Секреция протонов почками. Реабсорбция ионов бикарбоната почками
Добавил пользователь Валентин П. Обновлено: 27.01.2026
Холецистокинин стимулирует расслабление сфинктера Одди; увеличивает ток печёночной желчи; повышает панкреатическую секрецию; снижает давление в билиарной системе: вызывает сокращение привратника желудка, что тормозит перемещение переваренной пищи в двенадцатиперстную кишку. Холецистокинин является блокатором секреции соляной кислоты париетальными клетками желудка.
3. Почки, регулируя реабсорбцию и секрецию различных ионов в почечных канальцах, поддерживают их необходимую концентрацию в крови.
Реабсорбция натрия регулируется альдостероном и натрийуретическим гормоном, вырабатывающимся в предсердии. Альдостерон усиливает реабсорбцию натрия в дистальных отделах канальцев и собирательных трубочках. Секреция альдостерона увеличивается при снижении концентрации ионов натрия в плазме крови и при уменьшении объема циркулирующей крови. Натрийуретический гормон угнетает реабсорбцию натрия и усиливает его выведение. Выработка натрийуретического гормона возрастает при увеличении объема циркулирующей крови и объема внеклеточной жидкости в организме.
Концентрация калия в крови поддерживается за счет регуляции его секреции. Альдостерон усиливает секрецию калия в дистальном отделе канальцев и собирательных трубочках. Инсулин уменьшает выделение калия, увеличивая его концентрацию в крови, при алкалозе выделение калия увеличивается. При ацидозе - уменьшается.
Паратгормон паращитовидных желез увеличивает реабсорбцию кальция в почечных канальцах и высвобождение кальция из костей, что приводит к повышению его концентрации в крови. Гормон щитовидной железы тиреокальцитонин увеличивает выделение кальция почками и способствует переходу кальция в кости, что снижает концентрацию кальция в крови. В почках образуется активная форма витамина D, который участвует в регуляции обмена кальция.
Почки регулируют кислотно-щелочное равновесие, выделяя кислую или щелочную мочу. Выделение кислой мочи снижает общее количество кислых продуктов во внеклеточной жидкости, как и выделение оснований снижает их содержание в жидких средах организма.
В целом механизм, благодаря которому почки выделяют кислую или щелочную мочу, заключается в следующем: в просвет канальцев путем фильтрации непрерывно поступает большое количество ионов НСО3, выделение которых в конечную мочу удаляет основания из плазмы. Эпителиоциты выделяют в просвет канальцев также большое количество ионов Н+, таким образом удаляя их из крови. Если количество выделенных ионов Н+ превысит число ионов бикарбоната, попавших в первичную мочу, внеклеточная жидкость в сумме потеряет больше кислых продуктов.
И наоборот, если количество ионов НСО3, попавших в просвет канальца, превысит секрецию протонов, в сумме возникнет дефицит оснований.
Организм в сутки производит около 80 мэкв нелетучих кислот, источником которых главным образом является белковый метаболизм. Эти кислоты называют нелетучими, поскольку они, как Н2СО3, не способны выделяться легкими. Основной путь выведения этих кислот из организма состоит в выделении с мочой. Почки должны также препятствовать потере бикарбонатов с мочой. Выполнение этой задачи, с количественной точки зрения, более важно, чем выделение нелетучих кислот.
Ежедневно почки профильтровывают около 4320 мэкв бикарбоната (180 л/сут х 24 мэкв/л); в норме почти все это количество реабсорбируется канальцами, сохраняя таким образом главную буферную систему внеклеточной жидкости.
Далее будет рассмотрен вопрос о том, как реабсорбция бикарбоната и выделение протонов сопряжены с канальцевой секрецией ионов Н+. Поскольку бикарбонаты перед реабсорбцией связываются с протонами, образуя Н2СО3, необходимый объем секреции протонов в просвет канальцев должен составлять 4320 мэкв/сут.
Для освобождения организма от нелетучих кислот в мочу также должно быть выделено дополнительно 80 мэкв ионов Н+. Таким образом, в общей сложности секреция протонов в просвет канальцев составляет 4400 мэкв/сут.
При снижении концентрации ионов Н+ во внеклеточной жидкости (при алкалозе) почки теряют способность к реабсорбции бикарбонатов, прошедших через почечный фильтр; его выделение, таким образом, возрастает. Поскольку НСО3 в норме нейтрализует протоны во внеклеточной жидкости, такая потеря бикарбонатов равноценна добавлению ионов Н+ во внеклеточную жидкость. Следовательно, при алкалозе выведение НСО3 способствует приведению содержания протонов во внеклеточной жидкости к норме.
При ацидозе почки не выделяют бикарбонаты в мочу вследствие их полной реабсорбции из первичной мочи во внеклеточную жидкость. Происходит реабсорбция и новых ионов бикарбоната, образованных почками. В результате концентрация ионов Н+ во внеклеточной жидкости возвращается к норме.
Таким образом, почки регулируют содержание ионов Н+ внеклеточной жидкости благодаря трем основным механизмам: (1) секреции ионов H+; (2) реабсорбции бикарбонатов, попавших в первичную мочу, (3) образования новых ионов бикарбоната. Все эти процессы выполняются благодаря деятельности одной и той же системы, работа которой обсуждается в следующих разделах.
4. Через оптическую систему проходят не все лучи, которые исходят от предмета. Ограничение размера пучков лучей – результат совместного действия всех имеющихся в оптической системе диафрагм. Однако можно выделить одну (наименьшую) диафрагму, и считать, что остальные не ограничивают ход лучей. Такая диафрагма называется апертурной.
Апертурная диафрагма – это диафрагма, которая ограничивает размер осевого пучка, то есть пучка, идущего из осевой точки предмета.
Выходной зрачок – это изображение апертурной диафрагмы в пространстве изображений, сформированное последующей частью оптической системы в прямом ходе лучей.
Чтобы определить, какая из диафрагм оптической системы является апертурной, надо найти изображение всех диафрагм в пространстве предметов в обратном ходе лучей. Апертурная диафрагма – это диафрагма, изображение которой видно под наименьшим углом из осевой точки предмета. Если предмет находится на бесконечности, то апертурная диафрагма – это диафрагма, изображение которой имеет наименьшие линейные размеры.
Апертура определяет размер пучка лучей, входящего или выходящего из оптической системы. Передняя (задняя) апертура – это размер входного (выходного) зрачка. Числовая апертура – это произведение размера зрачка на показатель преломления.
Еще одной важной характеристикой оптической системы является положение входного (выходного) зрачка. Так как изображение чаще всего воспринимается или последующей оптической системой, или глазом, необходимо, чтобы выходной зрачок оптической системы совпадал с входным зрачком прибора или глаза по положению и размерам.
Для удаленного предмета или изображения входной (выходной) зрачок находится близко к оптической системе, поэтому положение зрачка ( или ) измеряется относительно оптической системы в обратных миллиметрах, то есть в килодиоптриях.
Для близкого предмета или изображения имеет значение положение зрачка относительно предмета или изображения, кроме того, расстояния от зрачка до прибора могут быть бесконечно велики (при телецентрическом ходе лучей), поэтому положение зрачка ( или ) измеряется в миллиметрах от предмета (изображения).
В системах, формирующих изображение дальнего типа, приемником изображения, как правило, является глаз. Для систем, работающих с глазом, выходной зрачок должен быть по размеру и положению согласован со зрачком глаза. При высокой освещенности объекта (в лабораторных приборах) диаметр зрачка глаза можно принять за равный 2 мм. Для приборов, работающих в условиях недостаточной освещенности, диаметр зрачка глаза считают равным 4 – 6 мм.
Глаз человека представляет собой сложную оптическую систему, которая состоит из роговицы, влаги передней камеры, хрусталика и стекловидного тела. Преломляющая сила глаза зависит от величины радиусов кривизны передней поверхности роговицы, передней и задней поверхностей хрусталика, расстояний между ними и показателей преломления роговицы, хрусталика, водянистой влаги и стекловидного тела. Оптическую силу задней поверхности роговицы не учитывают, поскольку показатели преломления ткани роговицы и влаги передней камеры одинаковы (как известно, преломление лучей возможно лишь на границе сред с различными коэффициентами преломления).
5. ысшая нервная деятельность – это процессы, происходящие в высших отделах центральной нервной системы животных и человека. К этим процессам относят условных и безусловных рефлексов, а также "высших" психических функций, которые обеспечивают адекватное поведение животных и человека в изменяющихся окружающих природных и социальных условиях. Высшую нервную деятельность центральной нервной системы следует отличать от работы центральной нервной системы по синхронизации работы различных частей организма между собой. Высшую нервную деятельность связывают с нейрофизиологическими процессами, проходящими в коре больших полушарий головного мозга и ближайшей к ней подкорке.
Термин "высшая нервная деятельность" впервые введён в науку И. П. Павловым, считавшим его эквивалентным понятию психическая деятельность. Все формы психической активности, включая мышление и сознание человека, Павлов считал элементами высшей нервной деятельности. Непрерывное совершенствование высшей нервной деятельности происходит в процессе обучения (усвоения чужого опыта), в результате которого человек приобретает способность выбирать наилучший из возможных вариантов, предвидеть результаты своей деятельности, изменять окружающие его условия, создавать новые, не имеющие аналогов материальные и духовные ценности, т. е. осуществлять психическую деятельность.
Реабсорбция бикарбонатов и секреция протонов. Подкисление мочи.
Одной из основных гомеостатических функций почек является поддержание концентрации ионов водорода (протонов Н + ). Все жидкости и ткани организма характеризуются определенным рН, что важно для процессов растворения, комплексообразования, нервно-мышечной проводимости, работы ферментов и т.д. В частности, рН артериальной крови поддерживается в достаточно узких пределах - 7,35-7,45, а для конкретного человека эти пределы еще уже. Однако нормальные физиологические процессы приводят к образованию 40-80 ммоль протонов при окислении аминокислот, неполном окислении энергетических субстратов (молочная кислота, кетокислоты) и др., которые в конечном итоге выводятся почками. Первый этап нейтрализации кислых продуктов осуществляется в плазме крови благодаря действию буферных систем. Буферные системы крови включают бикарбонатный буфер, гемоглобиновый, белковый, и, наконец, фосфатный буфер. Бикарбонатный буфер, представляющий основу буферного действия, состоит из слабой угольной кислоты, которая в соответствии с константой диссоциации частично диссоциирует на протон и ион бикарбоната, и ее натриевой соли.
Угольная кислота образуется при растворении в воде углекислого газа, а при участии фермента карбоангидразы может расщепляться на углекислый газ и воду.
Роль карбоангидразы в нашем организме очень велика. Практически все органы и системы, которые участвуют в поддержании кислотно-основного статуса или образовании кислых или основных продуктов, имеют этот фермент: легкие, эритроциты, почки, эпителий желудка и кишечника. Фермент позволяет регулировать содержание угольной кислоты, бикарбонатов и протонов. Содержание бикарбонатов в крови составляет 25 ммоль/л и занимает второе место после хлоридов (100 ммоль/л) по количеству анионов внеклеточной жидкости. Гемоглобиновый буфер оказывает свое действие через бикарбонатный, поэтому бикарбонатный буфер является основным. Роль фосфатного буфера в крови чрезвычайно мала, так как фосфаты - основные анионы внутриклеточной жидкости, их содержание в крови низко (около 1 ммоль/л) и буферная роль ничтожна. В моче же основным буфером является фосфатный, так как фосфаты выводится через почки в количестве до 70 ммоль/сутки.
На определенном этапе избыток углекислоты выводится легкими за счет увеличения числа дыханий в минуту (тахипноэ), однако протоны выводятся только почками.
Выведение протонов почками тесно связано с двумя другими процессами: реабсорбцией бикарбонатов и реабсорбцией натрия из почечного фильтрата. (ДАТЬ РИС.). Показано, что почечный эпителий непроницаем для бикарбонатов. Для того, чтобы профильтровавшиеся бикарбонаты, составляющие основу бикарбонатного буфера, возвратить в кровь, они должны соединиться с протоном и образовать угольную кислоту, которая в просвете канальца с помощью карбоангидразы расщепляется до воды и окиси углерода. Последняя диффузией поступает в клетки почечного эпителия, где при участии того же фермента образует через стадию угольной кислоты бикарбонаты. При избытке протонов или недостатке бикарбонатов почки могут образовывать дополнительное количество бикарбонатов из углекислого газа, полученного в результате метаболизма. Реабсорбция бикарбонатов сопряжена с реабсорбцией натрия и секрецией протона, так как натрий обменивается на протон. Секреция протона в свою очередь ограничена реабсорбцией натрия, и возможностями фосфатного буфера мочи, так как протон акцептируется фосфатом.
В сбалансированном состоянии бикарбонаты почти полоностью реабсорбируются, что сопровождается секрецией протонов и подкислением мочи. Первичная моча имеет такой же рН, что и плазма крови, т.е. она слабощелочная. При выходе из дистального канальца рН мочи снижается до 5,5-6,5 и она становится слабокислая. Процесс подкисления мочи с помощью указанного механизма наиболее характерен для кортикальных нефронов. В медуллярных нефронах выделение протона идет по схеме аммониогенеза.
При недостаточности указанной выше системы секреции протона, избытке протонов при ацидозе, а также у новорожденных детей до периода развития кортикальных нефронов, значительную роль в процессе выведения ионов водорода играет процесс выведения аммиака. В почках активируется фермент глутаминаза, который усиливает продукцию аммиака. Аммиак диффундирует в собирательные трубочки, где забуферивается протоном с образованием иона аммония. Собирательные трубочки для иона аммония непроницаемы, подкисления мочи не происходит, поэтому у новорожденных детей моча более щелочная.
При оценке вклада почек в регуляцию кислотно-основного состояния не следует забывать, что в дистальных канальцах осуществляется обмен иона натрия на протон и обмен иона натрия на ион калия, и процессы эти конкурирующие. Так, при гипокалиемии ион калия будет задерживаться в организме, а обмен иона натрия на протон усилится. В результате повышение реабсорбции натрия автоматически приведет к усилению реабсорбции бикарбонатов и защелачиванию крови – алкалозу, а также повышению уровня протонов в моче. Таким образом, при гипокалиемическом алкалозе пациент будет выделять кислую мочу.
Секреция протонов почками. Реабсорбция ионов бикарбоната почками
Секреция протонов почками. Реабсорбция ионов бикарбоната почками
а) Секреция протонов и реабсорбция ионов бикарбоната в почечных канальцах. Секреция протонов и реабсорбция ионов бикарбоната происходят практически во всех частях канальцевой системы, за исключением тонкого отдела петли Генле. В целом процесс реабсорбции бикарбоната по ходу тубулярного сегмента нефрона изображен на рисунке ниже.
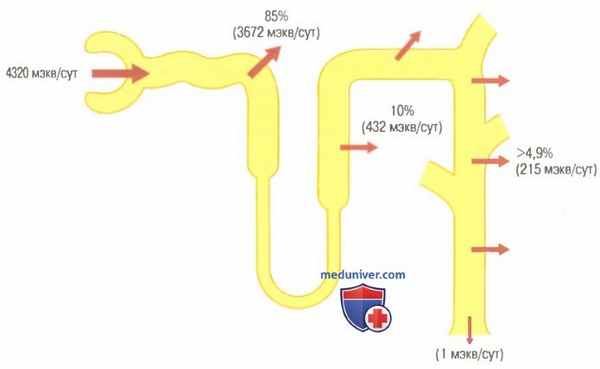
Реабсорбция бикарбонатов в различных отделах канальцевой системы нефрона, выраженная в процентах и миллиэквивалентах. Приведенные значения соответствуют нормальной реабсорбции за сутки
Следует помнить, что для реабсорбции каждого иона бикарбоната в просвет канальца необходимо секретировать один протон.
Около 80-90% реабсорбции ионов бикарбоната (и канальцевой секреции протонов) приходится на проксимальный каналец, поэтому лишь небольшое их количество поступает в дистальные сегменты нефрона. В толстом восходящем отделе петли Генле реабсорбируются другие 10% бикарбоната, прошедшего через почечный фильтр, а остаток поглощается дистальным канальцем и собирательной трубочкой. Как уже обсуждалось ранее, механизм реабсорбции бикарбонатов связан с канальцевой секрецией протонов, однако в различных сегментах канальцевой системы этот процесс протекает по-разному.
б) Секреция ионов водорода в проксимальных сегментах нефрона осуществляется благодаря вторично активному транспорту. Эпителиальные клетки проксимального канальца, толстого восходящего отдела петли Генле, начальных отделов дистальных канальцев секретируют протоны в просвет канальцев благодаря контртранспорту с ионами Na+ (для облегчения понимания просим вас изучить рисунок ниже).
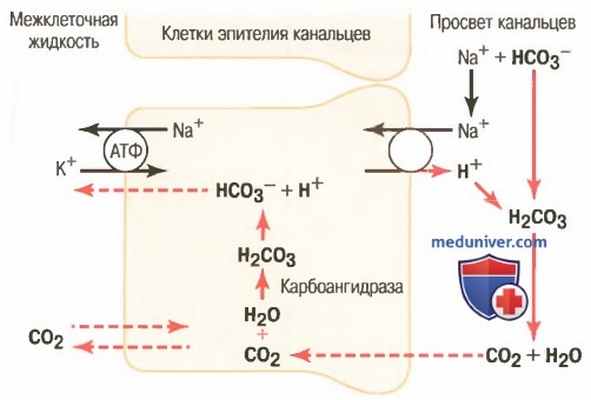
Внутриклеточные механизмы: (1) активная секреция протонов в просвет канальцев; (2) реабсорбция ионов бикарбоната в результате химического взаимодействия с протонами, образования угольной кислоты, которая разлагается на углекислый газ и воду; (3) реабсорбция ионов Na+ в обмен на протоны. Такой характер секреции ионов Н+ характерен для проксимальных канальцев, толстого восходящего отдела петли Генле и начальных отделов дистальных канальцев
Этот вторично активный механизм секреции связан с переносом ионов Na+ в клетку через апикальную мембрану с помощью белка, обменивающего натрий на протон.
Источником энергии для перемещения ионов Н+ в клетку служит производная градиента концентрации для Na+, который создается работой Na+/K+-насоса (АТФ-азы), расположенной на базолатеральной мембране эпителия канальцев. Таким способом реабсорбируется более 90% бикарбоната, что требует секреции в просвет канальцев ионов Н+ около 3900 мэкв/сут. Тем не менее, этот механизм не обеспечивает создание высокой концентрации ионов Н+ в моче: жидкость в просвете канальцев закисляется лишь в собирательных трубочках и каналах.
На рисунке выше показано, как при помощи процесса секреции ионов Н+ происходит реабсорбция бикарбоната. Секреция протонов начинается, когда СО2, попавший в клетку эпителия путем диффузии или в результате метаболических процессов, под влиянием фермента карбоангидразы соединяется с молекулой воды, формируя угольную кислоту. Угольная кислота диссоциирует на ионы Н+ и HCO3 - .
Протоны выделяются в просвет канальца с помощью противотранспорта с ионами Na+. Другими словами, перед тем как ионы Na+ переместятся через апикальную мембрану внутрь клетки, они вначале объединяются с мембранным белком-переносчиком. В то же время с другой стороны к белку-переносчику присоединяется ион Н+. Ион Na+ перемещается в клетку по градиенту концентрации, установленному АТФ-азой Na+/K+-насоса, расположенного на базолатеральной поверхности клетки. Энергия, затраченная на создание градиента концентрации, обеспечивает движение ионов Na+ внутрь клетки, а также обусловливает перемещение ионов Н+ в противоположном направлении: из клетки в просвет канальца.
Ионы HCO3 - , образованные в клетке при диссоциации молекулы H2CO3, затем пассивно перемещаются через базолатеральную мембрану в межклеточную жидкость почечной ткани и оттуда — через перитубулярные капилляры в кровь. Таким образом, на каждый выделенный в просвет канальца ион Н+ приходится один ион бикарбоната, реабсорбированный в кровь.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Секреция протонов почками. Реабсорбция ионов бикарбоната почками
Ткани и органы. Почки
Экскреция протонов и аммиака
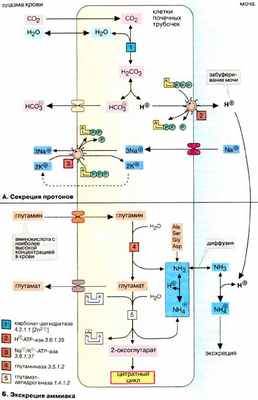
Почки и легкие играют основную роль в поддержании рН (гомеостаза) межклеточной жидкости в организме (см. рис. 281), причем почки вносят вклад в регуляцию кислотно-основного равновесия, осуществляя активную экскрецию протонов.
А. Секреция протонов
Клетки дистального отдела нефрона (извитого канальца и собирательных почечных трубочек) переносят протоны (H + ) из крови в просвет канальца (в мочу). Секреция идет против градиента концентрации, поскольку концентрация протонов в моче в 1000 раз превышает концентрацию в крови. При этом из крови в клетки почечных трубочек диффундирует диоксид углерода ( СО 2 ), который в цитоплазме гидратируется при участии карбонат-дегидратазы (карбоангидразы) [ 1 ] с образованием Н 2 СО 3 , диссоциирующей на ион бикарбоната ( НСО 3 - ) и протон. Протон секретируется из цитоплазмы в просвет канальца мембранной транспортной АТФ-зависимой системой [ 2 ], а ион бикарбоната всасывается через базолатеральную мембрану обратно в кровь. Для сохранения электронейтральности из канальца в кровь за счет реабсорбции переносятся ионы Na + . Суммарный процесс состоит в переносе протонов из крови в обмен на ионы Na + . Тем самым почки принимают участие в поддержании стабильного рН плазмы крови (равновесия СО 2 /НСО 3 - ) (см. рис. 281).
Ежедневно с мочой секретируется примерно 60 ммолей протонов. Однако в моче большая часть протонов нейтрализуется буферными системами, поэтому рН мочи лежит в слабокислой области (примерно до 4,8) Наиболее важной буферной системой является фосфатная (НРО 4 2- / Н 2 РО 4 - ). Определенный вклад в поддержание величины рН вносит аммиак за счет образования ионов аммония. В то время как экскреция фосфата зависит от количества фосфора, поступившего с пищей, выведение аммиака варьирует в широких пределах в зависимости от метаболических потребностей организма.
Б. Экскреция аммиака
Аммиак оказывает на клетки сильное токсическое действие. Основным путем обезвреживания аммиака в печени является образование мочевины (цикл мочевины, см. рис. 185). Главным источником аммиака в почках служит глутамин (уровень глутамина в крови составляет 0,5-0,7 мМ). Глутамин — один из конечных продуктов азотистого обмена, поступающий в кровь из мышц, головного мозга, печени и являющийся важнейшей транспортной формой аммиака в крови. В почках аммиак высвобождается из глутамина за счет гидролиза амидной группы [ 4 ] Вторая молекула аммиака образуется при окислительном дезаминировании глутамата с образованием 2-оксоглутаровой кислоты (см. рис. 181). Эта реакция катализируется глу таматдегидрогеназой [ 5 ] в присутствии НАД + или HАДФ + в качестве коферментов. Через 2-оксоглутарат этот процесс сопряжен с циклом лимонной кислоты. В качестве источника аммиака могут использоваться и другие аминокислоты, прежде всего аланин, а также серин, глицин и аспарагиновая кислота.
Аммиак диффундирует через клеточные мембраны в просвет канальца (в мочу), где соединяется с протонами, образуя соответствующую кислоту, ион аммония. В этой форме он уже не может реабсорбироваться мембранами клеток почечных трубочек и поэтому экскретируется в составе мочи. В сутки из организма выводится 30-50 ммолей аммиака.
При определенных изменениях обмена веществ выведение аммиака может быть полностью подавлено или существенно увеличено. Решающим фактором является величина рН крови, которая обычно составляет 7,4 (см. рис. 281). Если рН сдвигается в кислую область (ацидоз), выведение ионов аммония (аммиак + протон) усиливается. Это происходит за счет индукции синтеза глутаминазы, активность которой при ацидозе возрастает. То же происходит, например, при закислении организма за счет образования кетоновых тел при голодании и диабете. При сдвиге рН в щелочную область ( алкалоз ) выведение аммиака, напротив, подавляется.
Открытая медицинская библиотека
Статьи и лекции по медицине ✚ Библиотека студента-медика ✚ Болезни и способы их лечения.
Категории
Заболевания Реабсорбция бикарбонатов и секреция протонов. Подкисление мочи.
Одной из базовых гомеостатических функций почек является поддержание концентрации ионов водорода (протонов Н + ). Все жидкости и ткани организма характеризуются определенным рН, что важно для процессов растворения, комплексообразования, нервно-мышечной проводимости, работы ферментов и т.д. В частности, рН артериальной крови поддерживается в достаточно узких пределах - 7,35-7,45, а для конкретного человека эти пределы еще уже. При этом нормальные физиологические процессы приводят к образованию 40-80 ммоль протонов при окислении аминокислот, неполном окислении энергетических субстратов (молочная кислота͵ кетокислоты) и др., которые в конечном итоге выводятся почками. Первый этап нейтрализации кислых продуктов осуществляется в плазме крови благодаря действию буферных систем. Буферные системы крови включают бикарбонатный буфер, гемоглобиновый, белковый, и, наконец, фосфатный буфер. Бикарбонатный буфер, представляющий основу буферного действия, состоит из слабой угольной кислоты, которая в соответствии с константой диссоциации частично диссоциирует на протон и ион бикарбоната͵ и ее натриевой соли.
Угольная кислота образуется при растворении в воде углекислого газа, а при участии фермента карбоангидразы может расщепляться на углекислый газ и воду.
Роль карбоангидразы в нашем организме очень велика. Практически все органы и системы, которые участвуют в поддержании кислотно-основного статуса или образовании кислых или базовых продуктов, имеют данный фермент: легкие, эритроциты, почки, эпителий желудка и кишечника. Фермент позволяет регулировать содержание угольной кислоты, бикарбонатов и протонов. Содержание бикарбонатов в крови составляет 25 ммоль/л и занимает второе место после хлоридов (100 ммоль/л) по количеству анионов внеклеточной жидкости. Гемоглобиновый буфер оказывает свое действие через бикарбонатный, в связи с этим бикарбонатный буфер является основным. Роль фосфатного буфера в крови чрезвычайно мала, так как фосфаты - основные анионы внутриклеточной жидкости, их содержание в крови низко (около 1 ммоль/л) и буферная роль ничтожна. В моче же основным буфером является фосфатный, так как фосфаты выводится через почки в количестве до 70 ммоль/сутки.
На определенном этапе избыток углекислоты выводится легкими за счет увеличения числа дыханий в минуту (тахипноэ), однако протоны выводятся только почками.
Выведение протонов почками тесно связано с двумя другими процессами: реабсорбцией бикарбонатов и реабсорбцией натрия из почечного фильтрата. (ДАТЬ РИС.). Показано, что почечный эпителий непроницаем для бикарбонатов. Для того, чтобы профильтровавшиеся бикарбонаты, составляющие основу бикарбонатного буфера, возвратить в кровь, они должны соединиться с протоном и образовать угольную кислоту, которая в просвете канальца с помощью карбоангидразы расщепляется до воды и окиси углерода. Последняя диффузией поступает в клетки почечного эпителия, где при участии того же фермента образует через стадию угольной кислоты бикарбонаты. При избытке протонов или недостатке бикарбонатов почки могут образовывать дополнительное количество бикарбонатов из углекислого газа, полученного в результате метаболизма. Реабсорбция бикарбонатов сопряжена с реабсорбцией натрия и секрецией протона, так как натрий обменивается на протон. Секреция протона в свою очередь ограничена реабсорбцией натрия, и возможностями фосфатного буфера мочи, так как протон акцептируется фосфатом.
Читайте также: