Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
Добавил пользователь Евгений Кузнецов Обновлено: 29.01.2026
Адсорбция вирусов.Первая стадия репродукции вирусов — адсорбция, т. е. прикрепление вириона к поверхности клетки. Она протекает в две фазы. Первая фаза — неспецифическая, обусловленная ионным притяжением между вирусом и клеткой, включая и другие механизмы. Вторая фаза адсорбции — высокоспецифическая, обусловленная гомологией, комплемен-тарностью рецепторов чувствительных клеток и «узнающих» их белковых лигандов вирусов. Белки на поверхности вирусов, узнающие специфические клеточные рецепторы и взаимодействующие с ними, называются прикрепительными белками (в основном это гликопротеины) в составе липопротеиновой оболочки.
Специфические рецепторы клеток имеют различную природу, являясь белками, липидами, углеводными компонентами белков, липидов и др. Так, рецепторами для вируса гриппа является сиаловая кислота в составе гли-копротеинов и гликолипидов (ганглиозидов) клеток дыхательных путей. Вирусы бешенства адсорбируются на ацетилхолиновых рецепторах нервной ткани, а вирусы иммунодефицита человека — на СО4-рецепторах Т-хелперов, моноцитов и дендритных клеток. На одной клетке находится от десяти до ста тысяч специфических рецепторов, поэтому на ней могут адсорбироваться десятки и сотни вирионов.
Наличие специфических рецепторов лежит в основе избирательности поражения вирусами определенных клеток, тканей и органов. Это так называемый тропизм(греч. tropos — поворот, направление). Например, вирусы, репродуцирующиеся преимущественно в клетках печени, называются гепатотропными, в нервных клетках — нейротропными, в иммунокомпетентных клетках — иммунотропными и т. д.
Проникновение вирусов в клетку.Вирусы проникают в клетку путем рецептор-зависимого эндоцитоза (виропексиса), или слияния оболочки вируса с клеточной мембраной, или же в результате сочетания этих механизмов.
1. Рецептор-зависимый эндоцитоз происходит в результате захватывания и поглощения вириона клеткой: клеточная мембрана с прикрепленным вирионом впячивается с образованием внутриклеточной вакуоли (эндосомы), содержащей вирус. За счет АТФ-зависимого «протонного» насоса содержимое эндосомы закисляется, что приводит к слиянию липопротеиновой оболочки сложно организованного вируса с мембраной эндосомы и выходу вирусного нуклеокапсида в цитозоль клетки. Эндосомы объединяются с лизосомами, которые разрушают оставшиеся вирусные компоненты. Процесс выхода безоболочечных (просто организованных) вирусов из эндосомы в цитозоль остается малоизученным.
2. Слияние обточки вириона с клеточной мембраной характерно только для некоторых оболочечных вирусов (парамиксовирусов, ретровиру-сов, герпесвирусов), в составе которых имеются белки слияния. Происходит точечное взаимодействие вирусного белка слияния с липидами клеточной мембраны, в результате чего вирусная липопротеиновая оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса попадает в цитозоль.
А) «Раздевание» (депротеинизация) вирусов.В результате высвобождается его внутренний компонент, способный вызывать инфекционный процесс. Первые этапы «раздевания» вируса начинаются в процессе его проникновения в клетку путем слияния вирусных и клеточных мембран или же при выходе вируса из эндосомы в цитозоль. Последующие этапы «раздевания» вируса тесно взаимосвязаны с их внутриклеточным транспортом к местам депротеинизации. Для разных вирусов существуют свои специализированные участки «раздевания» в клетке: для пикорнавирусов— в цитоплазме с участием лизосом, аппарата Гольджи; для герпесвирусов — околоядерное пространство или поры ядерной мембраны; для аденовирусов — сначала структуры цитоплазмы, а затем ядро клетки. Конечными продуктами «раздевания» могут быть нуклеиновая кислота, нуклеопротеин (нуклеокапсид) или сердцевина вириона. Так, конечным продуктом раздевания пикарновирусов является нуклеиновая кислота, ковалентно связанная с одним из внутренних белков. А у многих оболочечных РНК-содержащих вирусов конечными продуктами «раздевания» могут быть нуклеокапсиды или сердцевины, которые не только не препятствуют экспрессии вирусного генома, а, более того, защищают его от клеточных протеаз и регулируют последующие биосинтетические процессы.
В) Синтез вирусных компонентов. Синтез белков и нуклеиновых кислот вируса, который разобщен во времени и пространстве. Синтез осуществляется в разных частях клетки, поэтому такой способ размножения вирусов называется дизъюнктивным (от лат. disjunctus — разобщенный).
С)Синтез вирусных белков. В зараженной клетке вирусный геном кодирует синтез двух групп белков:
1. неструктурных белков, обслуживающих внутриклеточную репродукцию вируса на разных его этапах;
2. структурных белков, которые входят в состав вириона (геномные, связанные с геномом вируса, капсидные и су-перкапсидные белки).
К неструктурным белкамотносятся: 1) ферменты синтеза РНК или ДНК (РНК- или ДНК-полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома; 2) белки-регуляторы; 3) предшественники вирусных белков, отличающиеся своей нестабильностью в результате быстрого нарезания на структурные белки; 4) ферменты, модифицирующие вирусные белки, например, протеиназы и протеинкиназы.
Синтез белков в клетке осуществляется в соответствии с хорошо известными процессами транскрипции(от лат. transcriptio — переписывание) путем «переписывания» генетической информации с нуклеиновой кислоты в нуклео-тидную последовательность информационной РНК (иРНК) и трансляции (от лат. translatio — передача) — считывания иРНК на рибосомах с образованием белков. Передача наследственной информации в отношении синтеза иРНК у разных групп вирусов неодинакова.
I. ДНК-содержашие вирусы реализуют генетическую информацию так же, как и клеточный геном, по схеме:
геномная ДНК вируса -» транскрипция иРНК -» трансляция белка вируса.
Причем ДНК-содержашие вирусы используют для этого процесса клеточную полимеразу (вирусы, геномы которых транскрибируются в ядре клетки — аденовирусы, па-повавирусы, герпесвирусы) или собственную РНК-полимеразу (вирусы, геномы которых транскрибируются в цитоплазме, например поксвирусы).
II. Плюс-нитевые РНК-содержашие вирусы(например, пикорнавирусы, флавивирусы, тогавирусы) имеют геном, выполняющий функцию иРНК; он распознается и транслируется рибосомами. Синтез белков у этих вирусов осуществляется без акта транскрипции по схеме:
геномная РНК вируса-> трансляция белка вируса.
III. Геном минус-однонитевых РНК-содержаших вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов) и двунитевых (реовирусов) служит матрицей, с которой транскрибируется иРНК, при участии РНК-полимеразы, связанной с нуклеиновой кислотой вируса. Синтез белка у них происходит по схеме:
геномная РНК вируса-» транскрипция и-РНК- трансляция белка вируса.
IV. Ретровирусы(вирусы иммунодефицита человека, онкогенные ретровирусы) имеют уникальный путь передачи генетической информации. Геном ретровирусов состоит из двух идентичных молекул РНК, т. е. является диплоидным. В составе ретровирусов есть особый вирусоспецифический фермент — обратная транскриптаза, или ревертаза, с помощью которой осуществляется процесс обратной транскрипции, т. е. на матрице геномной РНК синтезируется комплементарная однонитевая ДНК (кДНК). Комплементарная нить ДНК копируется с образованием двунитевой комплементарной ДНК, которая интегрирует в клеточный геном и в его составе транскрибируется в иРНК с помощью клеточной ДНК-зависимой РНК-полимеразы. Синтез белков для этих вирусов осуществляется по схеме:
геномная РНК вируса-> комплементарная ДНК-» транскрипция иРНК
-»трансляция белка вируса.
Репликация вирусных геномов,т. е. синтез вирусных нуклеиновых кислот, приводит к накоплению в клетке копий исходных вирусных геномов, которые используются при сборке вирионов. Способ репликации генома зависит от типа нуклеиновой кислоты вируса, наличия вирусоспецифических или клеточных полимераз, а также от способности вирусов индуцировать образование полимераз в клетке.
Механизм репликации отличается у вирусов, имеющих:
1) двунитевую ДНК;
2) однонитевую ДНК;
3) плюс-однонитевую РНК;
4) минус-одноните-вую РНК;
5) двунитевую РНК;
6) идентичные плюс-нитевые РНК (ретровирусы).
1. Двунитевые ЛНК-вирусы. Репликация двунитевых вирусных ДНК происходит обычным полуконсервативным механизмом: после рас-
плетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. К этим вирусам относится большая
группа вирусов, которые содержат двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме, как папилломавирусы. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре.
Уникальный механизм репликации характерен для гепаднавирусов (вируса гепатита В). Геном гепаднавирусов представлен дву-нитевой кольцевой ДНК, одна нить которой короче (неполная плюс-нить) другой нити. Первоначально достраивается (рис. 3.7). Затем полная двунитевая ДНК с помощью клеточной ДНК-зависимой РНК-полимеразы транскрибируется с образованием небольших молекул иРНК и полной однонитевой плюс-РНК. Последняя называется прегеномной РНК; она является матрицей для репликации генома вируса. Синтезированные иРНК участвуют в процессе трансляции белков, в том числе вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). С помощью этого фермента мигрирующая в цитоплазму прегеномная РНК обратно транскрибируется в минус-нить ДНК, которая, в свою очередь, служит матрицей для синтеза плюс-нити ДНК. Этот процесс заканчивается образованием двунитевой ДНК, содержащей неполную плюс-нить ДНК.
2.Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы. Парвовирусы
используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы послед
него. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
3.Плюс-однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов — пикорнавирусы, флавивирусы, тогавирусы (рис.3.8), у которых геномная плюс-нить РНК выполняет функцию иРНК. Например, РНК
полиовирусов после проникновения в клетку связывается с рибосомами, работая как иРНК, и на ее основе синтезируется большой полипептид, который расщепляется на фрагменты:
РНК-зависимую РНК-полимеразу, вирусные протеазы и капсидные белки. Полимераза на основе геномной плюс-нити РНК синтезирует минус-нить РНК; формируется временно двойная РНК, названная промежуточным
репликативным звеном. Это промежуточное репликативное звено состоит из полной плюс-нити РНК и многочисленных частично завершенных минус-нитей. Когда образованы все минус-нити, они используются как шаблоны
для синтеза новых плюс-нитей РНК. Этот механизм используется как для размножения геномной РНК вируса, так и для синтеза большого количества вирусных белков.
4.Минус-однонитевые РНК-вирусы. Минус -однонитевые РНК-вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус-
нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства
5.Двунитевые РНК-вирусы. Механизм репликации этих вирусов (реовирусов и ротави-русов) сходен с репликацией минус-однонитевых РНК-вирусов. Отличие состоит в том, что образовавшиеся в процессе транскрипции плюс-нити функционируют не только как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей
РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
6. Ретровирусы (плюс-нитевые диплоидные РНК-содержащие вирусы). Обратная транс-криптаза ретровирусов синтезирует (на матрице РНК-вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо (рис. 3.10). Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. Многочисленные вирионные РНК образуются в результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы.
Формирование вирусов.Вирионы формируются путем самосборки: составные части вириона транспортируются в места сборки вируса — участки ядра или цитоплазмы клетки. Соединение компонентов вириона обусловлено наличием гидрофобных, ионных, водородных связей и стерического соответствия.
Существуют следующие общие принципы сборки вирусов:
Формирование вирусов— многоступенчатый процесс с образованием промежуточных форм, отличающихся от зрелых вирионов по
составу полипептидов.
□ Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов.
□ У сложноустроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса).
Причем сборка вирусов, реплицирующихся в ядре клетки, происходит с участием мембраны ядра, а сборка вирусов, репликация которых идет в цитоплазме, осуществляется с
участием мембран эндоплазматической сети или плазматической мембраны, куда встраиваются гликопротеины и другие белки оболочки вируса.
□ У ряда сложноустроенных вирусов минус-нитевых РНК-вирусов (ортомиксовирусов, парамиксовирусов) в сборку вовлекается так называемый матриксный белок (М-белок), который расположен под модифицированной клеточной ембраной. Обладая гидрофобными свойствами, он выполняет роль посредника между нуклеокапсидом и вирусной липопротеиновой оболочкой.
□ Сложноустроенные вирусы в процессе формирования включают в свой состав некоторые компоненты клетки хозяина, например липиды и углеводы.
Выход вирусов из клетки.Полный цикл репродукции вирусов завершается через 5—6 ч (вирус гриппа и др.) или через несколько суток (гепатовирусы, вирус кори и др.). Процесс репродукции вирусов заканчивается выходом их из клетки, который происходит взрывным путем или почкованием, экзоцитозом.
□ Взрывной путь: из погибающей клетки одновременно выходит большое количество вирионов. По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки.
□ Почкование, экзоцшпт присущи вирусам, имеющим липопротеиновую оболочку, которая является производной от клеточных мембран. Сначала образовавшийся нуклеокапсид или сердцевина вириона транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида или сердцевины вириона с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. При этом клетка способна длительно сохранять жизнеспособность и продуцировать вирусное потомство.
Почкование вирусов, формирующихся в цитоплазме, может происходить либо через плазматическую мембрану (например, парамиксовирусы, тогавирусы), либо через мембраны эндоплазматической сети с последующим их выходом на поверхность клетки (например, буньявирусы).
Вирусы, формирующиеся в ядре клетки (например, герпесвирусы), почкуются в перинуклеарное пространство через модифицированную ядерную мембрану, приобретая таким образом липопротеиновую оболочку. Затем они транспортируются в составе цитоплазма-тических везикул на поверхность клетки.
Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
Фундаментальное отличие вирусов от других инфекционных агентов состоит в механизмах их репродукции: вирусы не способны к самостоятельному размножению. Репликация генетического материала вирусов (ДНК или РНК), а также экспрессия вирусных генов (вирусный ген -» вирусный белок) осуществляются при помощи механизмов репликации, транскрипции, сплайсинга и трансляции инфицированной клетки-хозяина. Циклы репродукции вирусов в инфицированной клетке зависят от типа вирусной нуклеиновой кислоты.
Репродукция +РНК-вирусов
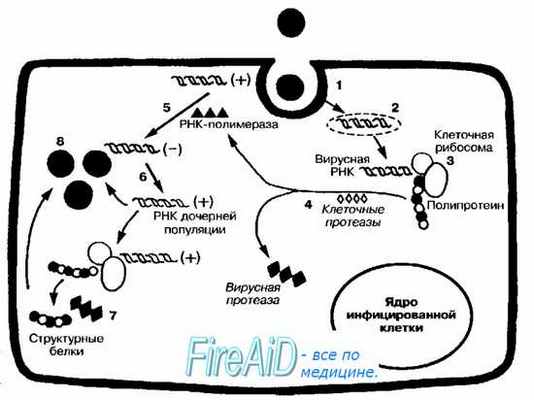
Рис. 5-1. Репродуктивный цикл +РНК-содержащих вирусов. После адсорбции вирус проникает в клетку путём виропексиса (1). Вирусная +РНК высвобождается в цитоплазме (2). Поскольку молекулярная симметрия вирусной +РНК аналогична мРНК, то +РНК может непосредственно распознаваться и транслироваться рибосомами (3). Образуется гигантская полипротеиновая молекула. Клеточные протеазы «нарезают» образующийся вирусный полипротеин (4) с образованием РНК-зависимой РНК-полимеразы, вирусной протеазы, ингибиторов синтеза клеточных РНК и различных структурных белков. РНК-полимераза катализирует образование -РНК на матрице родительской +РНК (5). Затем уже на матрице -РНК происходит многократный синтез молекул +РНК (6), участвующих в синтезе вирусных белков (7) либо входящих в состав генома дочерних популяций вирусов (8).
+РНК-вирусы (например, пикорна- и тогавирусы) проникают в чувствительную клетку путём виропексиса. Репродуктивный цикл начинается после высвобождения вирусного генома в цитоплазме (рис. 5-1).
Пикорнавирусы. Репродукция пикорнавирусов
Пикорнавирусы размножаются подобно большинству +РНК-содержащих вирусов. Среди них наиболее хорошо изучен полиовирус (возбудитель полиомиелита), имеющий небольшие размеры, голый капсид и однонитевую молекулу +РНК. Адсорбционные возможности полиовиру-са ограничены клетками приматов и человека, имеющих специфические рецепторы. Репликация +РНК полностью происходит в цитоплазме клетки. Репродуктивный цикл подразделяют на раннюю и позднюю стадии.
Ранняя стадия репродукции включает процессы, происходящие после проникновения вируса в клетку до образования большого количества копий +РНК в процессе репликации. Поздняя стадия репродукции включает синтез белков капсида. Образованные в репликативном синтезе молекулы +РНК функционируют как мРНК, транслируясь в один гигантский полипептид, нарезаемый протеазами на капсидные белки. После самосборки капсидов формируется зрелая дочерняя популяция. Поскольку пикорнавирусы лишены суперкапсида, то высвобождение популяции сопровождается массивным повреждением клеточной мембраны с последующим лизисом инфицированной клетки.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
68. Пикорнавирусы. Структура вирионов. Репродукция
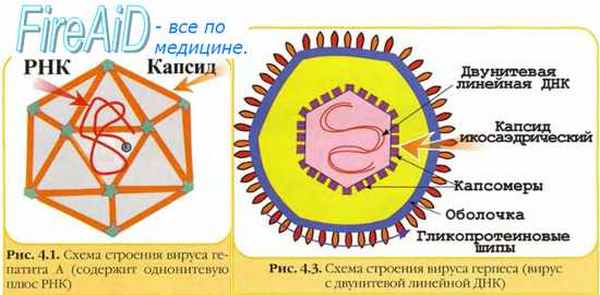
Пикорнавирусы — семейство безоболочечных вирусов, содержащих однонитевую плюс-РНК. Семейство насчитывает более 230 представителей и состоит из 8 родов, среди которых наибольшее значение имеют Enterovirus, Rhinovirus, Cardiovirus, Aphthovirus и Hepatovirus. Вирусы поражают позвоночных; многие представители семейства патогенны для человека.
Структура. Пикорнавирусы относятся к простым вирусам (без оболочки). Диаметр вируса — около 30 нм. Однонитевая плюс-РНК с протеином VPg окружена икосаэдрическим кап- сидом. Капсид состоит из 12 пятиугольников (пентамеров), каждый из которых, в свою очередь, состоит из 5 белковых субъединиц (протомеры). Протомеры образованы 4 вирусными полипептидами (VP1, VP2, VP3, VP4).
Репродукция. Вирус взаимодействует с рецепторами на поверхности клетки. Геном вируса может поступить в клетку путем эндоцитоза (1') с последующим выходом нуклеиновой кислоты (2) из вакуоли или путем инъекции РНК через цитоплазматическую мембрану (1) клетки. На конце РНК имеется вирусный протеин (3) — VPg. Геном используется как иРНК для синтеза белка (4,5). Один большой полипротеин (4) транслируется с вирусного генома. Затем полипротеин расщепляется на индивидуальные вирусные протеины, включая РНК-зависимую полимеразу. Полимераза синтезирует минус-нить-матрицу с поверхности плюс-нити и реплицирует геном. VPg ковалентно присоединяется к (5') концу вирусного генома. Структурные белки собираются в капсид (6), в него включается геном, образуя вирион. Вирионы освобождаются из клетки посредством ее лизиса. Репродукция происходит в цитоплазме клеток и сопровождается цитопатическим действием. В культурах клеток под агаровым покрытием вирусы образуют бляшки.
Энтеровирусы обитают преимущественно в кишечнике человека и вызывают разнообразные по клиническим проявлениям болезни человека. Источником инфекции являются больные и носители. Из организма больного вирусы выделяются с носоглоточной слизью и фекалиями, из организма вирусоносителя — с фекалиями. Энтеровирусы передаются через воду, почву, пищевые продукты, предметы обихода, загрязненные руки, через мух. В первые 1-2 недели болезни они выделяются из носоглотки, обуславливая воздушно-капельный путь передачи. Вызывают полиомиелит, полиомиелитоподобные заболевания, менингиты, энцефалиты, миалгию, миокардит, острые респираторные заболевания, гастроэнтерит, конъюнктивит, лихорадочные заболевания с сыпью и без нее и др.
Риновирус — РНК-содержащие вирусы рода Rhinovirus семейства Picornaviridae; наиболее частые возбудители острых инфекций верхних дыхательных путей (ОРВИ). Идентифицировано свыше 100 серотипов. Типовой вид — риновирус А человека (74 серотипа, причем серотип 1 делят на 2 антигенных подтипа). Вид риновирус В человека включает 25 серотипов. Рецептором риновирусов является межклеточная адгезивная молекула I (ICAM-I), которая экспрессируется на эпителиальных клетках, фибробластах и эндотелиальных клетках. Риновирусы могут передаваться двумя механизмами: аэрозольным и контактно-бытовым. Вирусы проникают в организм через нос, полость рта, конъюнктиву. Процесс начинается в верхних дыхательных путях.
Вирус ящура — РНК-содержащий вирус рода Aphthovirus семейства Picornaviridae; вызывает ящур — зоонозную инфекционную болезнь, характеризующуюся лихорадочным состоянием, язвенными (афтозными) поражениями слизистой оболочки рта, кожи кистей и стоп. Вирус ящура по морфологии и химическому составу сходен с другими пикорнавирусами. Известны 7 серотипов вируса, включающих более 60 вариантов, которые различаются по антигенной структуре. Обладает высокой вирулентностью и дерматропностью. Вирус может длительно (несколько недель) выживать в объектах окружающей среды, в пищевых продуктах; чувствителен к дезинфектантам. Естественным резервуаром вируса ящура служат больные животные, крупный рогатый скот. От больных животных вирус выделяется с молоком, со слюной и мочой. Человек заражается при уходе за больными животными, а также при употреблении сырого молока и молочных продуктов. Восприимчивость человека к ящуру невысокая.
Репродукция вируса
Репродукция вируса – это процесс размножения вирусных частиц в чувствительных к ним клетках. Репродуцируются только вирулентные вирусы, обладающие высокой степенью патогенности [3] .
Термин «репродукция» восходит к латинскому слову «production» – производство [3] . Действительно, процесс размножения вирусов в корне отличается от бинарного деления клеток живых организмов. Он больше напоминает процесс, происходящий на заводском конвейере [4] . Процесс репродукции вируса внутри инфицированной клетки называют жизненным циклом вируса. Цикл репродукции у РНК-вирусов продолжается 4–8 часов, а у ДНК-вирусов – 12–24 часа [2] .
Содержание:
Общие закономерности репродукции вируса
К самостоятельному размножению вирусы не способны. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечиваются биосинтетическими процессами клетки-хозяина. Для вирусов характерен дизъюнктивный (разобщенный) тип репродукции. Он осуществляется при взаимодействии вируса с инфицируемой клеткой. В этом случае белковые молекулы и нуклеиновые кислоты образуются отдельно друг от друга. После чего происходит сборка дочерних популяций [3] .
Особенности репродукции вирусов зависят от типа вирусного генома. Однако отмечается существование целого ряда общих закономерностей репродукции вируса:
- Все вирусы, содержащие молекулу РНК, кроме вирусов гриппа и ретровирусов, репродуцируются в цитоплазме клетки. Геномы ретровирусов и вирусов гриппа при репродукции проникают в ядро клетки-хозяина [2] .
- Все вирусы, содержащие молекулу ДНК, кроме вирусаоспы репродуцируются в ядре и в цитоплазме клетки. В ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме – трансляция вирусных белков и сборка дочерних вирионов. Вирусоспы размножается только в цитоплазме клетки [2] .
- Процесс синтеза нуклеокапсидных белков происходит на свободных полирибосомах (не связанных с мембраной). Процесс синтеза суперкапсидных белков проходит на рибосомах, ассоциированных с мембранами [2] .
- Белки вирусов после образования подвергаются протеолитическому процессингу (разрезанию или расщеплению) [2] .
- Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране проходят гликозирование (присоединении к полипептиду углеводных остатков) [2] .
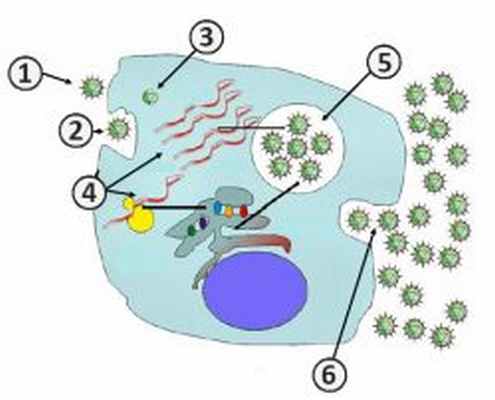
Этапы репродукции (жизненного цикла) вируса
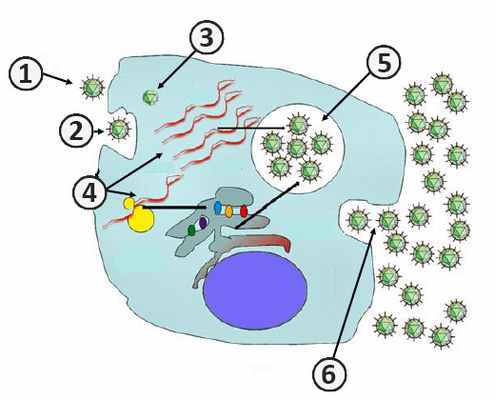
1. Адсорбция вируса на мембране клетки.; 2. Проникновение вируса в клетку.; 3. Депротеинизация.; 4. Синтез компонентов вирусов.; 5. Формирование дочерних вирионов.; 6. Выход вирионов [2] .
Этапы репродукции вируса
Репродукцию или жизненный цикл вируса делят на шесть последовательных этапов:
- Адсорбция на мембране клетки [2] .
- Проникновение в клетку [2] .
- Депротеинизация [2] .
- Синтез компонентов вирусов[2] .
- Формирование дочерних вирионов[2] .
- Выход вирионов[2] .
Процесс адсорбции вириона на поверхности клетки
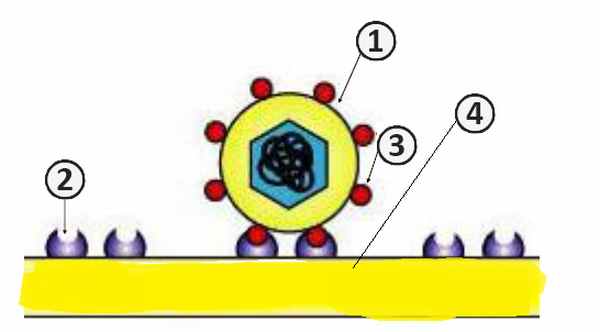
1. Вирион.; 2. Клеточный рецептор.; 3. Прикрепительный белок; 4. Клеточная мембрана [2] .
Адсорбция на мембране клетки
Адсорбция вириона на мембране клетки идет по пути взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вируса на клеточной мембране существуют специфические рецепторы, с которым он и связывается. По химической природе рецепторы, фиксирующие вирус, могут являться мукопротеиновыми либо липопротеиновыми. Распознавание клеточных рецепторов осуществляют капсидные или суперкапсидные белки вириона [2] .
Антирецепторы вирионов являются прикрепительными белками. Они могут иметь форму шипов, нитей, грибовидных структур [2] .
В самом процессе адсорбции большую роль играют электрические заряды. Вирусы обычно отрицательно заражены, а участки клеточной стенки – положительно [2] .
Процесс адсорбции занимает от пяти до девяноста минут. Количество специфических рецепторов на поверхности одной клетки 10 4 –10 5 [2] .
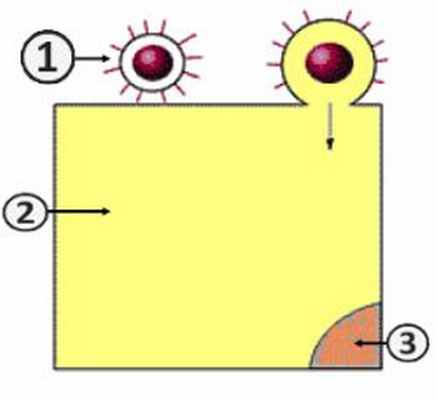
Проникновение вируса в клетку (Путь I)
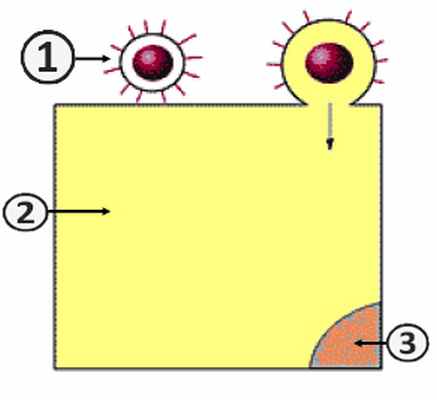
Слияние вирусной оболочки с клеточной мембраной
1. Вирион.; 2. Инфицируемая клетка.; 3. Ядро [2] .
Проникновение в клетку
Путь проникновения вируса в клетку зависит от наличия оболочки у вириона [2] . Существует два пути:
- Путь I – слиянии вирусной оболочки с клеточной мембраной [2] .
- Путь II – рецептор-опосредованный эндоцитоз [2] .
Путем слияния суперкапсида с клеточной мембраной (путь I) в клетку проникают оболочечные вирусы. Этот процесс обусловлен наличием специфических белков слияния. При этом наблюдается высвобождение нуклеокпсида в цитоплазму клетки [2] .
Путем рецептор-опосредованного эндоцитоза (путь II) в клетку проникают безоболочечные вирусы. Первоначально вирион связывается со специфическими рецепторами, расположенными на клеточной поверхности. Затем наблюдается инвагинация (впячивание) клеточной мембраны, образование эндосом (внутриклеточных вакуолей) и их слияние с лизосомами. В заключении, вирусный геном в цитоплазме клетки освобождается из эндосомы [2] .
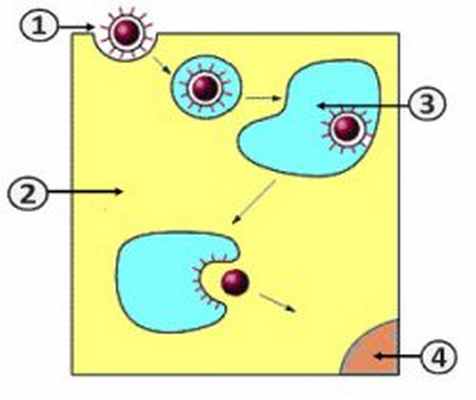
Проникновение вируса в клетку (Путь II)
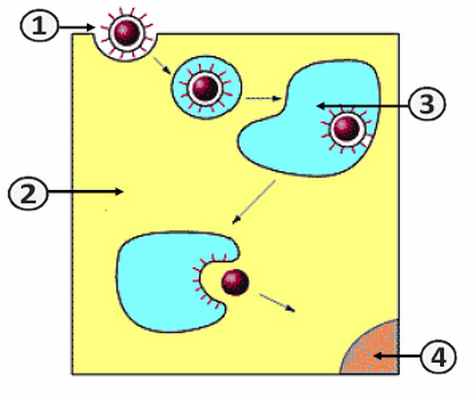
1. Вирион.; 2. Клетка.; 3. Эндосома.; 4. Ядро [2] .
Депротеинизация
Процесс депротеинизации (освобождения вирусной нуклеиновой кислоты – раздевание вируса) осуществляют протеолитические ферменты клетки (протеазы и липазы) [2] .
Смысл этого процесса состоит в удалении капсидов (вирусных оболочек). Конечные продукты раздевания вируса – сердцевины, нуклеокапсиды, нуклеиновые кислоты. Некоторые вирусы в качестве конечного продукта представлены нуклеиновыми кислотами, связанными с внутренним вирусным белком. После прохождения этапа депротеинизации выделить вирус из культуры клеток невозможно. Такое положение называют теневой фазой или фазой эклипса (затмения). В этот период вирус перестает существовать в качестве оформленного вириона [2] .
Синтез компонентов вируса
Синтез компонентов вирусов заключается в репликации вирусных нуклеиновых кислот и синтезе вирусных белков. Под репликацией понимается процесс самовоспроизведения нуклеиновых кислот, генов и хромосом, в основе которого лежит ферментативный синтез ДНК или РНК, проходящий по матричному синтезу [1] .
Место синтеза компонентов дочерних вирионов зависит от типа генома:
- реализация генетической информации у ДНК-содержащих вирусов идет по пути: ДНК → транскрипция → иРНК → трансляция → белок;
- реализация генетической информации у +РНК-содержащих вирусов идет без этапа транскрипции: +-РНК → трансляция → белок;
- реализация генетической информации у РНК-содержащих вирусов с негативным геномом идет по схеме: минус-РНК → транскрипция → иРНК → трансляция → белок;
- РНК-содержащие ретровирусы идут по следующему пути передачи информации: РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок [2] .
ДНК-содержащий вирус, проникший в цитоплазму, транспортирует нуклеокапсид к ядру клетки. Вирусная ДНК проникает в структуры клеточного ядра, где и совершается транскрипция или переписывание информации с ДНК на РНК при помощи клеточной полимеразы. Исключение – вирус оспы. Несмотря на то, что он относится к ДНК-содержащим, но его транскрипция протекает в цитоплазме при участии ДНК-полимеразы, проникающего в клетку в составе вириона [2] .
Результатом транскрипции является и то, что на одной из нитей ДНК синтезируется иРНК. В последствии, она перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках [2] .
Синтез белков наблюдается в рибосомах клетки-хозяина. Одновременно в ядре клетки протекает репликация (образование) дочерних нуклеиновых кислот на матрице материнской ДНК [2] .
Синтезированные дочерние молекулы ДНК в составе нуклеокапсида путем почкования перемещаются из ядра клетки в цитоплазму. При этом они захватывают фрагмент ядерной мембраны. В цитоплазме процесс репродукции завершается [2] .
Репликация «плюс»-РНК-содержащих вирусов происходит в цитоплазме. В данном случае функцию иРНК выполняет вирусная нуклеиновая кислота. В итоге трансляции на рибосомах формируется белковая молекула, которая разрезается клеточными протазами на структурные или неструктурные вирусные белки. Одновременно образуется полимераза, способствующая образованию «минус»-РНК на матрице родительской «плюс»-РНК. На матрице «минус»-РНК происходит синтез молекул «плюс»-РНК, принимающих участие в биосинтезе белков дочерних вирионов [2] [4] .
Вирусы, содержащие «минус»-РНК, отличаются тем, что их геном не способен выполнить функцию иРНК. Поэтому в цитоплазме клетки на матрице «минус»-РНК первоначально синтезируется «плюс»-РНК. Процесс катализируется полимеразой (транскрипазой), находящейся в составе проникшего в клетку вириона [2] [4] .
Синтез «плюс»-РНК приводит к образованию полных и коротких нитей. Короткие «плюс»-РНК–нити принимают участие в синтезе ферментов и бликов для дочерних популяций. Полные нити «плюс»-РНК служат матрицей для синтеза молекул «минус»-РНК дочерних вирионов. Последние транспортируются к клеточной мембране и высвобождаются, захватывая при этом ее фрагмент. Этот фрагмент служит для вирусной частицы суперкапсидом [2] [4] .
РНК-содержащие ретровирусы отличаются тем, что после проникновения в клетку генетическая информация с их РНК переписывается на ДНК, то есть с помощью фермента ревертаза происходит обратная транскрипция. Ревертаза так же попадает в клетку вместе с ретровирусом. Вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание информации с ДНК на РНК) осуществляет клеточная ДНК-зависимая РНК полимераза [2] [4] .
Формирование дочерних вирионов
Сборка дочерних вирионов возможна только при узнавании вирусных нуклеиновых кислот и белков, и самопроизвольном их соединении друг с другом. На мембранах эндоплазматического ретикулума взаимодействуют нуклеиновая кислота и белки просто устроенных вирусов, что приводит к образованию упорядоченной структуры [2] .
Сложно устроенные вирусы характеризуются многоступенчатой сборкой. Первоначально их нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды. Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под участками модифицированными оболочечными вирусными белками. В результате происходит самосброска вирионов. Количество зрелых вирионов, сформировавшихся в клетке, варьирует от 10 до 10000 и более [2] .
Выход вирионов
Высвобождение дочерних вирионов из клетки может быть осуществлено двумя способами:
- взрывной – путем лизиса клетки;
- путем почкования[2] .
Путь лизиса клетки тесно связан ее деструкцией. Он характерен для безоболочечных вирусов, не имеющих суперкапсидной оболочки (суперкапсида) [2] .
Выход путем почкования характерен для оболочечных вирусов. При этом клетка-хозяин некоторое время сохраняет жизнеспособность. Содержащие суперкасид вирусы, высвобождаются в течении 2–6 часов. В начале суперкапсидные белки устанавливаются на наружной поверхности мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсида [2] .
XII Международная студенческая научная конференция Студенческий научный форум - 2020

Размер вирусов определяют ультрафильтрацией, ультрацентрифугированием и электроноскопией [5].
Типы взаимодействия вируса с клеткой. Имеются два основных типа взаимодействия вируса и клетки. При первом типе вирусный геном функционирует в зараженной клетке более или менее автономно. Репродукция его происходит независимо от репродукции клеточного генома. Вирусы, автономно размножающиеся в клетке, относятся к группе вирулентных. При таком характере взаимодействия вируса и клетки образуется новое поколение вирионов. В этом случае говорят о продуктивном взаимодействии. Когда же цикл репродукции прерывается на какой-либо промежуточной стадии и инфекционного вирусного потомства не образуется, такое взаимодействие вируса и клетки называют абортивным. В тех случаях, когда симбиоз клеточного и вирусного геномов оказывается кратковременным и после образования нового поколения вирусных частиц зараженная клетка (клетка-хозяин) гибнет, такую реакцию на вирусную инфекцию называют литической. Явление, когда клетка, в которой автономно размножается вирус, длительно сохраняет свою жизнеспособность, получило название латенции.
Второй тип взаимодействия вируса и клетки свойствен опухолевым вирусам, нуклеиновая кислота которых способна встраиваться (интегрироваться) тем или иным образом в клеточную хромосому в форме провируса, вызывая трансформацию клеток. Границы между вирусами с автономной репликацией геномов и интеграционными вирусами весьма условны, и один и тот же вирус в зависимости от вида клеток может вести себя либо как инфекционный, либо как интеграционный геном. Результатом такого взаимодействия вируса и клетки является изменение наследственных свойств клетки. Этот тип взаимодействия вируса и клетки называют вирогенией, подобно лизогении при взаимодействии фагов с бактериями. Вирусы, способные вызывать вирогению, относят к группе умеренных[2].
Адсорбция вируса. Заражение клетки начинается с адсорбции вируса на клеточной мембране, происходящей благодаря взаимодействию поверхностных белков вируса с мембранными рецепторами.
Температура, как правило, мало влияет на адсорбцию (при 4ºС и при 37ºС скорость этого процесса практически одинакова).
Связывание с мембранными рецепторами не защищает вирусы от нейтрализации антителами [2].
Проникновение вируса в клетку. Адсорбированные вирусы проникают в клетку с помощью эндоцитоза или путем слияния с клеточной мембраной. Оказавшись в цитоплазме, вирусы освобождаются от большинства белков (раздевание) и начинают реплицироваться. Проникновение в клетку, раздевание и репродукция вирусов зависят от интенсивности энергетического метаболизма клетки и биохимических изменений, происходящих в клеточной мембране и цитоскелете.
Адсорбция многих вирусов запускает эндоцитоз, начинающийся с образования на мембране окаймленных ямок, покрытых клатрином. Затем формируются эндосомы, в составе которых вирусы поступают в цитоплазму. Данный способ проникновения в клетку характерен для пикорнавирусов, вирусов гриппа и аденовирусов. Последующее слияние вирусов с мембраной эндосом стимулируется понижением рН в эндосоме [5].
Сборка вирионов и выход их из клетки. В связи с разнообразием структуры вирусов животных разнообразны и способы формирования вирионов, однако можно сформулировать следующие общие принципы сборки
у просто устроенных вирусов формируются провирионы, которые затем в результате модификаций белков превращаются в вирионы. У сложно устроенных вирусов сборка осуществляется многоступенчато — сначала формируются нуклеокапсиды или сердцевины, с которыми взаимодействуют белки наружных оболочек;
сборка сложно устроенных вирусов (за исключением сборки вирусов оспы и реовирусов) осуществляется на клеточных мембранах. Сборка ядерных вирусов происходит с участием ядерных мембран, сборка цитоплазматических вирусов — с участием мембран эндоплазматической сети или плазматической мембраны, куда независимо друг от друга прибывают все компоненты вирусной частицы;
у ряда сложно устроенных вирусов существуют специальные гидрофобные белки, выполняющие функции посредников между сформированными нуклеокапсидами и вирусными гликопротеидами. Такими белками являются матриксные белки у ряда минус-нитевых вирусов (ортомиксо-, парамиксо-, рабдовирусов);
сборка нуклеокапсидов, сердцевин, провирионов и вири-онов происходит не во внутриклеточной жидкости, а в специальных структурах, предсуществующих в клетке или индуцированных вирусом («фабриках»);
сложно устроенные вирусы для построения своих частиц используют ряд элементов клетки-хозяина, например липиды, некоторые ферменты, у ДНК-геномного SV40 - гистоны, у оболочечных РНК-геномных вирусов — актин, а в составе арена-вирусов обнаружены даже рибосомы. Клеточные молекулы несут определенные функции в вирусной частице, однако включение их в вирион может явиться и следствием случайной контаминации, как, например, включение ряда ферментов клеточных оболочек или клеточных нуклеиновых кислот [1].
Сборка ядерных вирусов начинается в ядре, обычно — в ассоциации с ядерной мембраной. Формирующиеся в ядре Промежуточные формы вируса герпеса почкуются в перинуклеарное пространство через внутреннюю ядерную мембрану, и вирус приобретает таким путем оболочку, которая является дериватом ядерной мембраны. Дальнейшая достройка и созревание вирионов происходят в мембранах эндоплазм этической сети и в аппарате Гольджи, откуда вирус в составе цитоплазматических везикул транспортируется на клеточную поверхность.
У непочкующихся липидосодержащих вирусов (вирусов оспы) сборка вирионов происходит в уже описанных цитоплазматических «фабриках», Липидная оболочка вирусов в «фабриках» формируется из клеточных липидов путем автономной самосборки, поэтому липидный состав оболочек значительно отличается от состава липидов в клеточных мембранах [3].
Выход вирусных частиц из клетки. Существует два способа выхода вирусного потомства из клетки: путем взрыва и путем почкования. Выход из клетки путем взрыва связан с деструкцией клетки, нарушением ее целостности, в результате чего находящиеся внутри клетки зрелые вирусные частицы оказываются в окружающей среде. Такой способ выхода из клетки присущ вирусам, не содержащим липопротеидной оболочки (пикорна-, рео-, парво-, папова-, аденовирусы). Однако некоторые из этих вирусов могут транспортироваться на клеточную поверхность до гибели клетки.
Выход из клетки путем почкования присущ вирусам, содержащим липопротеидную мембрану, которая является дериватом клеточных мембран. При этом способе клетка может длительное время сохранять жизнеспособность и продуцировать вирусное потомство, пока не произойдет полное истощение ее ресурсов [4]
2. Сергеев В. А. Ветеринарная вирусология. - Москва, 2002.
3. Сергеев В.А., Непоклонов Е.А., Алипер Т.И. Вирусы и вирусные вакцины:учебник. 524 стр., 2007 г.
4. Филдс Б., Найп Д. Вирусология, 1989. - 1440 с.
5. Сюрин В.Н. и др. Ветеринарная вирусология/В.Н. Сюрин, Р.В. Белоусова Н.В. Фомина - 2-е над., перераб. и доп. - М.: Агропромиздат, 1991. 431 с, [8.] л, ил.: ил.
Читайте также: