Половой хроматин. Тельца Барра
Добавил пользователь Алексей Ф. Обновлено: 27.01.2026
Половой хроматин. Тельца Барра
Х-хроматин (тельце Барра) представляет собой хромоцентр величиной около 1 мкм, красящийся всеми основными ядерными красителями более интенсивно, чем остальные хроматиновые структуры ядра. Фельген-положительная реакция свидетельствует о большой концентрации в нем ДНК.
Локализация Х-хроматина в ядре различна. В большинстве тканей он находится на внутренней поверхности ядерной оболочки и может иметь треугольную, плоско-выпуклую, трапециевидную, U-образную или гантелевидную форму. Иногда Х-хроматин имеет вид утолщения или зубца ядерной мембраны, соединенного с ядрышком тонкой хроматиновой нитью. В веретеновидных и палочковидных ядрах Х-хроматин располагается на одном из полюсов ядра.
Реже Х-хроматин располагается на ядрышке или в нуклеоплазме, при этой локализации он обладает сферической формой и трудно отличим от других хромоцентров, имеющих такой же размер, но неспецифических для пола. Поэтому в целях диагностики половой принадлежности клеток большинство исследователей учитывают хромоцентры, расположенные только у ядерной мембраны.
Положение Х-хроматина может меняться в одних и тех же клетках в зависимости от их функционального состояния, а также в процессе онтогенеза.
Х-хроматин обнаружен в клетках различных тканей у многих млекопитающих; у грызунов (хомяки, крысы, мыши, морские свинки) хроматиновые структуры ядер представлены большим количеством хромоцентров, затрудняющих выявление Х-хроматина. У человека половые различия в строении ядер установлены практически во всех тканях и органах.
Происхождение Х-хроматина. В процессе клеточного цикла хромосомы претерпевают закономерные преобразования, которые состоят в спирализации и деспирализации хромосом и их репродукции. В интерфазе максимально деспирализованные хромосомы образуют ядро с относительно гомогенным содержимым. Репродукция (синтез ДНК) хромосом происходит только в деспирализованном состоянии в период S-интерфазы.
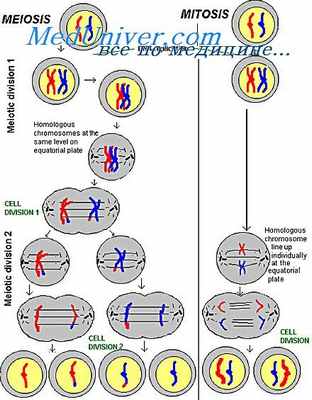
Спирализуясь, хромосомы вступают в профазу митоза и достигают наибольшей спирализации в метафазе митоза и мейоза. При этом они обладают минимальной специфической активностью. Вместе с тем установлено, что хромосомы всегда неравномерно спирализованы по длине и разделяются на гетерохроматические и эухроматические районы. Морфологически эти районы различаются по интенсивности окраски и структурной организации.
Эухроматические районы в интерфазном ядре деспирализуются, в то время как гетерохроматические имеют тенденцию оставаться в спирализованном компактном состоянии в виде хромоцентров с высоким содержанием ДНК. Спирализованность гетерохроматических районов сопровождается неактивным состоянием генов, содержащихся в них. Эта особенность свойственна также некоторым эухроматнческим районам с высоко функционально дифференцированными генами. Будучи спирализованными в стадии интерфазного ядра, эухроматические районы становятся также генетически неактивными.
Гетерохроматизация — универсальный механизм генетической инактивации хромосомных участков независимо от того, относятся ли они к гетерохроматическим или к эухроматнческим районам. Следовательно, хромоцентры, обнаруживаемые в интерфазном ядре, могут быть образованы как гетерохроматином, так и эухроматином. Одним из таких хромоцентров является Х-хроматин.
Еще Ваrr и Bertram высказали предположение о связи феномена Х-хроматина с Х-хромосомами. С тех пор Х-хромосомная природа Х-хроматина подтверждена и уточнена данными многочисленных исследователей.
Х-хроматин образован одной из Х-хромосом женской клетки, находящейся в гетерохроматизированном состоянии. Будучи спирализованной, эта хромосома генетически неактивна. В разных клетках сомы у женских особей, по принципу случайности, Х-хроматнн образует Х-хромосома, полученная либо от отца, либо от матери. Следовательно, клетки женского организма мозаичны по функции Х-хромосомы: в одних активна отцовская, в других материнская хромосома. Образование полового хроматина в женских клетках обусловлено генетически.
Это подтверждается тем, что в раннем периоде развития эмбриона человека, когда по виду гонад еще нельзя определить пол, яйцевые оболочки зародыша мужского пола не имеют Х-хроматина, несмотря на воздействие гормонов матери. У зародыша женского пола Х-хроматин появляется на 16-й день развития, когда в эмбрионе насчитывается 2500—5000 клеток.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Вопрос 9. Половой хроматин.
В 1949 г. Барр и Бертрам в ядрах нейронов самок кошек обнаружили интенсивно, окрашенную глыбку хроматина. Она имеет треугольную форму и прилежит к внутренней мембране ядра. Далее было установлено, что только одна из двух Х-хромосом в соматических клетках женских особей функционально активна, другая конденсируется (факультативный гетерохроматин), и в интерфазном ядре образует Х-хроматин, или тельце Барра. Этот процесс имеет случайный характер в эмбриональном периоде. В норме у женщин (46, ХХ)– одно тельце Барра, у мужчин (46, ХУ) оно отсутствует. Изменение числа Х-хромосом ведёт к изменению числа телец Барра.
Количество телец Барра всегда на единицу меньше, чем количество половых Х-хромосом в кариотипе. Сумма п Х – 1.
Этот феномен получил название правила СТЮАРТА. Например, если кариотип 47, ХХХ, то три Х-хромосом минус одна, равняется два тельца Барра. Присутствие полового хроматина у мужчин, а также наличие дополнительных или отсутствие телец Барра у женщин характерно для нарушений в системе половых хромосом.
Увеличение числа У-хромосм приводит к увеличению флуоресцирующих телец в интерфазных ядрах, названных У-хроматином.
Разработан экспресс-метод определения полового хроматина в соскобе буккального эпителия слизистой щеки. Материал соскоба, полученный с помощью шпателя, переносится на предметное стекло и окрашивается 1% -ным раствором ацетоарсеина, накрывается покровным стеклом и изучается с помощью светового микроскопа.
Используют определение полового хроматина для:
• Своевременное определение пола в решении вопросов, о наследственных сцепленных с полом заболеваний,
• Экспресс-диагностики хромосомных болезней, связанных с нарушением комплекса половых хромосом это с. Клайнфельтера 47, ХХУ. 48, ХХХУ,
Тернера 45, ХО,
Морриса 46, ХУ – женский фенотип,
Трипло – Х – суперженщина 47, ХХХ,
Дисомию по У-хромосоме 47, ХХУ.
+ Возможность выявить мозаицизм по половым хромосомам – гинандроморфизм – ХХУ/ХХ проявляются как, женские так и мужские признаки. Описаны самые разнообразные формы кариотипов ХУ/ХО, ХХ/ХО, ХХХ/ХО,ХХ/ХХХ, тройные мозаики ХО/ХХ/ХХХ.
• Определение пола в судебной медицине
• В онкологии для определения опухоли по половому хроматину и выбора правильного гормонального лечения
Вопрос 11. Метод генетики соматических клеток
Этот метод представляет собой культивирование, клонирование, гибридизацию и селекцию соматических клеток. КЛОНИРОВАНИЕ – получение потомков (большого числа клеток) от одной клетки за счет деления. Клонированные клетки используют для получения большого числа клеток для хромосомного анализа, для изучения особенностей обмена, для количественного учета мутаций, для доказательства гетерогенности клеточных популяций.
Селекцию в генетике соматических клеток применяют для отбора мутантов по резистентности, ауксотрофности. Отбор мутантов по резистентности основан на их выживании в присутствии какого – либо летального фактора. Отбор ауксотрофных клеток основан на их свойствах использовать для своего роста строго определенные вещества, не синтезируемые клеткой.
Путем гибридизации устанавливается локализация генов в хромосомах.
Гибридизация основана на слиянии совместно культивируемых клеток двух разных типов. Гибридные клетки или гетерокарионы, содержат ядра с хромосомными наборами двух разных видов, например человека и мыши, человека и крысы, человека и китайского хомячка.
Генотипы таких клеток находятся в состоянии дисбаланса и поэтому при клеточном делении гетерокарионы обычно теряют часть хромосом. В разных гибридных клетках утрачиваются хромосомы одного вида. В гибридных клетках «человек - мышь» постепенно исчезают хромосомы человека. Постепенная утрата хромосом человека может привести в конечном итоге к сохранению единственной хромосомы. Так сохранение 9 хромосомы человека в гибридной клетке «человек - мышь» позволило установить, что в ответ на введение вируса клетки этого клона начали вырабатывать интерферон. Поэтому установили, что ген ответственный за синтез интерферона расположен в 9 хромосоме.
Половой хроматин. Тельца Барра
1 ГБОУ ВПО «Тихоокеанский государственный медицинский университет» Министерства здравоохранения Российской Федерации
Возрастные функциональные особенности репродуктивной системы женщин находятся в тесной зависимости от изменений нейроэндокринного гомеостаза, которые, несомненно, регулируются определенными генами. Активность этих генов, в свою очередь, зависит от степени компактизации хромосом и связана с участками гетеро- и эухроматина. Инактивацию Х-хромосомы относят к одному из эпигенетических феноменов, которые рассматривает современная наука о наследуемых свойствах организма, не связанных с изменением собственно нуклеотидной последовательности, – эпигенетика. Число ядер, в которых присутствует гетерохроматин, зависит от степени пролиферации клеток в различных тканях, а также от гормонального статуса организма, влияния стресса, антибиотиков и глюкокортикоидов, физической активности женщины. Учитывая совершенно противоположные литературные данные по содержанию и изменчивости полового хроматина в ядрах клеток буккального эпителия, мы предприняли попытку определения телец Барра у женщин в возрастном аспекте. Доказано постепенное снижение количества полового хроматина в клетках буккального эпителия у женщин старших возрастных групп.
1. Олейник Е.А., Дюсенова А.А. Использование метода буккальных мазков в определении полового хроматина у женщин-спортсменок // Морфология. – 2006. – Т. 130, № 5. – С. 66.
2. Шуматова Т.А., Приходченко Н.Г., Оденбах Л.А., Ефремова И.В. Роль метилирования ДНК и состояния фолатного обмена в развитии патологических процессов в организме человека // Тихоокеанский мед. журн. – 2013. – № 4. – С. 39–43.
3. Anderson C.L., Brown C.J. Variability of X chromosome inactivation: effect on levels of TIMP1 RNA and role of DNA methylation // Hum Genet. – 2002. – Vol. 110. – P. 271–278.
4. Barr M.L., Bertram E.G. A morphological distinction between neurons of the male and female and the behaviour of the nucleolar satellite during accelerated nucleoprotein synthesis // Nature. – 1949. – Vol. 163. – P. 676.
5. Berletch J.B., Yang F., Xu J. et al. Genes that escape from X inactivation // Hum Genet. – 2011. – Vol. 130. – P. 237–245.
7. Bondy C.A. Care of girls and women with Turner syndrome: a guideline of the Turner Syndrome Study Group // J Clin Endocrinol Metab. – 2007. – Vol. 92. – P. 10–25.
8. Carrel L., Willard H.F. X-inactivation profile reveals extensive variability in X-linked gene expression in females // Nature. – 2005. – Vol. 434. – P. 400–404.
9. Deng X., Berletch J.B., Nguyen D.K., Disteche C.M. X chromosome regulation: diverse patterns in development, tissues and disease // Nat Rev Genet. – 2014. – Vol. 15. – P. 367–378.
10. Golovin DI, Zus’ BA. Sex chromatin in oncomorphology // Arkh Patol. – 1981. – Vol. 43(12). – P. 3–8.
11. Lessing D., Anguera M.C., Lee J.T. X chromosome inactivation and epigenetic responses to cellular reprogramming // Annu Rev Genomics Hum Genet. – 2013. – Vol. 14. – P.85–110.
12. Peeters S.B., Cotton A.M., Brown C.J. Variable escape from X-chromosome inactivation: identifying factors that tip the scales towards expression // BioEssays. – 2014. – Vol. 36. – P.746–756.
13. Simon M.D., Pinter S.F., Fang R. et al. High-resolution Xist binding maps reveal two-step spreading during X-chromosome inactivation // Nature. – 2013. – Vol. 504. – P. 465–469.
14. Splinter E., Wit E., Nora E.P. et al. The inactive X chromosome adopts a unique three-dimensional conformation that is dependent on Xist RNA // Genes Dev. – 2011. – Vol. 25. – P. 1371–1383.
15. Wu H., Luo J., Yu H. et al. Cellular resolution maps of x chromosome inactivation: implications for neural development, function, and disease // Neuron. – 2014. Vol. 81. – P. 103–119.
Определение половой принадлежности, а также своевременное выявление аномалий полового развития, наследование болезней, сцепленных с полом, представляет огромный интерес не только для врачей разных специальностей, но востребовано и в спорте, судебной медицине [2].
С самого раннего периода развития гистологии и цитологии в ядре были замечены интенсивно окрашивающиеся структуры. Их назвали прохромосомами, хромоцентрами, и считалось, что в этих местах хроматин проявляет положительный гетеропикноз. В настоящее время данные структуры определяются как гетерохроматин, в отличие от слабо окрашивающегося эухроматина. Впервые эти названия были предложены в 1933 году Гейтцем [11]. В 1937 году Гейтлер сообщал о половом ядерном диморфизме у насекомых, наблюдая двойное тело гетерохроматина у женских особей и одинарное тело у мужских особей, предполагая, что открытый им механизм может найти применение при определении пола [11]. Впоследствии, в 1949 году, в журнале «Nature» была опубликована работа Бара и Бертрама, в которой ученые описали морфологические различия в нейронах самок и самцов [4]. Открытие особых образований – глыбок гетерохроматина в интерфазных ядрах соматических клеток (телец Барра) позволило в дальнейшем использовать половой хроматин для решения некоторых диагностических вопросов медицинской генетики. Тельце Барра соответствует одной из двух Х-хромосом в клетках особей женского пола. В клетках у мужчин половой хроматин практически отсутствует, хотя некоторые авторы указывают на обнаружение 2–3 % подобных глыбок [5].
Первоначально для определения полового хроматина применяли биопсию кожи, но вскоре был описан метод исследования мазка слизистой оболочки ротовой полости. Исследование эпителия полости рта – это попытка избежать хирургической биопсии, заменить ее простым методом взятия материала и тем самым расширить возможность клинического применения метода. Так, у женщин-спортсменок проводят половой контроль перед олимпийскими играми [1]. Спортсменки, чей генетический пол не соответствует габитусу, имеющие морфологические признаки мужского соматотипа, имеют преимущество перед женщинами-спортсменками с женским кариотипом [1]. Безусловно, данный метод нашел применение и в медицине. Скрининговый тест, не требующий глобальных материальных затрат, применяется для быстрой диагностики количественных нарушений в половых хромосомах, таких как синдром Шерешевского – Тернера, Клайнфельтера, полисомии Х [7].
В настоящее время все больше и больше авторов придерживаются мнения о том, что половой хроматин все-таки изменяется. Интересны и труднообъяснимы наблюдения Sohval и Casselman, которые установили, что величина и окраска полового хроматина может изменяться под влиянием антибиотиков [9]. Другие исследователи показывают, что в ротовой полости происходит снижение полового хроматина при некоторых видах аллергии, а также при стрессе [12, 15]. У индивидуумов женского пола половой хроматин в ротовой полости снижается при приеме глюкокортикоидов [9, 15]. В опухолях различных локализаций обнаружена четкая обратно пропорциональная зависимость между уменьшением числа ядер с половым хроматином и увеличением митотического индекса [10]. Так, при раке молочной железы значительная часть клеток показала потерю полового хроматина в значительной части клеток опухолей. Положительная корреляция между количеством полового хроматина в клетках и частотой метастазирования опухоли была описана в некоторых работах [6].
Учитывая совершенно противоположные литературные данные по содержанию и изменчивости полового хроматина в ядрах клеток буккального эпителия от 90 % [8] до 28 % [1], мы предприняли попытку определения телец Барра у женщин в онтогенезе, используя различные методы окраски клеток.
Цель исследования – изучить динамику полового гетерохроматина в различные периоды онтогенеза женщины.
Материал и методы исследования
Материалом для настоящего исследования послужили клетки буккального эпителия, полученные при обследовании 100 женщин трех возрастных категорий: I – 18–20 лет, II – 38–55 лет, III – 56–75 лет. Соскоб эпителия производился с внутренней поверхности щеки. Ротовая полость предварительно прополаскивалась 2 % содовым раствором, что несколько уменьшало примесь слизи и микробов к исследуемому материалу. Эпителий слизистой полости рта снимался закругленным стеклом с намеченного места слизистой, выпяченной при надавливании пальцем снаружи. Соскоб слизистой после равномерного распределения по стеклу немедленно фиксировался в смеси равных частей спирта и эфира в течение 24 часов. Препараты окрашивались несколькими способами:
1) крезил-виолетом или по методу Папаниколау;
3) окраской по Фельгену, при фиксации материала в смеси Дэвидсона.
Микроскопирование осуществлялось с использованием светового микроскопа с иммерсионной системой, увеличение х1000 (Микроскоп Carl Zeiss Axioscope A1, камера для документирования AxioCam ICc5). Для более достоверной картины подсчет количества телец Барра проводился в 100 ядрах клеток буккального эпителия.
Результаты исследования и их обсуждение
Тельце Барра имеет вид маленькой темной массы овальной формы, примыкающей к внутренней поверхности ядерной оболочки (рисунок). Диаметр такого тельца равен примерно 1 мкм, поэтому его легко увидеть с помощью иммерсионного объектива. Надо заметить, что обнаружить во всех исследуемых ядрах данные структуры нам не удалось.
Иногда ядра клеток здоровых мужчин содержат дискретные массы конденсированного хроматина, которые примыкают к ядерной оболочке и выглядят как тельца Барра. Итак, тельце Барра нельзя увидеть в каждом ядре, в котором оно имеется, и поскольку в ядрах, в которых этих телец нет, иногда можно видеть похожие на них дискретные массы хроматина, заключение о наличии или отсутствии телец Барра нельзя делать на основании беглого осмотра отдельных клеток.
Содержание полового гетерохроматина в I группе (молодые женщины) составило 35,7 ± 3,1 % клеток, во II (женщины среднего возраста) – 33,2 ± 2,5 % клеток, в III (женщины старшей возрастной категории) – 25,75 ± 1,7 % клеток. Следовательно, с возрастом количество Х-полового хроматина у женщин в интерфазных ядрах буккального эпителия уменьшается.
Все процессы, протекающие в организме человека, контролируются генетическим аппаратом клеток. Но механизм специфической активации определенных генов в отдельно взятой клетке или клеточной популяции остается не изученным до конца. Исследование структур интерфазного ядра и специфических изменений в нем дает возможность в какой-то мере судить о супрессии или репрессии генов в зависимости от состояния организма. Возрастные функциональные особенности репродуктивной системы женщин находятся в тесной зависимости от изменений нейроэндокринного гомеостаза, которые регулируются определенными генами. Активность этих генов, в свою очередь, зависит от степени компактизации хромосом и связана с участками гетеро- и эухроматина [5, 9].
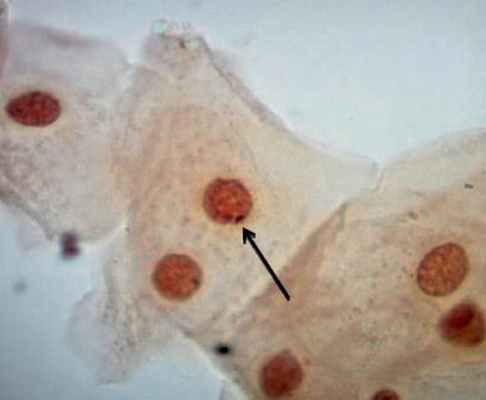
Тельце Барра в ядре клетки буккального эпителия женщины. Окраска ацетоорсеином, 1000х
Существование полов, а особенно то обстоятельство, что пол животного задается различиями по половым хромосомам, ставит перед системами регуляции экспрессии генов проблему: если гены, сцепленные с Х-хромосомой, будут экспрессироваться с разными интенсивностями у особей обоих полов, то количество продуктов экспрессии в клетках самки будет в 2 раза больше, чем в клетках самца. Чтобы избежать такой ситуации, существует явление компенсации доз генов [5, 9]. Суть явления состоит в выравнивании интенсивности экспрессии генов, расположенных на Х-хромосоме, между полами. У млекопитающих этот механизм представлен полной инактивацией одной из двух Х-хромосом в женском организме, в результате чего у самок активна только одна Х-хромосома, что эквивалентно ситуации с самцами. Мишенью для регуляции служит вся хромосома в целом, т.е. затрагиваются все промоторы на хромосоме. Хромосома переходит в состояние гетерохроматина [2, 3].
Возможный механизм инактивации Х-хромосомы связан с локусом Xic, где содержится ген, названный Xist. Ген Xist служит матрицей для синтеза Xist РНК (X inactive specific transcript), в которой нет открытых рамок считывания (она не является кодирующей). РНК Xist «обволакивает» данную Х-хромосому, тем самым ее инактивируя. Xist РНК синтезируется на обеих Х-хромосомах, но после инактивации такую РНК дает только неактивная Х-хромосома (где ее и можно встретить). Интенсивность же транскрипции Xist остается прежней, следовательно, перемена состояния хромосомы зависит не от нее, а от каких-то посттранскрипционных событий [11, 14].
Содержимое локуса Xic оказывает свое действие в цис-манере и несет в себе информацию, достаточную для того, чтобы «подсчитать» Х-хромосомы и инактивировать их все, кроме одной (правило n – 1). Правило n – 1 предполагает, что стабилизация РНК Xist происходит по умолчанию и что некий блокирующий механизм предотвращает такую стабилизацию лишь на одной из Х-хромосом, которой предстоит остаться активной. Это означает, что хотя наличие Xic необходимо (и достаточно), чтобы хромосома оказалась инактивирована, для установления активного состояния Х-хромосомы могут быть необходимы продукты других локусов [14].
Инактивация распространяется от Xic по всей Х-хромосоме. Если Xic присутствует в хромосоме-химере, возникшей в результате транслокации между Х-хромосомой и аутосомой, инактивация распространяется и на аутосомные участки (хотя этот эффект не всегда отличается полнотой). У нормальных самок Х-хромосом, конечно же, две, но в тех редких случаях, когда неправильное расхождение хромосом приводит к генотипу с тремя или более Х-хромосомами, активной останется лишь одна из них [13]. Это обстоятельство наводит на общую модель, в соответствии с которой с одной и только одной из Х-хромосом происходит некое особенное событие, охраняющее ее от воздействия инактивирующего механизма, который тем временем применяется по отношению ко всем остальным Х-хромосомам клетки. Для активности же Х-хромосомы необходимо «погасить» на ней экспрессию Xist. Сайленсинг Xist можно предотвратить делецией гена ДНК-метилтрансферазы; скорее всего, эффект обусловлен тем, что метилирование промотора Xist необходимо для прекращения его транскрипции [13]. Таким образом, метилирование ДНК в геноме является специфической формой клеточной памяти (эпигенетическая память), которая играет ключевую роль в развитии благодаря специфическому кодированию генной экспрессии в разных клетках. Несмотря на единый геном клетки организма имеют разные эпигеномы, что обеспечивает дифференциальную экспрессию, разные клеточные фенотипы и функции [2].
Заключение
В самом начале изучения полового хроматина считалось, что процент клеток, содержащих тельца Барра, является постоянным и не изменяется ни под влиянием возраста, ни под воздействием других внутренних или внешних факторов. Тем самым диагностика пола на основании оценки полового хроматина должна быть надежной. Эта точка зрения была основана на многочисленных наблюдениях [1, 7, 15]. Возможно, неизменность полового хроматина можно было бы связать с постоянной пропорцией клеток в организме, содержащих и не содержащих половой хроматин. Это означает, что каждая ткань имеет свой процент содержания полового хроматина, чему пока нет доказательств.
Удивительно низкие результаты получаются с эпителием ротовой полости, несмотря на то, что исследуемым материалом служат мазки, т.е. целые клетки. Число ядер, в которых присутствует гетерохроматин, зависит от степени пролиферации клеток в различных тканях, в частности в клетках буккального эпителия, а также от гормонального статуса организма. Можно предположить, что угасание функции половых желез посредством определенных факторов приводит к частичной деконденсации факультативного хроматина в некоторых клетках с целью усиления экспрессии генов Х-хромосомы для поддержания уровня гормонов в женском организме.
Рецензенты:
Матвеева Н.Ю., д.м.н., профессор, зав. кафедрой гистологии, эмбриологии и цитологии, ГБОУ ВПО «Тихоокеанский государственный медицинский университет» Министерства здравоохранения РФ, г. Владивосток;
Калиниченко С.Г., д.м.н., ст. научный сотрудник, ЦНИЛ ГБОУ ВПО «Тихоокеанский государственный медицинский университет» Министерства здравоохранения РФ, г. Владивосток.
Тельце Барра
Тельце Барра (X-половой хроматин) — свёрнутая в пло́тную (гетерохроматиновую) структуру неактивная X-хромосома, наблюдаемая в интерфазных ядрах соматических клеток самок плацентарных млекопитающих, включая человека. Хорошо прокрашивается осно́вными красителями [1] .
Из двух X-хромосом генома любая в начале эмбрионального развития может инактивироваться, выбор осуществляется случайно. У мыши исключением являются клетки зародышевых оболочек, также образующихся из ткани зародыша, в которых инактивируется исключительно отцовская X-хромосома [2] .
Таким образом, у самки млекопитающего, гетерозиготной по какому-либо признаку, определяемому геном X-хромосомы, в разных клетках работают разные аллели этого гена (мозаицизм). Классическим видимым примером такого мозаицизма является окраска черепаховых кошек — в половине клеток активна X-хромосома с «рыжим», а в половине — с «чёрным» аллелем гена, участвующего в формировании меланина. Коты черепаховой окраски встречаются крайне редко и имеют две X-хромосомы (анеуплоидия) [3] .
У людей и животных с анеуплоидией, имеющих в геноме 3 и более X-хромосом (см., напр., синдром Клайнфельтера), число телец Барра в ядре соматической клетки на единицу меньше числа X-хромосом.
Источники
- ↑ Биологический энциклопедический словарь. М., «Советская энциклопедия», 1986.
- ↑ М.Сингер, П.Берг. Гены и геномы. М., «Мир», 1998. (Том 2, стр.146)
- ↑Мозаицизм, черепаховые коты и генетически невозможные котята
- Генетика
- Цитология
- Половой диморфизм
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Тельце Барра" в других словарях:
тельце Барра — тельце Барра. См. половой хроматин. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
тельце барра — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ТЕЛЬЦЕ БАРРА – во всех соматических клетках генетически женского организма одна из Х хромосом инактивирована и известна как половой хроматин … Общая эмбриология: Терминологический словарь
Тельце Барра — интенсивно красящаяся структура, наблюдающаяся в ядрах разных типов клеток у самок млекопитающих. Представляет собой сильно спирализованную и поэтому неактивную Х хромосому. Инактивация одной из Х хромосом происходит случайным образом … Словарь по психогенетике
Тельце Барра Хроматиновое (Barr Body) — см. Хроматин половой. Источник: Медицинский словарь … Медицинские термины
Барра тельце — (М. L. Barr, род. в 1908 г., канадский гистолог) см. Хроматин половой … Большой медицинский словарь
Инактивация Х-хромосомы — Черепаховая окраска самок кошек является видимым примером инактивации Х хромосомы. Черный и оранжевый аллели гена окраски меха располагаются на Х хромосоме. Окраска конкретного участка меха определяется тем, какой из аллелей гена активен в данном … Википедия
половой хроматин — тельце Барра Гетерохроматинизированная в процессе лайонизации и интенсивно прокрашиваемая при анализе интерфазных ядер Х хромосома самок млекопитающих; выявление П.х. лежит в основе экспресс метода (т.е. без культивирования клеток и получения… … Справочник технического переводчика
Половой хроматин — плотное окрашивающееся тельце, обнаруживаемое в недслящихся (интерфазных см. Интерфаза) ядрах клеток у гетерогаметных (имеющих Х и Y Половые хромосомы) животных и человека. П. х. подразделяют на Х хроматин, или тельце Барра (открыт в 1949 … Большая советская энциклопедия
Гетерохроматин — Ядро фибробласта женщины с гетерохроматином, помеченным флуоресцентным красителем. Стрелкой указано положение тельца Барра Гетерохроматин участки … Википедия
Пол — I совокупность генетически детерминированных признаков особи, определяющих ее роль в процессе размножения. Развитие признаков мужского (обозначают знакома Марса ♂ ) и женского (знак Венеры ♀ ) полов определяется хромосомными наборами (см.… … Медицинская энциклопедия
Половой хроматин (тельца Барра).
В 1949 году М. Барр и Ч. Бертрам в ядрах нейронов кошки обнаружили небольшие тельца размером 0,8 – 1,1 мкм, названные тельцами Барра. Оказалось, что тельца Барра присутствует в интерфазных ядрах соматических клеток самок млекопитающих и отсутствует у самцов.
Аналогичные структуры были обнаружены в ядрах полиморфноядерных лейкоцитов в виде «барабанных палочек» (drиmsticks). Впоследствии было установлено, что эти тельца представляют собой конденсированную Х- хромосому, которая претерпевает инактивацию еще в раннем эмбриогенезе У человека между числом телец полового хроматина и количеством половых хромосом существует прямая связь: число телец полового хроматина на единицу меньше числа Х-хромосом. У здоровой женщины в соматических клетках можно обнаружить одно тельце полового хроматина, для нее это норма, для мужчины – это аномалия. Только на основании этого анализа врач может заподозрить синдром Клайнфельтера. Если у женщины не окажется ядер с половым хроматином, следует предположить кариотип с ХО, а если 2, 3 и более телец, то возможны кариотипы с ХХХ, ХХХХ, ХХХХХ хромосомами. В полиморфноядерных лейкоцитах крови женщин обнаруживается 1-6% «барабанных палочек», у мужчин они в норме не встречаются. Определение полового хроматина ПХ используется в диагностических целях нарушений половых Х - хромосом.
Диагностическое значение полового хроматипаВвиду доступности и простоты метода, определение полового хроматина нашло широкое применение при обследовании больных с нарушениями половых хромосом, определении пола плода при пренатальной диагностике заболеваний, сцепленных с полом, в судебной медицине для определении половой принадлежности пятен крови или частей трупа, в онкологии для назначения целенаправленной гормонотерапии, при трансплантации органов и тканей в качестве своеобразной метки пола донора и реципиента.
Метод дерматоглифики. Метод предназначен для изучения рельефа кожи на пальцах, ладонях и подошвах стоп. В этих участках тела имеются эпидермальные выступы – гребни, которые образуют сложные узоры. Каждый человек обладает своим рисунком кожных узоров, что позволяет использовать метод для идентификации личности в криминалистике.
Кожные узоры исследуются в различных направлениях: дактилоскопия – подушечек пальцев, пальмоскопия – ладоней и плантоскопия – подошв стопы. Папиллярные линии на пальцевых подушечках образуют три типа: дуга, петля, завиток. Количественным показателем дерматоглифики является гребневый счет. Дуговой узор встречается с частотой 6%, петлевой около 60% и завитковый занимает среднее положение 34%.
У основания пальцев расположены пальцевые трирадиусы (а,в,с,d). Вблизи браслетной складки, отделяющей кисть от предплечья, располагается главный трирадиус ( t ), угол, образованный на соединении линий atd, в норме не превышающий 57 о . Установлено, что при хромосомных аномалиях угол atdсоставляет: при синдроме Дауна 81 0 , Патау 108 0 , синдроме Шерешевского-Тернера 66 0 , синдроме Клайнфельтера 42 0 . Метод может использоваться в клинической генетике, в качестве дополнительного для прогноза проявления болезней с наследственной предрасположенностью
Биохимические методы.Это большая группа методов используется для диагностики болезней обмена веществ, причиной которых является изменения в деятельности ферментов, а причиной изменений являются генные мутации. В основе биохимических методов лежит определение либо первичного продукта деятельности мутантного гена – мутантного белка, либо определение промежуточных продуктов обмена, образующихся под влиянием дефектных ферментов. Методы трудоемки для исполнения не могут быть использованы для масовых исследований.
В последние годы во многих странах разработтаны специальные программы, названные скриннинг-программы. Для их выполнения используются наиболее простые и доступные экспресс-диагностические методы. Популяционно – статистический метод.Метод предназначен для изучения распространения отдельных генов в популяциях человека, иначе изучение геногеографии наследственных болезнейНа основании накопленных данных в этом плане, выделяются группы генов категории концентрации генов: гены, имеющие универсальное распространение, к числу которых относится большинство известных (фенилкетонурии, дальтонизма,различные формы слабоумия) , гены, имеющие локальное распространение (серповидно-клеточной анемии, врожденного вывиха бедра).Популяционно-статистический метод позволяет рассчитать генетическую структуру популяции по частотам генов и носителей генотипов в различных группах населения. Для этих целей служит уравнение Харди-Вайнберга. Метод генетики соматических клеток. Гибридизация соматических клеток – метод слияния клеток двух разных типов, полученных от разных людей, а также клеток человека с клетками мыши, морской свинки, китайского хомячка, обезъяны и других органгизмов. При слиянии таких клеток образуется гетерокарион – гибридная клетка с содержанием в одной цитоплазме ядрер обоих типов. После митотического деления, из двуядерных гетерокарионов образуются одноядерные клетки. Они образуют синкарион –гибридную клетку с хромосомами обеих родительских клеток.Например гибридные клетки человек-мышь имеют 43 пары хромосом, 23 пары от человека и 20 пар от мыши. При дальнейшем размножении происходит постепенное исчезновение хромосом одного вида. Этот метод позволяет построить генетические карты хромосом человека.
Читайте также: