листок внезародышевой мезодермы (18) и
внезародышевая эктодерма (19) -
II. Смыкание складок
1. а) З десь вид на будущая презумптивная первичная кишка (1) .
б) Е ё стенка образована
кишечной энтодермой (2) и
висцеральным листком спланхнотома (3).
3. С мыкание амниотических складок, состоящих из
париетальных листков спланхнотома (6) и
внезародышевой эктодерм ы (7 ) ,
6.3.3. Внезародышевые органы у млекопитающих
1. При эмбриональном развитии млекопитающих тоже образуются желточный мешок, амнион и аллантоис.
2. а) Вместо же серозной оболочки формируется хорион - ворсинчатая оболочка.
б) После внедрения хориона в слизистую оболочку матки с одной его стороны ворсинки сильно разрастаются и, глубоко проникая в слизистую, образуют вместе с ней новый орган - плаценту.
6.3.3.1. Способ образования
Амнион и желточный мешок образуются у высших млекопитающих и человека иначе, чем у птиц.
а) Он появляется в результате расщепления эпибласта на два листка -
зародышевый эпибласт (1) и
амниотическую эктодерму (2).
б) Её клетки выселяются из зародышевого щитка в полость бластоцисты и постепенно обрастают все имеющиеся поверхности -
амниотического пузырька (5,А),
желточного мешка (5,Б ) ,
а также самого трофобласта (5,В).
б) Он связан с хорионом с помощью амниотической ножки (8) из внезародышевой мезенхимы.
6.3.3.2. Состав стенок внезародышевых органов
Несмотрия на иной способ формирования, все перечисленные структуры имеют ту же природу, что и у птиц. -
| Стенка желточного мешка | В незародышев ая энтодерм а (гипобласт) - изнутри; мез е нхима - снаружи. |
| Стенка амниотического пузыря | Внезародышев ая эктодерм а - изнутри, мезенхим а - снаружи. |
| Стенка хориона | Трофобласт (тоже внезародышев ая эктодерм а ) - снаружи, мезенхим а - изнутри . |
1. а) Далее мы обратимся к строению основных типов тканей организма (общая гистология) и строению органов и система (частная гистология).
б) При этом будем касаться и эмбрионального происхождения соответствующих тканей и органов, что расширит приведённые в этом разделе сведения.
2. После чего (в последнем разделе курса) более детально рассмотрим вопросы раннего эмбриогенеза человека.
Cyt_embr_rus
Рис. 2.1.1. Типы яйцеклеток по количеству и расположению желтка (верхняя часть рисунка) и соответствующие им типы дробления (нижняя часть рисунка): А – олиго- и изолецитальная яйцеклетка млекопитающих и человека – дробление полное неравномерное; Б – поли- и телолецитальная яйцеклетка птицы – дробление неполное неравномерное; В – поли- и центролецитальная яйцеклетка амфибии – дробление неполное неравномерное.
1 – ядро яйцеклетки; 2 – желточные включения; 3 – анимальный полюс; 4 – вегетативный полюс; 5 – бластомеры.
Рис. 2.1.2. Схема дробления зародыша млекопитающих:
А – стадия темных и светлых бластомеров; Б, В – стадия обрастания темных бластомеров светлыми; Г – стадия морулы; Д, Е – стадия бластоцисты.
1 – темные бластомеры; 2 – светлые бластомеры; 3 – полость бластоцисты; 4 – оболочка оплодотворения; 5 – эмбриобласт; 6 – трофобласт.
К НАЧАЛУ ГЛАВЫЦитология и общая эмбриология
Рис. 2.1.3. Схема образования первичной полоски и миграции клеток при формировании зародышевых листков: А – образование первичной полоски и первичного узелка; Б – образование нотохордомезодермального зачатка; В – образование нотохорды (сагиттальный разрез); Г – образование мезодермы (сагиттальный разрез). Сплошные стрелочки обозначают направление перемещения материала в составе наружного листка (до погружения); пунктирные – в составе среднего листка (после погружения).
1 – эпибласт; 2 – материал будущей нервной пластинки; 3 – материал будущей нотохорды; 4 – первичный узелок; 5 – первичная ямка; 6 – первичная полоска; 7 – первичная борозда; 8 – нотохорда; 9 – мезодерма; 10 – энтодерма.
Рис. 2.1.4. Зародыш курицы 16-ти часов инкубации. Тотальный препарат: А – ув. Ч 2; Б – ув. Ч 20.
1 – зародышевый диск; 2 – зародышевый щиток; 3 – первичная полоска; 4 – первичный узелок;
5 – мезодермальный пласт (крылья).
К НАЧАЛУ ГЛАВЫЭмбриогенез птиц и млекопитающих
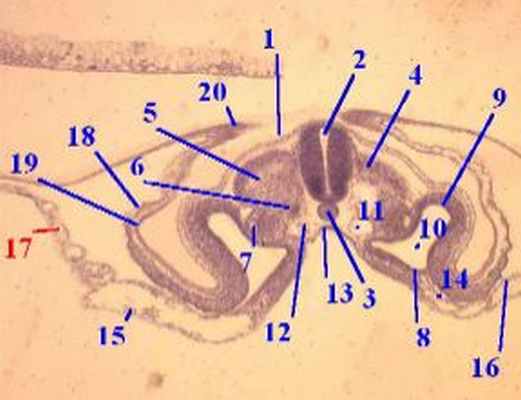
Рис. 2.1.5. Сегментация мезодермы у куриного зародыша (30 ча- сов инкубации, 8 пар сомитов).
1 – нервные валики; 2 – нервный желобок; 3 – нотохорда; 4 – сомиты; 5 – париетальный и висцеральный листки спланхнотома; 6 – передняя кишка; 7 – кишечный желобок (зачаток среднего отдела кишки); 8 – кровяные островки; 9 – остаток первичной полоски.
Рис. 2.1.6. Зародыш курицы 36-ти часов инкубации. Тотальный препарат. Ув. Ч 20.
1 – зародышевый диск; 2 – зародышевый щиток; 3 – светлое поле;
11 4 – темное поле; 5 – край обрастания; 6 – нервная трубка; 7 – соми-
12 ты; 8 – мозговые пузыри; 9 – амниотическая складка; 10 – зачаток сердца; 11 – остаток первичной полоски; 12 – кровяные островки.
К НАЧАЛУ ГЛАВЫЦитология и общая эмбриология
Рис. 2.1.7. Поперечный разрез зародыша курицы 42 часов инкубации на стадии дифференцировки зародышевых листков и образования осевого комплекса зачатков. Гистологический препарат. Ув. Ч 100.
– эктодерма: 1 а – внезародышевая, 1 б – зародышевая; 2 – зародышевая
– нервная трубка; 4 – нотохорда; 5 – мезодерма; 6 – сомит; 7 – спланхнотом: 8
ный листок, 9 – париетальный листок; 10 – целом; 11 – нефрогонотом; 12 – дерматом; 13 – склеротом; 14 – миотом; 15 – зачаток сердца.
Рис. 2.1.8. Зародыш курицы на стадии образования туловищной и амниотической складок. Гистологический препарат. Ув. Ч 100.
1 – туловищная складка; 2 – стенка амниона; 3 – зародышевая эктодерма; 4 – нервная трубка; 5 – нотохорда; 6 – зародышевая энтодерма; 7 – внезародышевые экто-, мезо- и энтодерма; 8 – зачаток сердца; 9 – кровеносные сосуды.
К НАЧАЛУ ГЛАВЫЭмбриогенез птиц и млекопитающих
Ðèñ. 2.1.9. Ñ хема развития внезародыше-
вых органов у млекопитающих: А – про-
цесс обрастания полости бластоцисты
энтодермой и мезодермой; Б – образо-
вание замкнутого энтодермального пу-
зырька – выстилки желточного мешка;
В – начало образования амниотических
складок; Г – образование туловищных
складок – обособление тела зародыша от
внезародышевых частей; Д – смыкание
амниотических складок, начало развития
аллантоиса; Е – замкнутая амниотическая
полость, развитый аллантоис, развитие
1 – тело зародыша; 2 – первичная кишка;
3 – туловищная складка; 4 – амниотическая
складка; 5 – амниотическая оболочка; 6 – хо-
рион; 6а – гладкий хорион; 6б – ворсинчатый
хорион; 7 – стенка желточного мешка;
8 – аллантоис; 9 – трофобласт; 10 – эпибласт; 11 – внезародышевая мезодерма; 12 – внезародышевая энтодерма (гипобласт); 13 – нервная трубка; 14 – нотохорда.
К НАЧАЛУ ГЛАВЫЦитология и общая эмбриология
Для самоконтроля решите следующие тестовые задания:
ТЕСТОВОЕ ЗАДАНИЕ 1
Выберите один наиболее правильный ответ.
Во время дробления зародыша млекопитающих происходит увеличение количе-
за счет митоза. Укажите тип дробления данной зиготы.
ТЕСТОВОЕ ЗАДАНИЕ 2
В зародышевом материале, направленном на
с тонкой стенкой, который заполненный серозной жидкостью. При исследовании в све-
товом микроскопе видно, что стенка пузырька образована одним слоем
ток. Внутри, на одном из полюсов, находятся темные клетки. Назовите
А. Стадия образования темных и светлых бластомеров. Б. Стадия обрастания темных бластомеров светлыми. В. Стадия бластоцисты.
Г. Стадия морулы.
Д. Стадия первичного узелка.
ТЕСТОВОЕ ЗАДАНИЕ 3 Выберите один наиболее правильный ответ.
В эмбриональном материале, направленном на исследование, выявляется зародыш, в котором определяются два слоя - эпибласт и гипобласт, образовавшиеся путем деламинации. Назовите период эмбрионального развития.
Б. 1-я фаза гаструляции. В. Дробление.
Г. 2-я фаза гаструляции. Д. Гисто- и органогенез.
ТЕСТОВОЕ ЗАДАНИЕ 5 Выберите один наиболее правильный ответ.
В эмбриональном материале, направленном на исследование, в эпибласте зародыша выявляются первичная полоска, первичный узелок. Назовите период эмбрионального развития.
Б. 1-я фаза гаструляции. В. Дробление.
Г. 2-я фаза гаструляции. Д. Гисто- и органогенез.
К НАЧАЛУ ГЛАВЫТрофобласт у млекопитающих обеспечивает:
1) процесс имплантации;
2) образование серозной жидкости бластоцисты;
3) обмен веществ с материнском организмом;
4) развитие плаценты.
Во время гаструляции происходит:
1) деление клеток;
2) перемещение клеток;
3) дифференцировка клеток, образование зародышевых листков;
4) индукционные взаимодействия.
ТЕСТОВОЕ ЗАДАНИЕ 6 Выберите один наиболее правильный ответ.
В эмбриональном материале, направленном на исследование, определяется зародыш на начальном этапе нейруляции. Что является условием начала этого процесса?
А. Дифференцировка. Б. Миграция клеток.
В. Эмбриональная индукция. Г. Детерминация.
В процессе дифференцировки из кишечной энтодермы образуются:
1) эпителий печени;
2) покровный и железистый эпителий пищевода;
3) покровный и железистый эпителий желудка;
4) покровный эпителий анального отдела прямой кишки.
При экспериментальном повреждении у зародыша сегментных ножек наблюдается нарушение развития органов:
1) половой системы;
2) пищеварительной системы;
3) мочевой системы;
4) дыхательной системы.
Зародышевая мезодерма подразделяется на:
При разрушении средней зоны сомита нарушается развитие: 1) волокнистой соединительной ткани;
К НАЧАЛУ ГЛАВЫЦитология и общая эмбриология
3) поперечно-полосатой сердечной мышечной ткани;
4) поперечно-полосатой скелетной мышечной ткани. Сомиты представлены:
Нарушение процессов дифферецировки спланхнотома может сопровож-
гладкой мышечной ткани;
2) поперечно-полосатой сердечной мышечной ткани;
форменных элементов крови;
ТЕСТОВОЕ ЗАДАНИЕ 8
Выберите один наиболее правильный ответ.
А. Мезотелий, соединительная ткань
производные, эпителий анального отдела прямой кишки
полости, пищевода, органов дыхания
и костные ткани
ТЕСТОВОЕ ЗАДАНИЕ 9
Выберите один наиболее правильный ответ.
При микроскопическом исследовании зародышевого материала в препарате определяется оболочка, которая образована внезародышевыми эктодермой и мезодермой. Укажите функцию данного органа.
А. Трофическая Б. Продукция околоплодных вод В. Кроветворная Г. Экскреторная Д. Эндокринная
1. Имеет ворсинки.
2. Предохраняет зародыш от действия гравитационного поля.
3. Является первым органом кроветворения.
4. Выполняет трофическую, выделительную, дыхательную и эндокринную функции.
5. Образуется из внезародышевой энтодермы и мезодермы.
6. Является местом образования гонобластов.
А) Хорион, Б) Амнион, В) Оба, Г) Ни один.
К НАЧАЛУ ГЛАВЫЭмбриогенез человека I
2.2. ЭМБРИОГЕНЕЗ ЧЕЛОВЕКА. ОПЛОДОТВОРЕНИЕ. ДРОБЛЕНИЕ. ПЕРВАЯ ФАЗА ГАСТРУЛЯЦИИ
АКТУАЛЬНОСТЬ ТЕМЫ: Изучение эмбриологии человека является научным фундаментом для таких медицинских дисциплин как акушерство, гинекология, педиатрия. На сегодняшний день эмбриология стала не только источником инфомации для понимания принципов дисморформогенеза и тератогенеза, механизмов развития органов и тканей, которые повторяются при их репарации (восстановление после повреждения), но и источником стволовых клеток. Выделенные впервые в 1990 году эмбриональные стволовые клетки стали центром научных и социальных дискуссий, поскольку их использование может стать основой восстановления утраченных в результате заболеваний функций таких жизненно важных органов как сердце, печень, почки, головной и спинной мозг. На знаниях закономерностей реализации и молекулярной регуляции ранних этапов эмбриогенеза основывается внедрение в клиническую практику экстракорпорального оплодотворения. Лечение мужского и женского бесплодия, невынашивание беременности, профилактика нарушений развития эмбриона и плода, пренатальная диагностика и мониторинг течения беременности невозможны без понимания процессов, которые происходят при внутриутробном развитии.
ЦЕЛЬ ОБУЧЕНИЯ (общая): Уметь определять периоды эмбриогенеза человека, их сущность, интерпретировать механизмы и значение процессов оплодотворения, дробления и ранней гаструляции для понимания основ бесплодия и гибели зародыша, которые изучаются на последующих этапах обучения.
Для этого необходимо уметь (конкретные цели):
Выявлять мужские и женские гаметы, интерпретировать особенности их образо-
вания, отличия от соматических клеток.
Трактовать суть периодов эмбриогенеза человека. Определять стадии оплодотво-
рения, трактовать их сущность и механизмы.
Выявлять особенности, локализацию и этапы дробления у человека, трактовать
его сущность и механизмы.
условия, проявления и механизмы имплантации бластоцисты,
их значение для развития зародыша человека.
Трактовать суть и морфологические проявления ранней фазы гаструляции чело-
внезародышевые органы человека, особенности их образования
ИНСТРУКЦИЯ ДЛЯ САМОСТОЯТЕЛЬНОЙ ПОДГОТОВКИ В развитии зародыша человека различают три перида: начальный (1-я неделя
Эмбриогенез тесно сопряжен с прогенезом — гаметогенезом. Половые клетки являются высокодифференцированными, способными к взаимодействию во время оплодотворения и образованию одноклеточного организма – зиготы. Гаметогенез (развитие половых клеток) определяет качество гамет и включает:
1) образование первичных половых клеток – гонобластов, их накопление в стенке
желточного мешка, миграцию
в зачатки гонад;
2) размножение предшественниц
половых клеток путем митоза;
К НАЧАЛУ ГЛАВЫЦитология и общая эмбриология
изменчивость за счет кроссинговера, который происходит в профазу 1-го мейоза;
образование в половых клетках гаплоидного набора хромосом в результате мейоза;
дифференцировку половых клеток (характерна для сперматогенеза).
Участниками процесса оплодотворения у человека являются вторичный овоцит
(из которого образуется яйцеклетка) и многочисленные спермии. Обратите внимание, что одним из современных методов диагностики мужского бесплодия является оценка спермограммы, которая учитывает количество, подвижность и строение сперматозоидов. Во время изучения этого вопроса обратите внимание на особенности строения
образуется комплексом Голь-
джи и содержит важные ферменты для осуществления процесса оплодотворения. Подвижность спермиев связана с наличием и структурной организацией жгутика, для которого обеспечивают продукцию энергии митохондрии, расположенные в промежуточ- ной части (рис. 2.2.1).
Яйцеклетка человека, как и у млекопитающих – олиго- и изолецитальная, поскольку содержит мало желтка, равномерно распределенного в цитоплазме. При анализе овоцита обратите внимание на особенности строения его цитоплазмы, богатой органеллами, наличие в ней кортикальных гранул. Уясните понятие «оотипическая детерминация». Снаружи овоцит окружен прозрачной оболочкой , которая содержит уникальные гликопротеины – ZP1, ZP2, ZP3, а также фолликулярным эпителием . Последний образован несколькими слоями клеток, связанных между собой плотными контактами и десмосомами, их отростки проникают через прозрачную оболочку и формируют контакты с овоцитом, обеспечиваю его трофику, защиту и регуляцию (рис. 2.2.2).
Следует уяснить, что оплодотворение обеспечивает взимодействие отцовской и материнской гамет, передачу их генетической информации с формированием одноклеточного зародыша – зиготы . Этому процессу предшествует завершение 2-го мейоти- ческого деления с формированием яйцеклетки , в которой активизируется обмен веществ. В результате оплодотворения восстанавливается диплоидный набор хромосом, характерный для человека.
Во время оплодотворения различают три фазы: дистантного , контактного взаимодействий è слияния гамет (сингамия). Эти фазы реализуются благодаря различным процессам и регуляторам (рис. 2.2.3). Дистантное взаимодействие обеспечивает прибли-
такой последовательности: 1)
под влиянием секрета эпителиальных кле-
ток слизистой оболочки матки – осуществляется путем модификации гликокаликса спермиев и деградации «оболочки», которая покрывает рецепторы спермиев. Данный процесс сопровождается также активацией метаболизма в сперматозоидах, что способствует повышению скорости биения жгутика; 2) хемотаксис – направленное движение спермиев в сторону овоцита, который выделяет специфические вещества – ãèíî-
гамоны ; 3) реотаксис – движение сперматозоидов
против движения слизи и ресничек
на поверхности эндометрия и слизистой оболочки
Контактное взаимодействие между овоцитом и сперматозоидами происходит путем преодоления барьеров овоцита – слоя фолликулярного эпителия и прозрачной оболочки. Этот сложный процесс обеспечивается за счет акросомальной реакции, во время которой происходит слияние плазмолеммы головки сперматозоида с наружной акросомальной мембраной и последующим освобождением ферментов – гиалуронидазы и акрозина (рис. 2.2.4 Б), которые разрушают барьеры на пути сперматозоида. Гиалуронидаза расщепляет связи между фолликулярными клетками, окружающими овоцит. Акрозин способствует пенетрации блестящей оболочки — защитной оболочки яйцевой клетки (рис. 2.2.5).
Ïðè слиянии гамет наблюдается проникновение сперматозоида в перивителлино-
ГАСТРУЛЯЦИЯ, ГИСТО- И ОРГАНОГЕНЕЗ ЧЕЛОВЕКА
Гаструляция, гИсто- и органогенез человека. Внезародышевые органы. Критические периоды развития.
Пользуясь лекциями (на web -странице кафедр ы есть презентац ии и текст лекц и й), учебниками , до полнительной л и тературо й и другими источниками , студент ы должны п о дготовит ь так ие теоретич еские вопросы :
1. Определение гаструляции. Фазы и способы гаструляции зародыша человека.
2. Первая фаза гаструляции у человека. Способы образования зародышевых листков и развития провизорных органов.
3. Вторая фаза гаструляции. Источник образования зачаточной мезодермы и осевого комплекса зачатков органов.
4. Дифференциация зародышевых листков: экто-, энто- и мезодермы.
5. Гисто- и органогенез.
6. Понятие о стволовых клетках и диффероне.
7. Критические периоды эмбриогенеза.
8. Источника развития амниона, желточного мешка, алантоиса и хориона.
9. Образование, строение и функциональное значение желточного мешка.
10. Развитие, строение и функция амниона.
11. Алантоис, его развитие и значение.
12. Образование и строение пуповины.
13. Формирование хориона, первичные и вторичные ворсинки, их морфологические особенности.
14. Плацента человека, его морфологические особенности и значения.
15. Строение плодовой части плаценты.
16. Строение материнской части плаценты.
17. Структурные компоненты гемохориального (плацентарного) барьера.
18. Система “мать-плод”.
19. Понятие о критических периодах развития.
Гаструляция
1. Формирование многослойного зародыша.
2. Следующая после дробления стадия ембр и огенезу.
3. Тип гаструляц ии определяется типом яйцеклетки и типом дробления зиготи.
4. Есть ранняя гаструляц и я и поздняя.
Морфогенез - формирование пространственной организации организма и его частей. Многоклеточный организм развивается из оплодотворенной яйцеклетки ( зиготы ) не только путем увеличения количества клеток (пролиферация), но и увеличением массы зародыша (рост). Одновременно определяется судьба образующихся клеток, то есть клетка « выбирает » один из многих возможных путей развития . Этот процесс известен как « детерминация». Детерминированные клетки специализируются (путем дифференцировки), т.е. приобретают определенную структуру и оказываются способными выполнять конкретную функцию. Одновременно осуществляется морфогенез - клетки координированным образом формируют органы и архитектуру пространственной организации тела. Формирование пространственной архитектуры зародыша (организма) и его частей (органов) осуществляется при реализации следующих морфогенетических процессов: направленная миграция клеток (в том числе направленный рост частей клеток, например, отростков нервных клеток), гибель клеток. Таким образом, пролиферация, рост, детерминация, дифференцировка, морфогенез, миграция клеток и их гибель - важные события (морфогенетические процессы) при развитии многоклеточного организма.
Все многообразие клеток организма ( различные типы клеток) развивается из одной единственной (зиготы). Говоря формально, все клетки организма - клоны, родоначальником которых является оплодотворенная яйцеклетка. Клон , по определению, совокупность идентичных клеток, развившихся из единой клонообразующей клетки. Однако для развивающегося организма это не так: существует множество различных клеточных типов, возникающие в результате различной - дифференциальной - активности генов. В чем же причина разнообразия при начальной идентичности? По сути, у нас один ответ - эндогенная программа многообразия посредством ядерного генома зиготы. Итак, генетическая программа - источник разнообразия клеток. Такое утверждение верно для одноклеточного зародыша. И это же утверждение становится неочевидным для многоклеточного зародыша, хотя бы в силу их различной взаимной позиции (например, на поверхности зародыша или в глубине, в левой или правой части зародыша, в верхней или нижней части). Следовательно, для многоклеточного зародыша обязаны существовать внешние по отношению к конкретной клетке зародыша факторы, определяющие дифференциальную активность генов в разных клетках зародыша. Рассмотрение такой возможности определяет гипотеза позиционной информации в терминах «морфогенетическое поле», «левое - правое», «гомеобокс», «гомеозисние гены». Иными словами, возникновение разнообразия различных клеток в многоклеточном зародыше смещается в плоскость межклеточных взаимодействий («индукция», «индукционные взаимодействия», «факторы индукции») . Иными словами , дифференциальная активность генов определяется позицией конкретной клетки и реализуется с помощью различного типа межклеточных взаимодействий.
В эмбриогенезе появляются различия между клетками: возникают различные клеточные типы. Конкретные типы клеток образуют ткани. Из клеток различной тканевой принадлежности формируются органы. Определение пути развития тотипотентных клеток концептуса и полипотентных клеток концептуса, эмбриона, плода при внутриутробном развитии и организма в постнатальном онтогенезе происходит в ходе детерминации - процесса в результате которого "компетентная клеточная система выбирает один из многих возможных путей развития" . Такая потенциальная возможность развиваться в разных направлениях обозначается как проспективная потенция .
Дифференциация - внешнее выражение детерминации . В ходе специализации конкретного клеточного типа (дифферон) формируются различные фенотипы клеток. Результат дифференцирования - специализированная клетка конкретной морфологи , выполняющая определенную функцию (состояние терминальной дифференцировки). По мере дифференцировки постепенно ограничиваются потенции клеток развиваться в разных направлениях. Дифференциация необратима и осуществляется только в одном направлении - от менее дифференцированной к более дифференцированной структуре. При дифференцировании клетки экспрессируют строго определенную при детерминации часть генома: транскрибируют специфические РНК и синтезируют специфические белки, что и определяет морфологические и функциональные признаки специализации клеток. Итак , различия между клетками, обладают одинаковым набором генов, определяет дифференциальная активность генов.
Гены оплодотворенной яйцеклетки репрессированы. Начало развития сопровождается дерепрессия определенных групп генов ( в первую очередь генов , контролирующих пролиферацию и общий метаболизм клетки). Первые тканеспецифические гены активируются на стадии гаструляции. Позже , когда происходит координированная составления сложных структур (органогенез), включаются другие гены. Различные клеточные типы экспрессируют разные гены.
Ограничение проспективных потенций (комитирование). По мере дифференцировки постепенно ограничиваются потенции клеток развиваться в разных направлениях. Например, клетки несегментированное мезодермы имеют потенции к дифференцировке в различных направлениях и образования миогенного, хондрогенного, остеогенной и других направлений дифференцировки. Клетки миотома сомитов детерминированы к развитию только в одном направлении , а именно к образованию миогенного клеточного типа (поперечнополосатая мышечная ткань скелетного типа).
Пролиферация и дифференцировка. Существенная черта дифференцировки заключается в том, что дифференцирование обычно наступает после пролиферации клеток. Клетки , которые быстро размножаются, как правило , является малодифференцированными (например, клетки базального слоя эпителия кожи или мезенхимные клетки). Напротив, высокодифференцированные клетки, как правило, теряют способность к пролиферации ( например , эритроциты и нейроны).
Дифференциация является необратимой и осуществляется только в одном направлении - от менее дифференцированной к более дифференцированной структуры. Для нетрансформированных клеток , а также для стволовых клеток в условиях in vitro не следует применять термин дедиференцирования (как правило, речь при этом идет о границах клеточного типа и его пластичности).
Морфогенез осуществляется при реализации различных морфогенетических процессов: рост , межклеточные взаимодействия , индукция , направленная миграция клеток , направленный рост частей клеток ( например , отростков нервных клеток) , гибель клеток.
Рост - увеличение массы и , как правило, линейных размеров за счет увеличения количества клеток, морфо - функциональных единиц органов, самих органов, систем органов и т.д. Увеличение массы без клеточных делений наблюдается при гипертрофии клеток в нормальных (например, гипертрофия хрящевых клеток, гипертрофия миометрия при беременности) и патологических условиях. В организме вырабатываются многочисленные гуморальные факторы, стимулирующие рост, а также пролиферацию различных клеточных типов, - факторы роста .
Межклеточные взаимодействия и индукция . Специализацию клеток и образование новых структур направляют межклеточные взаимодействия и индукция. Природу клеточных взаимодействий в морфогенезе объясняет концепция позиционной информации .
Общий план тела определяется очень рано. Позже , в течение всего периода формирования органа или целого организма, детали морфогенеза уточняются с помощью сигналов позиционной информации. Согласно концепции позиционной информации , клетка « знает » свое местоположение в координатной системе зачатка органа и дифференцируется в соответствии с этим положением. Позиционную информацию клетка получает от других клеток. Более того, клетка достигает состояния терминальной дифференцировки только при условии своевременного получения ею серии последовательных сигналов позиционной информации. Зона, в пределах которой эффективно действуют сигналы позиционной информации, называется морфогенетическим полем . В течение ряда последующих клеточных делений клетки морфогенетического поля «помнят» о своем первоначальном назначении. Постоянная активность гомеозисних генов определяет в клетке память о позиционной информации.
В 1969 г. Льюис Вольперт предложил модель позиционной информации (« французского флага ») , согласно которой положения клетки в морфогенетическом поле определяется в соответствии с существующей системой координат. Модель предполагает наличие специальных позиционных химических сигналов , или морфогенное , продуцируемых клетками - источниками и формируют во внеклеточном пространстве диффузные градиенты. Клетки- мишени регистрируют градиенты морфогенности интерпретируют их для определения своей локализации в морфогенетических поле. Существуют и другие представления о механизме , определяющем положение клеток в морфогенетических поле. Так, согласно модели последовательной передачи сигнала от клетки к клетке (sequential cell context model), предполагается последовательная индукция различных сигналов в прочих группах клеток с последующим их совместным действием на экспрессию специфических генов в других прилагают клетках.
Индукционные взаимодействия . В органогенезе - координированной сборке различных тканевых структур - важное значение имеют индукционные взаимодействия между эмбриональными зачатками. В ходе индукции клетки одного зачатка (источник) влияют на клетки другого зачатка (мишень). Источник инструктирует мишень к дифференцировке в конкретную структуру или позволяет дифференцировку. Возникшая структура придает индуцирующий влияние на другую мишень, и появляется новая структура и т.д. Эмбриогенез - сплошная череда индукционных взаимодействий.
Первичная эмбриональная индукция - влияние хордомезодермы на дорсальную эктодерму , результат - образование зачатка нервной системы.
Закладка конечностей. В результате индукционного влияния клеток латеральной мезодермы на эктодерму возникает локальное утолщение эктодермы , вместе со скоплением мезодермальный клеток формирует зачаток конечности.
Нейруляция яйцеклетки. Сегментация мезодермы.
1. Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных животных. – М.: Наука, 1964. – Т. 1. – 433 с.
2. Гудвин Б. Аналитическая физиология клеток и развивающихся организмов / пер. с англ. – М.: Мир, 1978. – 330 с.
4. Иванов П.П. Первичная и вторичная метамерия тела // Журнал общей биологии. – 1944. – № 5. – С. 61-95.
6. Лебедкин С.И., Герке П.Я. Основы теоретической анатомии человека. – Рига: изд-во АН Латвийской ССР, 1963. – 356 с.
7. Ливанов Н.А. Пути эволюции животного мира // Ученые записки Казанск. гос. ун-та. – 1945. – Т. 105. – Кн. 3. – Вып. 9. – С. 114.
8. Петренко В.М. Морфогенез корней нижней полой вены в эмбриогенезе человека // Морфология. – 1998. – Т. 114. – № 5. – С. 56-59.
9. Петренко В.М. О морфогенезе брыжеечных лимфатических узлов у новорожденных белой крысы // Успехи современного естествознания. – 2011. – № 9. – С. 49-52.
10. Петренко В.М. Квазисегментарное устройство тела человека // Международный журнал прикладных и фундаментальных исследований. – 2014. – № 8. – Ч. 1. – С. 59-62.
13. Станек И. Эмбриология человека / пер. со словац. – Братислава: изд-во Словац. АН «Веда», 1977. – 440 с.
Введение
Метамерия и сегментация тела играют важную роль в жизнедеятельности животных [6,10,11,14]. Метамерия состоит в повторении внутренних органов по длинной оси тела животного, от нее не всегда отличают внешнюю сегментацию [14]. Н.А. Ливанов заметил, что «…метамерия представляет явление сложное, которое в известной мере – во «внешней» метамерии, может быть связано с движением животного и с выработкой локомоторных придатков, параподий, а во всей полноте будет раскрыто… при анализе «внутренней» метамерии тела» [7]. По мнению Л.А. Зенкевича, одним из важных условий возникновения метамерии тела является уплотнение наружных покровов тела, что усиливает защиту животного, но ограничивает подвижность тела. Противоречие снимает образование эластичных межсегментных соединений [14]. А. Ланг считал, что тело животного разделяется на метамеры, благодаря появлению тех или иных органов, метамерное расположение которых полезно животному: в связи с его змеевидными движениями (придатки для ползания) или с необходимостью увеличить плодовитость (повторное расположение гонад), или с потребностью улучшить условия кровообращения (повторное расположение анастомозов между главными продольными сосудами). Следствием метамерии отдельных органов является метамерия мезодермы и разделение ее на сомиты уже в эмбриогенезе [14]. П.П. Иванов [4] разделил метамеры на ларвальные (головные, челюстные), связанные с фукциями нащупывания и захвата субстрата, зарывания в грунт, и постларвальные (туловищные). В отличие от П.П. Иванова, В.Н. Беклемишев [1] считал метамерию результатом приспособления животного не к локомоции, а к размножению. Поэтому ларвальные сегменты лишены полового аппарата. Согласно П.П. Иванову [4], постларвальная сегментация тела у вторичноротых животных возникает вследствие дополнительной, но не полной сегментации заднего или туловищного отдела тела путем повторного образования некоторых органов, например гонад, или путем метамерных сужений и расширений продольных органов, например образования печеночных выступов и перехватов между ними в кишечнике. Под это определение подпадает морфогенез центральных краниальных брыжеечных лимфоузлов [9].
Механизм формирования сомитов в эмбриогенезе остается неизвестным, его изучение не выходит за чисто описательный уровень, хотя и проведено уже немало различных экспериментов [3,5]. Разные авторы указывают на влияние хорды и нервной трубки, гензеновского узелка и боковой пластинки мезодермы. A.M. Turing (1952) математически показал, что распространение какого-либо метаболита на протяжении аксиальной мезодермы носит волнообразный характер, из чего некоторые эмбриологи сделали вывод о существовании каких-то веществ, обладающих морфогенетическим давлением. Однако результаты ряда опытов на зародышах амфибий отрицают существование каких-либо влияний, передающихся от головного конца дорсальной мезодермы к ее хвостовому концу, регулирующих ее сегментацию. Возможно сегментация аксиальной мезодермы не зависит от какого-либо регуляторного сигнала, исходящего от головного участка, а может начаться в любой точке по оси зародыша независимо от участков, лежащих кпереди от нее. При этом не исключена возможность, что в каждом изолированном отрезке оси зародыша инициируется новый кранио-каудальный регулирующий механизм [3].
Цель исследования: рассмотреть возможную роль общеизвестных процессов эмбрионального органогенеза в образовании сомитов.
Материал и методы исследования
Работа проведена на 30 эмбрионах человека 4-8 нед. (5-30 мм теменно-копчиковой длины). После фиксации в жидкости Буэна материал заливали в парафин с последующим изготовлением серийных срезов толщиной 5-7 мкм в трех основных анатомических плоскостях. Срезы окрашивали гематоксилином и эозином, смесью Маллори, альциановым и толуидиновым синими при рН = 1,0-5,0 под ферментативным контролем.
Результаты исследования их обсуждение
В еще несегментированной аксиальной мезодерме эмбриона человека определяются сгущения клеток. Быстро растущие кластеры все более темных клеток разделяются постепенно утолщающимися прослойками более светлых клеток: цепь сомитов удлиняется, последовательно присоединяя новые звенья, в которых клетки приобретают радиальную ориентацию. У амфибий они описаны как розетки, в их пределах аффинитет между мезодермальными клетками выше, чем между соседними розетками [3]. Образование сомитов происходит с конца 3-й нед и до начала 6-й нед [12]. В эти сроки наблюдается скручивание тела эмбриона вокруг продольной оси в процессе интенсивного каудального удлинения эмбриона, плавающего вокруг сужающегося зародышевого ствола, в окружении уплотняющихся оболочек. Эмбрион до фиксации освобождают от стягивающих при фиксации оболочек, чтобы получить его недеформированные срезы. На 6-й нед заметно накопление протеогликанов в зачатках позвонков. Охрящевление туловищного скелета ясно выражено на 7-й нед, когда прекращается кручение эмбриона, которое, видимо, с 4-5 нед тормозят сердце и печень, интенсивно растущие в каудальном направлении.
Наиболее интенсивно сомитообразование происходит у эмбриона человека 4-й нед [12]. Обособление закладки любого органа в эмбриогенезе сопряжено с формированием его автономного сосудистого русла. В литературе [12,13] сообщается лишь, что дорсальные ветви дорсальной аорты идут по обе стороны от нервной трубки, в рыхлую мезенхиму между сомитами – межсегментарные артерии. Им после рождения соответствуют межреберные и поясничные артерии. В конце 4-й нед новообразование поясничных сомитов наблюдается дорсокаудальнее бифуркации аорты (рис. 1-3).
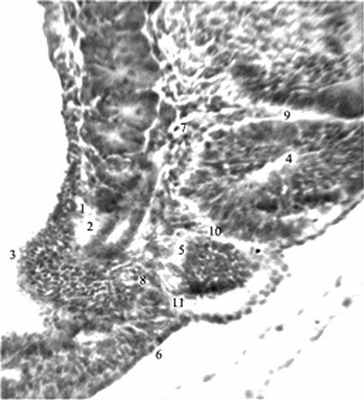
Рис. 1. Эмбрион человека 5 мм длины (4 недель), сагиттальный срез: 1,2 – вещество и проток мезонефроса; 3 – метанефрогенная бластема; 4,5 – поясничные сомиты на разных этапах развития; 6 – недифференцированная
дорсальная мезодерма; 7,8 – протоковая вена
и ее корни; 9-11 – межсомитные сосуды.
Гематоксилин и эозин. Ув. 200.
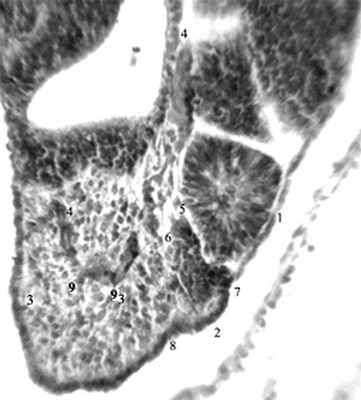
Рис. 2. Эмбрион человека 5 мм длины (4 недель), сагиттальный срез: 1-2 –сомиты на разных этапах морфогенеза; 3-3 – несегментированная аксиальная мезодерма; 4-4 – протоковая вена; 5,6 – межсомитные сосуды; 7,8 – межсомитные борозды; 9 – ветви протоковой вены,
внедряющиеся в аксиальную мезодерму.
Гематоксилин и эозин. Ув. 200.
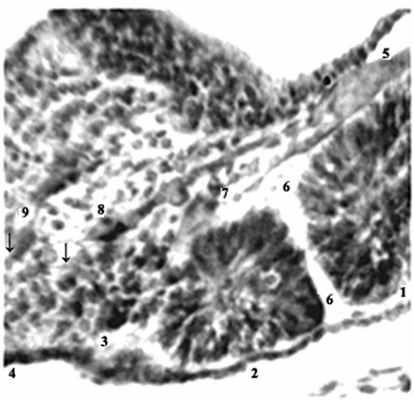
Рис. 3. Эмбрион человека 5 мм длины (4 недель), сагиттальный срез: 1,2 – сомиты; 3,4 – сегментация недифференцированной дорсальной мезодермы (новообразование сомитов);
5 – протоковая вена; 6-9 – межсомитные сосуды. Стрелки указывают на светлые клинья
мезенхимы и микрососуды, внедряющиеся
в тяж дорсальной мезодермы.
Гематоксилин и эозин. Ув. 300.
В этой области, между протоком мезонефроса (вентрально) и сомитами (дорсально) проходит протоковая вена, вентрокаудальный приток посткардинальной вены [8]. Вслед за протоком и веществом мезонефроса протоковая вена поворачивает вентромедиально. От ее изгиба отходят ветви, вентральные – к метанефрогенной бластеме, дорсальные – к сомитам и несегментированной мезодерме. Межсегментарные сосуды вместе с рыхлой мезенхимой внедряются в толщу тяжа дорсальной мезодермы, в промежутки между еще только намечающимися сомитами.
Изложенное позволяет предположить, что в пространственно-временном аспекте сомитообразование сопряжено с кручением «мягкого» тела эмбриона в процессе его интенсивного каудального удлинения. Кручение является особым видом сдвига, приводящему к смещению слоев в неупругих телах вплоть до «течения» материала, как в жидкостях. Рыхлая мезенхима пластична, очень легко деформируется с усилением циркуляции межклеточной жидкости. Все это должно стимулировать рост протокапилляров, лишенных базальной мембраны. Кручение тела эмбриона сопровождается сходной деформацией продольных тяжей дорсальной мезодермы. На ее протяжении возникают участки сужения, где определяются выступы рыхлой мезенхимы с кровеносными микрососудами. Их стенки испытывают повышенное наружное давление (касательное напряжение кручения ?), что индуцирует их уплотнение и утолщение, начиная с эндотелия, – развитие его базальной мембраны и адвентициальной оболочки. Резистентность сосуду придает и кровяное давление (жидкость несжимаема). В результате возникает клин растущей жесткости, который: 1) «отсекает» от тяжа дорсальной мезодермы сомит; 2) фиксирует тяж мезодермы в этом месте, что приводит к каудальному распространению волны кручения и сегментирования дорсальной мезодермы. Такое (ступенчатое, пошаговое) ее взаимодействие с кровеносными сосудами, когда последние внедряются в мезодермальный тяж, «нарезая» сомиты, можно сравнить с зубчатой передачей, а с учетом кручения – с винтообразной передачей. Размер ее «шага» (→ сомита), т.е. сегментирования мезодермы, определяют, очевидно, мезодермальные клетки: тип (способ их группировки) и пределы (объем сомита) их роста адекватны свойствам их белков, информация о первичной структуре которых записана в геноме клеток. Сосудисто-мезенхимные перегородки аксиальной мезодермы растут между формирующимися розетками, т.е. в «слабые» места дифференцирующегося тяжа мезодермы – места снижающегося аффинитета ее клеток. Хотя в этих же участках определяются «перехваты» тела эмбриона, обусловленные кручением – генетическая детерминация и эпигенетическая регуляция процесса ?
Заключение
Кровеносные сосуды участвуют в морфогенезе сомитов как (раз)делители их зачатков в условиях продольного растяжения и кручения тела эмбриона с его мягким скелетом. Предлагаемая двухволновая модель сегментирования осевой мезодермы подкрепляет мое же предположение [10] о важной роли аорты в становлении квазисегментарного устройства тела человека, начиная с его эмбрионального периода развития. Волны дифференциации осевой мезодермы: 1) детерминации (или распространения компетентности [2]) – продольное растяжение мезодермы при удлинении эмбриона с напряжением адгезии клеток индуцирует их пролиферацию и сгущение, что стимулирует рост микрососудов; 2) регуляции процесса (как в виртуальной модели периодического морфогенеза E.С.Zeeman [2]) – кручение обусловливает поперечную перетяжку мезодермы, что облегчает ее разделение на сомиты сосудисто-мезенхимными клиньями. Причем вторая волна дифференциации (~ кручение) инициируется ее первой волной (удлинение тела эмбриона → краниокаудальный градиент напряжения мезодермы). Моя гипотеза корреллирует с разными предположениями о механике становлении метамерии животного в эволюции: 1) А.Ланга – связь с локомоцией и размещением сосудов; 2) Б. Гатчека и Э. Перрье – способность пролиферировать на заднем конце тела однородные небольшие участки [14].
Читайте также: