Позные и локомоторные рефлексы спинного мозга. Шагательные движения
Добавил пользователь Валентин П. Обновлено: 29.01.2026
Спинальные вегетативные рефлексы. Спинальный шок
На уровне спинного мозга осуществляются многие типы сегментарных вегетативных рефлексов, большинство которых обсуждаются в других статьях на сайте (просим вас пользоваться формой поиска выше). К ним относят:
(1) изменения сосудистого тонуса в результате локального нагревания кожи;
(2) потоотделение в результате локального нагревания поверхности тела;
(3) кишечно-кишечные рефлексы, контролирующие некоторые моторные функции кишечника;
(4) брюшинно-кишечные рефлексы, тормозящие моторную активность желудочно-кишечного тракта в ответ на раздражение брюшины;
(5) эвакуационные рефлексы для опорожнения полного мочевого пузыря или переполненной толстой кишки.
Кроме того, все сегментарные рефлексы могут иногда возбуждаться одновременно в форме так называемого массивного рефлекса, изложенного далее.
Участки поверхности тела, где появляется отраженная боль от разных внутренних органов Висцеральная и париетальная передача болевых импульсов от аппендикса
а) Массивный рефлекс. Иногда у спинального животного или человека активность спинного мозга чрезвычайно повышается, что сопровождается массивным импульсным разрядом в значительной его части. Обычно это происходит в связи с действием сильного болевого раздражения кожи или чрезмерного переполнения внутренних органов, например при перерастяжении мочевого пузыря или кишки. Независимо от типа стимула возникающий в результате рефлекс, называемый массивным рефлексом, вовлекает большую часть или даже весь спинной мозг.
Эффекты представляют собой:
(1) мощный сгибательный спазм значительной части скелетных мышц тела;
(2) опорожнение прямой кишки и мочевого пузыря;
(3) часто подъем артериального давления до максимальных величин, иногда до уровней систолического давления, значительно превышающих 200 мм рт. ст.;
(4) обильное потоотделение обширных областей поверхности тела.
Поскольку массивный рефлекс может продолжаться в течение нескольких минут, он, вероятно, является результатом активации большого числа реверберирующих контуров, что одновременно возбуждает большие области спинного мозга. Это похоже на механизм развития эпилептических судорог, связанный с реверберацией возбуждения, возникающей в головном мозге вместо спинного.
Пересечение спинного мозга и спинальный шок
Когда спинной мозг внезапно пересекается в верхней части шеи, сначала практически все функции спинного мозга, включая спинномозговые рефлексы, мгновенно подавляются, вплоть до полного их выключения. Эту реакцию называют спинальным шоком. Причиной такой реакции является то, что нормальная активность спинальных нейронов зависит в большой степени от постоянного тонического возбуждения спинного мозга под действием импульсов, поступающих к нему по нисходящим нервным волокнам от высших центров, особенно по ретикулоспинальным, вестибулоспинальным и кортикоспинальным трактам.
В течение нескольких часов или недель возбудимость спинальных нейронов постепенно восстанавливается. По-видимому, это общее естественное свойство нейронов повсюду в нервной системе, т.е. после потери источника облегчающих импульсов нейроны увеличивают собственную естественную степень возбудимости, чтобы компенсировать потерю хотя бы частично. Большинству неприматов, чтобы возвратить к норме возбудимость центров спинного мозга, требуется от нескольких часов до нескольких суток. Однако у человека восстановление часто задерживается на несколько недель, а иногда полного восстановления совсем не происходит. В других случаях, наоборот, происходит чрезмерное восстановление с результирующей повышенной возбудимостью некоторых или всех функций спинного мозга.
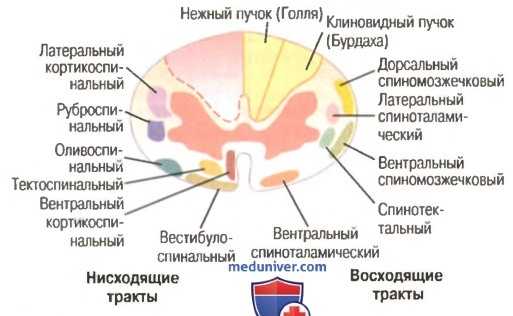
Поперечное сечение спинного мозга. Справа показаны главные восходящие тракты, слева — главные нисходящие тракты
Далее перечислены некоторые из спинальных функций, особенно страдающих во время или после спинального шока.
1. В начале спинального шока сразу и очень значительно падает артериальное давление, иногда опускаясь ниже 40 мм рт. ст., что свидетельствует о практически полной блокаде активности симпатической нервной системы. Давление обычно возвращается к норме в течение нескольких дней (даже у человека).
2. Все рефлексы скелетных мышц, интегрированные в спинном мозге, во время первых стадий шока блокируются. Животным для восстановления этих рефлексов до нормы требуется от нескольких часов до нескольких дней; людям — от 2 нед до нескольких месяцев. И у животных, и у человека некоторые рефлексы могут в итоге стать чрезмерно возбудимыми, особенно в тех случаях, когда на фоне пересечения основной части путей между головным и спинным мозгом некоторые облегчающие пути сохраняются. Первыми восстанавливаются рефлексы на растяжение, в дальнейшем постепенно восстанавливаются более сложные рефлексы в соответствующем порядке: сгибательные, антигравитационные позные и частично шагательные.
3. Рефлексы крестцового отдела спинного мозга, контролирующие опорожнение мочевого пузыря и прямой кишки, подавлены у человека в течение первых недель после пересечения спинного мозга, но в большинстве случаев они в итоге восстанавливаются.
Видео анатомия восходящих трактов спинного мозга (восходящих путей)
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Рефлексы спинного мозга
В данном разделе рассматриваются только соматические рефлексы (вегетативные рефлексы, см. раздел 3.7). Рефлексы спинного мозга достаточно просты. По форме это, в основном, сгибатель-ные и разгибательные рефлексы сегментарного характера. Надсегментарные рефлексы, наряду с сегментарными, осуществляются только с помощью шейного отдела.
А. Классификация соматических рефлексов спинного мозга. Все спинальные рефлексы можно объединить в две группы по следующим признакам. Во-первых, по рецепторам, раздражение которых вызывает рефлекс: а) проприорецептивные, б) висцероре-цептивные и в) кожные рефлексы. Последние являются защитными. Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висце-рорецептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины. Во-вторых, спинальные рефлексы целесообразно объединить по органам (эффекторам рефлекса): а) рефлексы конечностей, б) брюшные, в) органов таза. Рассмотрим рефлексы конечностей: сгибательные, разгибательные, ритмические и рефлексы позы.
Б. Сгибательные рефлексы - фазные и тонические.
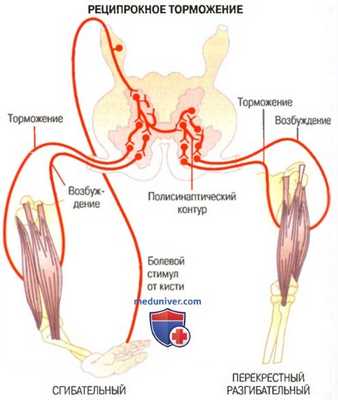
Фазные рефлексы - это однократное сгибание конечности при однократном раздражении рецепторов кожи или проприорецепторов. Одновременно с возбуждением мотонейронов мышц-сгибателей происходит реципрокное торможение мотонейронов мышц-разгибателей. Рефлексы, возникающие с рецепторов кожи, имеют защитное значение. Фазные рефлексы с проприорецепторов участвуют в формировании акта ходьбы.
Тонические сгибательные (как и разгибательные) рефлексы возникают при длительном растяжении мышц и возбуждении проприорецепторов, их главное назначение - поддержание позы. Тоническое сокращение скелетных мышц является фоновым для осуществления всех двигательных актов, осуществляемых с помощью фазных сокращений мышц.
В. Разгибательные рефлексы, как и сгибательные, бывают фазными и тоническими, возникают с проприорецепторов мышц-разгибателей, являются моносинаптическими.
Фазные рефлексы возникают на однократное раздражение мышечных рецепторов, например, при ударе по сухожилию четырехглавой мышцы ниже надколенной чашечки. При этом возникает коленный разгибательный рефлекс вследствие сокращения
 |
четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся - постсинаптическое ре-ципрокное торможение с помощью вставочных тормозных клеток Реншоу) - см. рис. 5.13. Рефлекторная дуга коленного рефлекса замыкается во втором - четвертом поясничных сегментах (Ц-Ь4). Фазные разгибательные рефлексы участвуют, как и сгибатель-ные, в формирование акта ходьбы.
Тонические разгибательные рефлексы представляют собой длительное сокращение мышц-разгибателей при длительном растяжении их сухожилий. Их роль - поддержание позы. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют также миотатическими.
Г. Рефлексы позы - перераспределение мышечного тонуса, возникающее при изменении положения тела или отдельных его частей. Рефлексы позы осуществляются с участием различных отделов ЦНС. На уровне спинного мозга замыкаются шейные позные рефлексы, наличие которых установил голландский физиолог Р. Магнус, (1924) в опытах на кошке. Имеется две разновидности этих рефлек-* сов - возникающие при наклоне и при поворотах головы.
При наклоне головы вниз (вперед) увеличиваются тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние разгибаются. При наклоне головы вверх (назад) возникают противоположные реакции - передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, а задние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей. Эти рефлексы возникают с проприорецепто-ров мышц шеи и фасций, покрывающих шейный отдел позвоночника. В условиях натурального поведения они увеличивают шансы животного достать пищу, находящуюся выше или ниже головы.
Вторая группа шейных позных рефлексов возникает с тех же рецепторов, но только при поворотах или наклоне головы вправо или влево. При этом повышается тонус мышц-разгибателей обеих конечностей на стороне, куда повернута (наклонена) голова, и повышается тонус мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие смещения центра тяжести в сторону поворота (наклона) головы - именно на этой стороне повышается тонус мышц-разгибателей обеих конечностей.
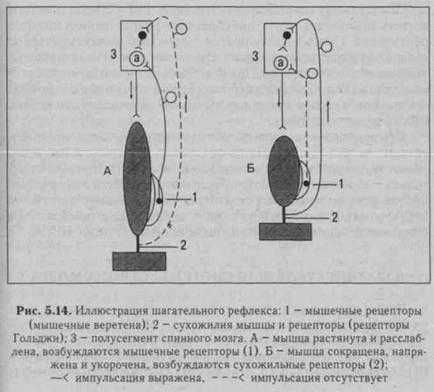
Д. Ритмические рефлексы - многократное повторное сгибание и разгибание конечностей. Примером этих рефлексов может быть шагательный рефлекс, который наблюдают у спинальной собаки, подвешенной с помощью лямок в станке.
Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим а-мотонейронам спинного мозга и возбуждают их (рис. 5.14 - А). Далее а-мотонейроны посылают импульсы к этой же скелетной мышце, что ведет к ее сокращению. Как только мышца сократилась (рис. 5.14 - Б), возбуждение мышечных веретен прекращается или сильно ослабевает (они уже не растянуты), начинают возбуждаться сухожильные рецепторы. Импульсы от последних поступают тоже в первую очередь в свой центр в спинном мозге, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение ос-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление (удлинение) ведет снова к возбуждению мышечных веретен и а-мотонейронов -мышца вновь сокращается. Вследствие ее сокращения возбуждают-
ся сухожильные рецепторы и тормозные клетки в спинном мозге, что вновь ведет к расслаблению скелетной мышцы, и т.д. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю. При этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу.
Для обеспечения поочередного сгибания и разгибания конечностей при шагательном рефлексе мышцы-сгибатели и разгибатели должны сокращаться и расслабляться последовательно друг за другом, что достигается с помощью торможения центра-антагониста при возбуждении центра агониста. Причем, если на одной ноге сокращены сгибатели, на другой ноге сокращаются разгибатели, что обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров сгибателей и разгибателей. На одноименной же стороне при возбуждении центра мышцы-сгибателя тормозится центр мышцы-разгибателя.
Шагательные координированные движения у спинального животного возможны в отсутствие обратной афферентации от проприо;, рецепторов. Они осуществляются с помощью межсегментарных связей на уровне спинного мозга. О наличии межсегментарных связей свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конечностей спинальной собаки при достаточно длительном и сильном раздражении одной конечности при интакт-ных афферентных путях.
При повреждении спинного мозга развивается гипертонус мышц, которые получают иннервацию от нижних сегментов, в частности гипертонус мышц нижних конечностей (рис. 5.15). Причина гипертонуса - возбуждение а-мотонейронов под влиянием афферентной импульсации от мышечных рецепторов (они обладают спонтанной активностью, активируются также с помощью а-мотонейронов) и выключение тормозных влияний вышележащих отделов ЦНС.
Физиология спинного мозга
I. Структурно-функциональная характеристика. Спинной мозг представляет собой тяж длиной 45 см у мужчин и около 42 см – у женщин. Он имеет сегментарное строение (31-33 сегмента). Каждый его сегмент связан с определенной частью тела. Спинной мозг включает пять отделов: шейный (С1-С8), грудной (Th1-Th12), поясничный (L1-L5), крестцовый (S1-S5) и копчиковый (Co1-Co3). В процессе эволюции в спинном мозге сформировалось два утолщения: шейное (сегменты, иннервирующие верхние конечности) и пояснично-крестцовое (сегменты, иннервирующие нижние конечности) как результат повышенной нагрузки на эти отделы. В указанных утолщениях соматические нейроны наиболее крупные, их больше, в каждом корешке этих сегментов больше нервных волокон, они имеют наибольшую толщину. Общее количество нейронов спинного мозга – около 13 млн. Из них 3% - мотонейроны, 97% - вставочные нейроны, из которых часть нейронов, которые относятся к вегетативной нервной системе.
► Классификация нейронов спинного мозга. Нейроны спинного мозга классифицируются по следующим признакам: 1) по отделу нервной системы (нейроны соматической и вегетативной нервной системы); 2) по назначению (эфферентные, афферентные, вставочные, ассоциативные); 3) по влиянию (возбуждающие и тормозные).
1. Эфферентные нейроны спинного мозга, относящиеся к соматической нервной системе, являются эффекторными, поскольку они иннервируют непосредственно рабочие органы – эффекторы (скелетные мышцы), их называют мотонейронами. Различают ά- и γ- мотонейроны.
ά-Мотонейроны иннервируют экстрафузальные мышечные волокна (скелетная мускулатура), их аксоны характеризуются высокой скоростью проведения возбуждения – 70-120 м/с. ά-Мотонейроны подразделяют на две подгруппы: ά1 – быстрые, иннервирующие быстрые белые мышечные волокна, их лабильность достигает 50 имп/с, и ά2 – медленные, иннервирующие медленные красные мышечные волокна, их лабильность – 10-15 имп/с. Низкая лабильность ά-мотонейронов объясняется длительной следовой гиперполяризацией, сопровождающей ПД. На одном ά- мотонейроне насчитывается до 20 тысяч синапсов: от кожных рецепторов, проприорецепторов и нисходящих путей вышележащих отделов ЦНС.
γ-Мотонейроны рассеяны среди ά-мотонейронов, их активность регулируется нейронами вышележащих отделов ЦНС, они иннервируют интрафузальные мышечные волокна мышечного веретена (мышечного рецептора). При изменении сократительной деятельности интрафузальных волокон под влиянием γ-мотонейронов изменяется активность мышечных рецепторов. Импульсация от мышечных рецепторов активирует ά-мотонейроны мышцы-антагониста, тем самым регулируется тонус скелетных мышц и двигательные реакции. Эти нейроны обладают высокой лабильностью – до 200 имп/с, но их аксонам свойственна низкая скорость проведения возбуждения – 10-40 м/с.
2. Афферентные нейронысоматической нервной системы локализуются в спинальных ганглиях и ганглиях черепных нервов. Их отростки, проводящие афферентную импульсацию от мышечных, сухожильных и кожных рецепторов, вступают в соответствующие сегменты спинного мозга и образуют синаптические контакты либо непосредственно на ά-мотонейронах (возбуждающие синапсы), либо на вставочных нейронах.
3. Вставочные нейроны (промежуточные, интернейроны) устанавливают связь с мотонейронами спинного мозга, с чувствительными нейронами, а также обеспечивают связь спинного мозга с ядрами ствола мозга, а через них – с корой большого мозга. Интернейроны могут быть как возбуждающими, так и тормозными, имеющими высокую лабильность – до 1000 имп/с.
4. Нейроны вегетативной нервной системы. Нейроны симпатической нервной системы являются вставочными, расположены в боковых рогах грудного, поясничного и частично шейного отделов спинного мозга (C8-L2). Эти нейроны фоновоактивны, частота разрядов 3-5 имп/с. Нейроны парасимпатического отдела нервной системы также вставочные, локализуются в сакральном отделе спинного мозга (S2-S4) и также фоновоактивны.
5. Ассоциативные нейроны образуют собственный аппарат спинного мозга, который устанавливает связь между сегментами и внутри сегментов. Ассоциативный аппарат спинного мозга участвует в координации позы, тонуса мышц, движений.
► Ретикулярная формация спинного мозга состоит из тонких перекладин серого вещества, пересекающихся в различных направлениях. Нейроны РФ имеют большое количество отростков. Ретикулярная формация обнаруживается на уровне шейных сегментов между передними и задними рогами, а на уровне верхнегрудных сегментов – между боковыми и задними рогами в белом веществе, примыкающем к серому.
► Нервные центры спинного мозга. В спинном мозге находятся центры регуляции большинства внутренних органов и скелетной мускулатуры.
1. Центры симпатического отдела вегетативной нервной системы локализованы в следующих сегментах: центр зрачкового рефлекса – С8 – Th2, регуляция деятельности сердца – Th1 – Th5, слюноотделения – Th2 – Th4, регуляция функции почек – Th5 – L3. Кроме этого здесь имеются сегментарно расположенные центры, регулирующие функции потовых желез и сосудов, гладких мышц внутренних органов, центры пиломоторных рефлексов.
2. Парасимпатическую иннервацию получают из спинного мозга (S2 – S4) все органы малого таза: мочевой пузырь, часть толстой кишки ниже ее левого изгиба, половые органы. У мужчин парасимпатическая иннервация обеспечивает рефлекторный компонент эрекции, у женщин – сосудистые реакции клитора и влагалища.
3. Центры управления скелетной мускулатуройнаходятся во всех отделах спинного мозга и иннервируют по сегментарному принципу скелетную мускулатуру шеи (C1 – C4), диафрагмы (C3 – C5), верхних конечностей (C5 – Th2), туловища (Th3 – L1) и нижних конечностей (L2 – S5).
Повреждения определенных сегментов спинного мозга или его проводящих путей вызывают специфические двигательные нарушения и расстройства чувствительности.
Каждый сегмент спинного мозга участвует в чувствительной иннервации трех дерматомов. Имеется дублирование и двигательной иннервации скелетных мышц, что повышает надежность их деятельности.
На рисунке показана иннервация метамеров (дерматомов) тела сегментами мозга: С – метамеры, иннервируемые шейными, Th – грудными, L – поясничными. S – крестцовыми сегментами спинного мозга, F – черепно-мозговыми нервами.
II. Функции спинного мозга– проводниковая и рефлекторная.
►Проводниковая функция. Проводниковая функция спинного мозга осуществляется с помощью нисходящих и восходящих проводящих путей.
Афферентная информация поступает в спинной мозг через задние корешки, эфферентная импульсация и регуляция функций различных органов и тканей организма осуществляется через передние корешки (закон Белла – Мажанди).
Каждый корешок представляет собой множество нервных волокон.
Все афферентные входы в спинной мозг несут информацию от трех групп рецепторов: 1) от кожных рецепторов (болевых, температурных, прикосновения, давления, вибрации); 2) от проприорецепторов (мышечных - мышечных веретен, сухожильных – рецепторов Гольджи, надкостницы и оболочек суставов); 3) от рецепторов внутренних органов – висцерорецепторов (механо- и хеморецепторов).
Медиатором первичных афферентных нейронов, локализующихся в спинальных ганглиях, является, по-видимому, субстанция Р.
Значение афферентной импульсации, поступающей в спинной мозг, заключается в следующем: 1) участие в координационной деятельности ЦНС по управлению скелетной мускулатурой. При выключении афферентной импульсации от рабочего органа управление им становится несовершенным. 2) участие в процессах регуляции функций внутренних органов. 3) поддержание тонуса ЦНС; при выключении афферентной импульсации наступает уменьшение суммарной тонической активности ЦНС. 4) несет информацию об изменениях окружающей среды. Основные проводящие пути спинного мозга приведены в таблице 1.
Таблица 1. Основные проводящие пути спинного мозга
| Восходящие (чувствительные) пути | Физиологическое значение |
| Клиновидный пучок (Бурдаха) проходит в задних столбах, импуьсация поступает в кору | Осознаваемая проприорецептивная импульсация от нижней части туловища и ног |
| Тонкий пучок (Голля), проходит в задних столбах, импульсация поступает в кору | Осознаваемая проприорецептивная импульсация от верхней части туловища и рук |
| Задний спинно-мозжечковый (Флексига) | Не осознаваемая проприорецептивная импульсация |
| Передний спинно-мозжечковый (Говерса) | То же |
| Латеральный спиноталамический | Болевая и температурная чувствительность |
| Передний спиноталамический | Тактильная чувствительность, прикосновение, давление |
| Нисходящие (двигательные) пути | Физиологическое значение |
| Латеральный кортикоспинальный (пирамидный) | Импульсы к скелетным мышцам |
| Передний кортикоспинальный (пирамидный) | То же |
| Руброспинальный (Монакова) проходит в боковых столбах | Импульсы, поддерживающие тонус скелетных мышц |
| Ретикулоспинальный, проходит в передних столбах | Импульсы, поддерживающие тонус скелетных мышц с помощью возбуждающих и тормозящих влияний на ά- и γ-мотонейроны, а также регулирующие состояние спинальных вегетативных центров |
| Вестибулоспинальный, проходит в передних столбах | Импульсы, обеспечивающие поддержание позы и равновесия тела |
| Тектоспинальный, проходит в передних столбах | Импульсы, обеспечивающие осуществление зрительных и слуховых двигательных рефлексов (рефлексов четверохолмия) |
III. Рефлексы спинного мозга. Спинной мозг выполняет рефлекторную соматическую и рефлекторную вегетативную функции.
Сила и длительность всех спинальных рефлексов увеличиваются при повторном раздражении, при увеличении площади раздражаемой рефлексогенной зоны вследствие суммации возбуждения, а также при увеличении силы стимула.
Соматические рефлексы спинного мозга по своей форме в основном являются сгибательными и разгибательными рефлексами сегментарного характера. Соматические спинальные рефлексы можно объединить в две группы по следующим признакам:
╠ Во-первых, по рецепторам, раздражение которых вызывает рефлекс: а) проприоцептивные, б) висцероцептивные, в) кожные рефлексы. Рефлексы, возникающие с проприорецептров, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висцерорецептивные (висцеромоторные) рефлексы возникают с рецепторов внутренних органов и проявляются в сокращении мышц брюшной стенки, грудной клетки и разгибателей спины. Возникновение висцеромоторных рефлексов связано с конвергенцией висцеральных и соматических нервных волокон к одним и тем же интернейронам спинного мозга.
╠ Во-вторых, по органам: а) рефлексы конечностей; б) брюшные рефлексы; в) яичковый рефлекс; г) анальный рефлекс.
1. Рефлексы конечностей. Эту группу рефлексов в клинической практике исследуют наиболее часто.
▓ Сгибательные рефлексы.Сгибательные рефлексы делятся на фазные и тонические.
▒ Фазные рефлексы – это однократное сгибание конечности при однократном раздражении кожи или проприорецепторов. Одновременно с возбуждением мотонейронов мышц-сгибателей происходит реципрокное торможение мотонейронов мышц-разгибателей. Рефлексы, возникающие с рецепторов кожи, являются полисинаптическими, они имеют защитное значение. Рефлексы, возникающие с проприорецепторов, могут быть моносинаптическими и полисинаптическими. Фазные рефлексы с проприорецепторов участвуют в формировании акта ходьбы. По степени выраженности фазных сгибательных и разгибательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения.
В клинике исследуют следующие сгибательные фазные рефлексы: локтевой и ахиллов (проприоцептивные рефлексы) и подошвенный рефлекс (кожный). Локтевой рефлекс выражается в сгибании руки в локтевом суставе, возникает при ударе рефлекторным молоточком по сухожилию m. вiceps brachii (при вызове рефлекса рука должна быть слегка согнута в локтевом суставе), его дуга замыкается в 5-6-ом шейных сегментах спинного мозга (С5 – С6). Ахиллов рефлекс выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию, рефлекторная дуга замыкается на уровне крестцовых сегментов (S1 – S2). Подошвенный рефлекс – сгибание стопы и пальцев при штриховом раздражении подошвы, дуга рефлекса замыкается на уровне S1 – S2.
▒ Тонические сгибательные, а также разгибательные рефлексы возникают при длительном растяжении мышц, их главное назначение – поддержание позы. Тоническое сокращение скелетных мышц является фоновым для осуществления всех двигательных актов, осуществляемых с помощью фазических сокращений мышц.
▓ Разгибательные рефлексы, как сгибательные, бывают фазными и тоническими, возникают с проприорецепторов мышц-разгибателей, являются моносинаптическими. Одновременно со сгибательным рефлексом возникает перекрестный разгибательный рефлекс другой конечности.
▒ Фазные рефлексы возникают в ответ на однократное раздражение мышечных рецепторов. Например, при ударе по сухожилию четырехглавой мышцы бедра ниже коленной чашечки возникает коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы бедра. Во время разгибательного рефлекса мотонейроны мышц-сгибателей тормозятся с помощью вставочных тормозных клеток Реншоу (реципрокное торможение). Рефлекторная дуга коленного рефлекса замыкается во втором – четвертом поясничных сегментах (L2 – L4). Фазные разгибательные рефлексы участвуют в формировании ходьбы.
▒ Тонические разгибательные рефлексы представляют собой длительное сокращение мышц-разгибателей при длительном растяжении сухожилий. Их роль – поддержание позы. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикального положения. Тоническое сокращение мышц спины обеспечивает осанку человека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют также миотатическими.
▓ Рефлексы позы – перераспределение мышечного тонуса, возникающее при изменении положения тела или отдельных его частей. Рефлексы позы осуществляются с участием различных отделов ЦНС. На уровне спинного мозга замыкаются шейные позные рефлексы. Имеется две группы этих рефлексов – возникающие при наклоне и при повороте головы.
▒ Первая группа шейных позных рефлексов существует только у животных и возникает при наклоне головы вниз (кпереди). При этом увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние разгибаются. При наклоне головы вверх (кзади) возникают противоположные реакции – передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, а задние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей. Эти рефлексы возникают с проприорецепторов мышц шеи и фасций, покрывающих шейный отдел позвоночника. В условиях естественного поведения они увеличивают животному шанс достать пищу, находящуюся выше или ниже уровня головы.
Рефлексы позы верхних конечностей у человека утрачены. Рефлексы нижних конечностей выражаются не в сгибании или разгибании, а в перераспределении мышечного тонуса, обеспечивающего сохранение естественной позы.
▒ Вторая группа шейных позных рефлексов возникает с тех же рецепторов, но только при поворотах головы вправо или влево. При этом повышается тонус мышц-разгибателей обеих конечностей на стороне, куда повернута голова, и повышается тонус мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие изменения положения центра тяжести после поворота головы. Центр тяжести смещается в сторону поворота головы – именно на этой стороне повышается тонус мышц-разгибателей обеих конечностей. Подобные рефлексы наблюдаются и у человека.
▓ Ритмические рефлексы – многократное повторное сгибание и разгибание конечностей. Примерами могут служить чесательный и шагательный рефлексы.
2. Брюшные рефлексы (верхний, средний и нижний) проявляются при штриховом раздражении кожи живота. Выражаются в сокращении соответствующих участков мускулатуры стенки живота. Это защитные рефлексы. Для вызова верхнего брюшного рефлекса раздражение наносят параллельно нижним ребрам непосредственно под ними, дуга рефлекса замыкается на уровне грудных сегментов спинного мозга (Th8 – Th9). Средний брюшной рефлекс вызывают раздражением на уровне пупка (горизонтально), дуга рефлекса замыкается на уровне Th9 – Th10. Для получения нижнего брюшного рефлекса раздражение наносят параллельно паховой складке (рядом с ней), дуга рефлекса замыкается на уровне Th11 – Th12.
3. Кремастерный (яичковый) рефлекс заключается в сокращении m. сremaster и поднимании мошонки в ответ на штриховое раздражение верхней внутренней поверхности кожи бедра (кожный рефлекс), это также защитный рефлекс. Его дуга замыкается на уровне L1 – L2.
4. Анальный рефлекс выражается в сокращении наружного сфинктера прямой кишки в ответ на штриховое раздражение или укол кожи вблизи заднего прохода, дуга рефлекса замыкается на уровне S2 – S5.
Вегетативные рефлексы спинного мозга осуществляются в ответ на раздражение внутренних органов и заканчиваются сокращением гладкой мускулатуры этих органов. Вегетативные рефлексы имеют в спинном мозге свои центры, которые обеспечивают иннервацию сердца, почек, мочевого пузыря и т.д.
IV. Спинальный шок. Перерезка или травма спинного мозга вызывает явление, получившее название спинального шока. Спинальный шок выражается в резком падении возбудимости и угнетении деятельности всех рефлекторных центров спинного мозга, расположенных ниже места перерезки. Во время спинального шока раздражители, которые обычно вызывали рефлексы, оказываются недейственными. В то же время деятельность центров, расположенных выше перерезки, сохраняется. После перерезки исчезают не только скелетно-моторные рефлексы, но и вегетативные. Снижается кровяное давление, отсутствуют сосудистые рефлексы, акты дефекации и мочеиспускания.
Продолжительность шока различна у животных, стоящих на различных ступенях эволюционной лестницы. У лягушки шок продолжается 3-5 минут, у собаки – 7-10 дней, у обезьяны – больше 1 месяца, у человека – 4-5 месяцев. Когда шок проходит, рефлексы восстанавливаются. Причиной спинального шока является выключение вышерасположенных отделов головного мозга, оказывающих на спинной мозг активирующее влияние, в котором большая роль принадлежит ретикулярной формации ствола мозга.
Позные и локомоторные рефлексы спинного мозга. Шагательные движения
а) Рефлексы положения и локомоции. Позные и локомоторные рефлексы спинного мозга. Положительная реакция опоры. Надавливание на подушечку лапы децеребрированного животного ведет к разгибанию конечности, которое противодействует давлению, приложенному к стопе. Данный рефлекс настолько сильный, что если животное с пересеченным несколько месяцев назад спинным мозгом (т.е. на фоне усиленных спинальных рефлексов) поставить на стопы, рефлекторное разгибание конечностей часто сопровождается напряжением мышц, достаточным для удержания веса тела. Этот рефлекс называют положительной реакцией опоры.
В основе положительной реакции опоры лежит сложный контур во вставочных нейронах, подобный контурам, ответственным за сгибательный и перекрестный разгибательный рефлексы. Место давления на подушечку стопы определяет направление, в котором конечность будет разгибаться; давление с одной стороны вызовет разгибание в этом направлении — эффект, называемый реакцией магнита. Это помогает животному удержаться от падения в эту сторону.
б) Спинальные выпрямительные рефлексы. Если спинальное животное положить на бок, оно будет совершать некоординированные движения, пытаясь подняться. Это называют спиналъным выпрямительным рефлексом. Наличие такого рефлекса свидетельствует об интеграции некоторых относительно сложных позных рефлексов на уровне спинного мозга. Действительно, животное, выхоженное после пересечения грудного отдела спинного мозга между уровнями иннервации передних и задних конечностей и практически здоровое в других отношениях, может подниматься из положения лежа и даже ходить, используя задние конечности в дополнение к передним. У опоссума с подобной перерезкой грудного отдела спинного мозга шагательные движения задних конечностей почти не отличаются от этих движений у нормальных животных, за исключением того, что они не синхронизированы с движениями передних конечностей.
Сгибательный рефлекс, перекрестный разгибательный рефлекс и реципрокное торможение Диагональные шагательные движения, выполняемые спинальным животным
Шагательные движения и ходьба
а) Ритмические шагательные движения одной конечности. У спинальных животных часто наблюдаются ритмические шагательные движения. Если поясничный отдел спинного мозга отделить от остальной его части и рассечь вдоль центральной линии с целью блокады нервной связи между двумя сторонами спинного мозга и, следовательно, между обеими конечностями, каждая из задних конечностей все еще может выполнять шагательные движения. Так, направленное вперед сгибание конечности через секунду или несколько позже сменяется направленным назад разгибанием. Затем опять осуществляется сгибание, и цикл повторяется снова и снова.
Такое колебательное движение вперед-назад в связи с последовательными сокращениями сгибателей и разгибателей может происходить даже после перерезки чувствительных нервов и, по-видимому, является результатом взаимодействия в пределах самого спинного мозга контуров реципрокного торможения, циркулирующего между нейронами, регулирующими мышцы-агонисты и мышцы-антагонисты.
Сенсорные сигналы от подушечек лап и рецепторов положения вокруг суставов играют важную роль в регуляции давления на стопы и частоты шагательных движений, когда стопам предоставляется возможность касаться поверхности. На самом деле спинальный механизм регуляции шагания, вероятно, еще сложнее. Например, если кончик стопы неожиданно наталкивается на препятствие при выпаде вперед, движение временно останавливается; затем последовательно осуществляется подъем стопы и перемещение ее вперед так, чтобы оказаться над препятствием. Это называют рефлексом спотыкания. Таким образом, спинной мозг является «умным» регулятором ходьбы.
б) Реципрокное шагание противоположных конечностей. Если поясничный отдел спинного мозга не расщеплен посередине, каждый раз, когда происходит шагательное движение одной конечности вперед, противоположная конечность обычно движется назад. Этот эффект является результатом реципрокной иннервации двух конечностей.
Шагание на месте. Чесательный рефлекс и рефлексы мышечного спазма
а) Диагональное шагание всех четырех конечностей - рефлекс шагания на месте. Если спинальное животное (выхоженное после высокой перерезки спинного мозга на уровне шеи над областью иннервации верхних конечностей) удерживать над полом так, чтобы его лапы свободно свисали вниз (для облегчения понимания просим вас изучить рисунок ниже), растяжение конечностей иногда возбуждает шагательные рефлексы, вовлекающие все четыре конечности.

Диагональные шагательные движения, выполняемые спинальным животным
В целом, шагательные движения передних и задних конечностей осуществляются по диагонали. Эта диагональная реакция является еще одним проявлением реципрокной иннервации, на этот раз осуществляемой вдоль всей длины спинного мозга в направлении вверх и вниз между передними и задними конечностями. Такую организацию шагания называют рефлексом шагания на месте.
б) Рефлекс галопирования. Другим типом рефлекса у спинальных животных является рефлекс галопирования, при котором обе передние конечности движутся вместе назад, тогда как обе задние конечности движутся вперед. Это происходит, когда почти равные стимулы растяжения или давления действуют одновременно на конечности с обеих сторон тела; неравная стимуляция вызывает диагональный шагательный рефлекс. Такое представление поддерживается особенностями шагания и галопа в нормальных условиях. При шагании одновременно стимулируются только одна передняя и одна задняя конечности, что способствует продолжению шагания.
Во время галопа все конечности стимулируются примерно в равной степени, и животное сразу ударяет о землю обеими передними и обеими задними конечностями.
Сгибательный рефлекс, перекрестный разгибательный рефлекс и реципрокное торможение
Чесательный рефлекс
Особенно важным спинальным рефлексом у некоторых животных является чесательный рефлекс, который запускается ощущениями зуда или щекотки. Этот рефлекс включает две функции: (1) ощущение положения, которое позволяет лапе точно найти место раздражения на поверхности тела; (2) чесателъные движения вперед-назад.
Ощущение положения чесательного рефлекса является высокоразвитой функцией. Так, если блоха заползает на плечо спинального животного, его задняя лапа может определить ее положение, несмотря на то, что для приведения лапы к месту, где ползает блоха, должны одновременно и весьма координированно сокращаться 19 мышц конечности. На еще большую сложность рефлекса указывает факт, что при переходе блохи на другую сторону первая лапа прекращает чесание, а противоположная лапа начинает движения вперед-назад и в итоге находит блоху.
Движения вперед-назад, как и шагательные локомоторные движения, вовлекают контуры с реципрокной иннервацией, которые вызывают осцилляцию.
Спинальные рефлексы, вызывающие мышечный спазм
У людей часто наблюдается локальный мышечный спазм. Во многих, если не в большинстве случаев, причиной такого спазма является локальная боль.
а) Мышечный спазм в результате перелома кости. Один тип клинически важного спазма происходит в мышцах, окружающих сломанную кость. Этот спазм является результатом болевых импульсов, возникающих в сломанных концах кости, что заставляет мышцы, окружающие эту область, сокращаться тонически. Уменьшение боли при введении локального анестетика в область перелома снижает спазм; глубокий общий наркоз, например эфирный, также уменьшает спазм. Часто предварительное использование одного из этих видов обезболивания необходимо для успешного преодоления спазма в процессе приведения концов переломанной кости в правильное положение.
б) Спазм брюшных мышц при перитоните. Другим типом локального спазма, вызываемого спинальными рефлексами, является спазм мышц передней брюшной стенки в результате раздражения пристеночной брюшины при перитоните. В этом случае облегчение боли позволяет спастически сокращенным мышцам расслабиться. Тот же тип спазма часто происходит во время хирургической операции. Например, при операции на брюшной полости болевые импульсы от пристеночной брюшины часто заставляют брюшные мышцы сильно сокращаться, иногда выдавливая кишечник через хирургический разрез. По этой причине для проведения операций внутри брюшной полости обычно требуется глубокая анестезия.
в) Судороги мышц. Другим типом локального спазма является типичная мышечная судорога. Электромиографические исследования указывают, что причина некоторых мышечных судорог следующая. Любой местный раздражающий фактор или нарушение метаболизма в мышце, например сильный холод, прекращение кровоснабжения или перенапряжение, может вызвать проведение болевых или других сенсорных сигналов от мышцы к спинному мозгу, что, в свою очередь, вызывает рефлекторное сокращение мышцы по механизму обратной связи. Полагают, что сокращение еще больше стимулирует сенсорные рецепторы, заставляя спинной мозг усилить интенсивность сокращения.
Таким образом, благодаря развитию положительной обратной связи даже небольшое первичное раздражение вызывает все более сильное сокращение, вплоть до развития выраженного судорожного сокращения мышцы.
Читайте также: