Спинальные вегетативные рефлексы. Спинальный шок
Добавил пользователь Владимир З. Обновлено: 27.01.2026
В основе рефлекторной деятельности спинного мозга лежит рефлекс, структурно-функциональной основой которого является рефлекторная дуга. Различают моносинаптические и полисинаптические рефлекторные дуги.
Спинальные рефлексы подразделяют на соматические (двигательные) и вегетативные.
Двигательные рефлексы в свою очередь делятся на тонические (направлены на поддержание мышечного тонуса, поддержание конечностей и всего туловища в определенном статическом положении) и фазические (обеспечивают движение конечностей и туловища).
К тоническим относятся: миотатический рефлекс, шейные тонические рефлексы положения, рефлекс опоры (впервые их описание дал голландский физиолог Рудольф Магнус, 1924 г.), сгибательный тонический рефлекс.
К фазическим рефлексам относят: сухожильные рефлексы, рефлексы на укорочение с телец Гольджи, подошвенные, брюшные, сгибательные защитные, разгибательный перекрестный, ритмические.
Миотатический рефлекс – рефлекс растяжения, например, когда человек занимает вертикальное положение, то за счет гравитационных сил он может упасть (сгибание в суставах нижних конечностей), но с участием миотатических рефлексов это не происходит, т.к. при растяжении мышцы активируются мышечные веретена, которые расположены параллельно экстрафузальным волокнам скелетной мышцы. Импульсация от мышечных рецепторов идет через афферентный нейрон и попадает на альфа-мотонейроны данной мышцы. В результате происходит укорочение экстрафузальных водокон. Тем самым длина мышцы возвращается к исходной. Миотатический рефлекс свойственен всем мышцам, хорошо выражен и легко вызывается у мышц сгибателей, направлен против гравитационных сил, для поддержания равновесия, мышечного тонуса. Надо отметить, что импульсация от рецепторов одновременно через вставочные тормозные клетки Реншоу попадает на альфа-мотонейроны антагониста этой мышцы, поэтому при укорочении агониста мышца-антагонист не препятствует этому процессу.
Рецептивным полем шейных тонических рефлексов положения являются проприорецепторы мышц шеи и фасций, покрывающих шейный участок позвоночника. Центральная часть рефлекторной дуги имеет полисинаптический характер, т.е. включает вставочные нейроны. Рефлекторная реакция вовлекает мышцы туловища и конечностей. Кроме спинного мозга в ней участвуют и моторные ядра мозгового ствола, иннервирующие мышцы глазных яблок. Шейные тонические рефлексы возникают при поворотах и наклонах головы, что вызывает растяжение мышц шеи и активирует рецептивное поле рефлекса.
Рефлекс опоры (отталкивания) – при стоянии на поверхности усиливается тонус мышц разгибателей.
Сгибательный тонический рефлекс наблюдается, например, у лягушки или у кролика, при котором характерно подогнутое положение конечностей. Этот рефлекс направлен на поддержание определенной позы, что возможно при наличии определенного тонуса мышц.
Сухожильный рефлекс – рефлекс на укорочение с телец Гольджи
Подошвенный рефлекс – раздражение кожи стопы приводит к подошвенному сгибанию пальцев и стопы нижней конечности.
Брюшные рефлексы – напряжение брюшных мышц, возникающее при ноцицептивных афферентных влияниях. Это защитный рефлекс.
Сгибательные защитные рефлексы - возникают при раздражении болевых рецепторов кожи, мышц и внутренних органов направлены на избежание различных повреждающих воздействий.
Разгибательный перекрестный рефлекс: рефлекторное сгибание одной из конечностей нередко сопровождается сокращением контрлатеральной конечности, на которую в естественных условиях (при ходьбе) переносится дополнительный вес тела.
К ритмическим рефлексам у млекопитающих относится чесательный рефлекс. Его аналогом у земноводных является потирательный рефлекс. Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
Шагательный рефлекс – согласованная двигательная активность верхних и нижних конечностей. Для осуществления этого рефлекса необходимо межсегментарное взаимодействие мышц рук, ног и туловища. Механизмы шагательных движений заложены в спинном мозге, но включение спинального механизма производится со среднего мозга.
Вегетативные спинальные рефлексы: сосудистые, потоотделение, мочеиспускание, дефекация. Вегетативные рефлексы обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов.
5.4 Спинальный шок
Спинальный шок возникает после перерыва спинного мозга и представляет собой глубокое, но обратимое угнетение двигательных и вегетативных рефлексов, осуществляющихся ниже перерыва. Угнетение рефлексов связано с выпадением активирующей стимуляции со стороны головного мозга. У лягушек, у которых зависимость спинного мозга от головного выражена значительно меньше, чем у высших животных, спинальный шок длится несколько минут, у человекообразных обезьян и человека — несколько месяцев.
У человека полная арефлексия после перерыва спинного мозга является начальной стадией полной параплегии (двусторонний паралич ниже места перерыва спинного мозга). В последующем двигательные и вегетативные рефлексы постепенно восстанавливаются. Вначале появляются сгибательные рефлексы пальцев, имеющие характер патологических рефлексов (рефлекс Бабинского), после этого возникают более значительные и затем генерализованные спинальные рефлексы и движения типа спинальных автоматизмов. В хронической стадии появляются также разгибательные рефлексы, которые иногда переходят в разгибательные спазмы; больные с такими спазмами могут некоторое время стоять («спинальное стояние»). Все указанные рефлексы являются следствием растормаживания спинального локомоторного аппарата.
Подобные стадии, стадии угнетения и гиперактивации, характерны и для изменений вегетативных рефлексов при спинальном шоке. Сначала наступает полная вегетативная арефлексия, затем отмечается постепенное усиление симпатических рефлексов, переходящее в гиперрефлексию: в ответ на ноцицептивное раздражение возникают усиленное потоотделение и сужение сосудов.
Более быстрому восстановлению спинальных двигательных и вегетативных рефлексов способствует электростимуляция спинного мозга ниже (в ряде случаев и выше) перерыва. Такое воздействие благоприятствует выработке спинальных рефлекторных реакций при интероцептивных раздражениях с внутренних органов (возникновение рефлекторного мочеиспускания, рефлекторной дефекации и др.) и в какой-то мере восполняет афферентный дефицит.
5.5 Нарушения интегративной деятельности нервной системы
К типовым патологическим процессам в ЦНС относят образование новых патологических интеграций нейронов:
— генератора патологически усиленного возбуждения;
— патологической системы нервной регуляции.
Генератор патологически усиленного возбуждения — это агрегат гиперактивных нейронов, продуцирующий чрезмерный неконтролируемый поток импульсов (Г. Н. Крыжановский). В результате межнейрональных взаимодействий генератор приобретает способность развивать самоподдерживающуюся активность, которая не зависит от специальной стимуляции.
Возникновение генератора начинается либо с первичной гиперактивации нейронов, либо с первичного нарушения их торможения.
При первичной гиперактивации нейронов (значительная и устойчивая деполяризация), тормозные механизмы сохранены, но они функционально недостаточны. В этом случае имеет место вторичная недостаточность торможения, которая возрастает по мере развития генератора, при преобладании возбуждения. При первичной недостаточности тормозных механизмов в результате взаимодействия организма с этиологическим фактором, появляются растормаживание и вторичная гиперактивация нейронов.
Первичная гиперактивация нейронов возникает вследствие усиленных и длительных возбуждающих воздействий:
— при синаптической стимуляции. Например, формирование генератора в ноцицептивной системе: хронически раздражаемые рецепторы в тканях, эктопические очаги в поврежденных нервах, неврома (хаотически разросшиеся афферентные волокна) являются источником постоянной импульсации, под влиянием которой в центральном аппарате ноцицептивной системы формируется генератор;
— при действии возбуждающих аминокислот, в частности, глутаминовой (например, при ишемии);
— значительное увеличение в межнейрональном пространстве концентрации К + , который вызывает деполяризацию нейронов.
Первичное нарушение торможения нейронов формируется в условиях действия веществ, избирательно повреждающих тормозные механизмы:
— при действии столбнячного токсина, нарушающего выделение пресинаптическими окончаниями тормозных медиаторов;
— при действии стрихнина, блокирующие глициновые рецепторы на постсинаптических нейронах спинного мозга, где глицин оказывает тормозной эффект;
— при действии некоторых конвульсантов, нарушающих постсинаптическое торможение.
Патогенетическое значение ГПВУ состоит в том, что он обуславливает избыточную активацию тех отделов ЦНС, где локализованы популяции нейронов генератора патологически усиленного возбуждения. В результате данные отделы приобретают значение патологических детерминант, вызывающих образование патологических систем.
Специфика деятельности генераторов во многом определяет особенности соответствующих нейропатологических синдромов, имеющих генераторную природу. Действие первичного генератора служит причиной образования генератора вторичного, что способствует резистентности ПС. Кроме генераторов патологически усиленного возбуждения источниками патологической стимуляции других отделов могут становиться различные патологически измененные образования периферической или ЦНС.
Патологическая детерминанта — нерегулируемое гиперактивное образование ЦНС, которое, подчиняя другие связанные с ней отделы ЦНС посредством формирования патологической доминанты, определяет характер их деятельности, индуцируя возникновение ПС.
Гиперактивность детерминантной структуры обусловлена действующим в ней генератором. Патологическая детерминанта активируется модально-специфическими для нее стимулами (например, световыми, если она возникла в зрительной системе) либо стимулами, вызвавшими ее образование (последнее характерно для патологии ВНД и нервно-психических расстройств). Под влиянием первичной детерминанты могут появляться вторичные детерминанты либо в той же ПС либо в других физиологических системах. В последнем случае возникают новые ПС. В определенных условиях вторичная детерминанта может приобрести значение ведущего патогенетического механизма. Патологическая детерминанта является наиболее резистентным звеном патологической системы: при естественной ликвидации системы либо при ее фармакологическом подавлении патологическая детерминанта сохраняется дольше других частей и исчезает последней. При действии новых патогенных агентов, нарушающих механизмы тормозного контроля и усиливающих изменения, оставшиеся от детерминанты, последняя восстанавливается и играет важную роль в восстановлении ПС.
Доминанта, согласно определению А. А. Ухтомского, — это господствующая в данный момент активная функциональная структура ЦНС. Роль доминанты могут играть: активный нервный центр, констелляция центров, физиологическая система.
Существенной особенностью доминанты как функциональной физиологической структуры является сопряженное торможение других образований ЦНС, что и делает ее доминантой.
Сопряженное торможение происходит через:
— тормозящее влияние со стороны самой доминанты;
— избирательное торможение со стороны систем общего интегративного контроля в ЦНС, определяющего приоритетность той или иной реакции и ее доминирующее значение.
Патологию доминантных отношений составляют:
— недостаточность сопряженного торможения, что ведет к нарушению или невозможности образования физиологической системы, в т. ч. систем адаптации, компенсации и саногенеза;
— чрезмерное торможение, что ведет к нарушению или невозможности образования других физиологических систем и к выпадению функции ЦНС.
В обоих случаях нарушается интегративная и адаптивная деятельность нервной системы.
Крайним выражением генерализованного нарушения торможения являются общие судороги, возникающие при отравлении ядами (стрихнином, столбнячным токсином и др.). С недостаточностью тормозного контроля связаны судороги в завершающих стадиях терминальных состояний.
Патологическая система представляет собой новую (не существующую в физиологических условиях) патологическую интеграцию, возникающую из первично и вторично измененных образований ЦНС, деятельность которой имеет дизадаптивное или прямое патогенное значение для организма (Г. Н. Крыжановский). ПС формируется самой поврежденной нервной системой под влиянием гиперактивного образования ЦНС, приобретающего значение первичной патологической детерминанты.
К образованию ПС предрасполагают:
1. Ослабление тормозного контроля и рост возбудимости образований ЦНС, которые через влияния детерминанты вовлекаются в формирование ПС.
2. Недостаточность общего интегративного контроля ЦНС.
Образования и деятельность ПС являются как результатом, так и механизмом дальнейшего развития патологического процесса.
Примерами ПС в нервной системе могут служить: функционирование эпилептического очага, патологическая хроническая боль, артериальная гипертензия и язвенная болезнь, многие синдромы психических расстройств.
Спинальные вегетативные рефлексы. Спинальный шок
Спинальные вегетативные рефлексы. Спинальный шок
На уровне спинного мозга осуществляются многие типы сегментарных вегетативных рефлексов, большинство которых обсуждаются в других статьях на сайте (просим вас пользоваться формой поиска выше). К ним относят:
(1) изменения сосудистого тонуса в результате локального нагревания кожи;
(2) потоотделение в результате локального нагревания поверхности тела;
(3) кишечно-кишечные рефлексы, контролирующие некоторые моторные функции кишечника;
(4) брюшинно-кишечные рефлексы, тормозящие моторную активность желудочно-кишечного тракта в ответ на раздражение брюшины;
(5) эвакуационные рефлексы для опорожнения полного мочевого пузыря или переполненной толстой кишки.
Кроме того, все сегментарные рефлексы могут иногда возбуждаться одновременно в форме так называемого массивного рефлекса, изложенного далее.
Участки поверхности тела, где появляется отраженная боль от разных внутренних органов Висцеральная и париетальная передача болевых импульсов от аппендикса
а) Массивный рефлекс. Иногда у спинального животного или человека активность спинного мозга чрезвычайно повышается, что сопровождается массивным импульсным разрядом в значительной его части. Обычно это происходит в связи с действием сильного болевого раздражения кожи или чрезмерного переполнения внутренних органов, например при перерастяжении мочевого пузыря или кишки. Независимо от типа стимула возникающий в результате рефлекс, называемый массивным рефлексом, вовлекает большую часть или даже весь спинной мозг.
Эффекты представляют собой:
(1) мощный сгибательный спазм значительной части скелетных мышц тела;
(2) опорожнение прямой кишки и мочевого пузыря;
(3) часто подъем артериального давления до максимальных величин, иногда до уровней систолического давления, значительно превышающих 200 мм рт. ст.;
(4) обильное потоотделение обширных областей поверхности тела.
Поскольку массивный рефлекс может продолжаться в течение нескольких минут, он, вероятно, является результатом активации большого числа реверберирующих контуров, что одновременно возбуждает большие области спинного мозга. Это похоже на механизм развития эпилептических судорог, связанный с реверберацией возбуждения, возникающей в головном мозге вместо спинного.
Пересечение спинного мозга и спинальный шок
Когда спинной мозг внезапно пересекается в верхней части шеи, сначала практически все функции спинного мозга, включая спинномозговые рефлексы, мгновенно подавляются, вплоть до полного их выключения. Эту реакцию называют спинальным шоком. Причиной такой реакции является то, что нормальная активность спинальных нейронов зависит в большой степени от постоянного тонического возбуждения спинного мозга под действием импульсов, поступающих к нему по нисходящим нервным волокнам от высших центров, особенно по ретикулоспинальным, вестибулоспинальным и кортикоспинальным трактам.
В течение нескольких часов или недель возбудимость спинальных нейронов постепенно восстанавливается. По-видимому, это общее естественное свойство нейронов повсюду в нервной системе, т.е. после потери источника облегчающих импульсов нейроны увеличивают собственную естественную степень возбудимости, чтобы компенсировать потерю хотя бы частично. Большинству неприматов, чтобы возвратить к норме возбудимость центров спинного мозга, требуется от нескольких часов до нескольких суток. Однако у человека восстановление часто задерживается на несколько недель, а иногда полного восстановления совсем не происходит. В других случаях, наоборот, происходит чрезмерное восстановление с результирующей повышенной возбудимостью некоторых или всех функций спинного мозга.
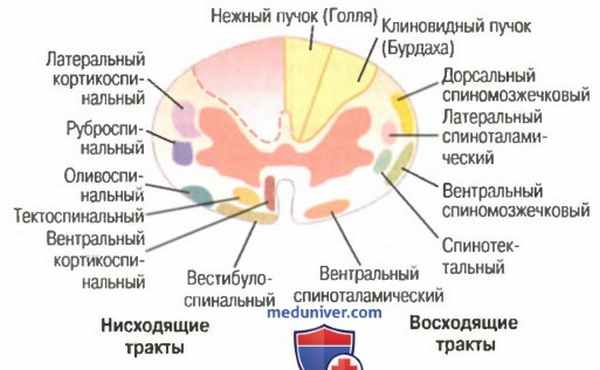
Поперечное сечение спинного мозга. Справа показаны главные восходящие тракты, слева — главные нисходящие тракты
Далее перечислены некоторые из спинальных функций, особенно страдающих во время или после спинального шока.
1. В начале спинального шока сразу и очень значительно падает артериальное давление, иногда опускаясь ниже 40 мм рт. ст., что свидетельствует о практически полной блокаде активности симпатической нервной системы. Давление обычно возвращается к норме в течение нескольких дней (даже у человека).
2. Все рефлексы скелетных мышц, интегрированные в спинном мозге, во время первых стадий шока блокируются. Животным для восстановления этих рефлексов до нормы требуется от нескольких часов до нескольких дней; людям — от 2 нед до нескольких месяцев. И у животных, и у человека некоторые рефлексы могут в итоге стать чрезмерно возбудимыми, особенно в тех случаях, когда на фоне пересечения основной части путей между головным и спинным мозгом некоторые облегчающие пути сохраняются. Первыми восстанавливаются рефлексы на растяжение, в дальнейшем постепенно восстанавливаются более сложные рефлексы в соответствующем порядке: сгибательные, антигравитационные позные и частично шагательные.
3. Рефлексы крестцового отдела спинного мозга, контролирующие опорожнение мочевого пузыря и прямой кишки, подавлены у человека в течение первых недель после пересечения спинного мозга, но в большинстве случаев они в итоге восстанавливаются.
Видео анатомия восходящих трактов спинного мозга (восходящих путей)
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Физиология человека и животных
3. Спинальные соматические и вегетативные рефлексы. Сегментарный и межсегментарный принцип работы спинного мозга
Рефлексы спинного мозга делятся на вегетативные и двигательные.
К вегетативным рефлексам относятся мочеиспускательный, дефекационный, потоотделительный, сосудистые рефлексы и др. К двигательным – кожно-мышечные, проприоцептивные и висцеромоторные рефлексы. По форме это в основном сгибательные и разгибательные рефлексы сегментарного характера.
Проприоцептивные рефлексы возникают при раздражении рецепторов со скелетных мышц и участвуют в формировании акта ходьбы и регуляции мышечного тонуса. В группу органных рефлексов входят рефлексы конечностей, брюшные, яичковый и анальный рефлекс. В свою очередь рефлексы конечностей могут быть сгибательные, разгибательные, ритмические и познотонические.
Сгибательные и разгибательные рефлексы делятся на фазные и тонические. Фазные рефлексы – это однократное сгибание или разгибание конечности при однократном раздражении кожи или проприоцепторов. Сгибательные фазные релексы – это локтевой, подошвенный, ахиллов и т.д., разгибательный фазный рефлекс – это коленный (рисунок 2). Тонические сгибательные и разгибательные рефлексы возникают при длительном сокращении или расслаблении мышц и направлены на поддержание позы.
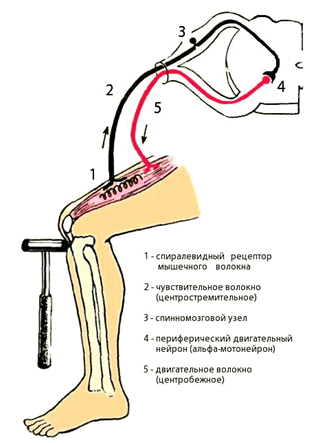
Рисунок 2 – Двухнейронная моносинаптическая
рефлекторная дуга коленного рефлекса
Рефлексы позы способствуют перераспределению мышечного тонуса при изменении положения тела или отдельных его частей. На уровне спинного мозга замыкаются шейные позные рефлексы, возникающие при наклоне головы или повороте головы. Эти рефлексы больше выражены у животных. Например, у кошки при наклоне головы вниз (кпереди) увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей. При наклоне головы вверх – наоборот. У человека эти рефлексы выражаются только в перераспределении мышечного тонуса для сохранения естественной позы тела. Вторая группа этих рефлексов возникает при поворотах головы вправо или влево. При этом увеличивается тонус мышц-разгибателей обеих конечностей на той стороне, куда повернута голова, и увеличивается тонус мышц-разгибателей конечностей на противоположной стороне тела. Эти рефлексы направлены на поддержание положения центра тяжести.
Универсальный принцип рефлекторной деятельности спинного мозга – это принцип общего конечного пути, открытый Ч.Шеррингтоном («воронка» Ч.Шеррингтона). Дело в том, что соотношение количества волокон в афферентных (задних корешках) и эфферентных (передних корешках) путях спинного мозга составляет примерно 5:1, поэтому широкую часть «воронки» составляют афферентные пути задних корешков, а узкую – эфферентные пути передних корешков спинного мозга.
Спинальный шок – это изменение мышечного тонуса и отсутствие рефлексов, наблюдающееся сразу после повреждения спинного мозга и связей его с вышележащими структурами ЦНС. Он развивается вследствие выключения влияния вышележащих отделов ЦНС на спинной мозг. Спинальный шок у лягушек длится несколько минут, у собак – несколько дней, у человека – около 2 месяцев. После исчезновения тормозного влияния вышележащих структур ЦНС резко увеличивается тонус мышц, иннервируемых с сегментов спинного мозга ниже места повреждения. Сгибательные и разгибательные рефлексы нижних конечностей резко повышаются. Это происходит вследствие повышения спонтанной активности γ-мотонейронов, приводящей к повышению спонтанной активности мышечных рецепторов и соответственно к повышению тонуса скелетных мышц.
Рефлексы вегетативной нервной системы
Нейроны вегетативной нервной системы участвуют в осуществлении многих рефлекторных реакций, называют вегетативными рефлексами. Последние могут спричинюватися раздражением как интеро-, так и екстерорецепторів. Критерий зачисления рефлекса к вегетативному - поступления импульсов к эфферентного периферического органа с ЦНС симпатичными или парасимпатичними нервами.
Рефлексы ганглиев вегетативной нервной системы. Рефлексы метасимпатичного отдела
Немало вегетативных ганглиев выполняют функцию вынесенных на периферию рефлекторных центров. Они имеют все структуры, необходимые для выполнения рефлекторного переключения. Не исключение и интрамуральные ганглии и нервные сплетения, имеющиеся в пустых органах. Эти ганглии - часть эфферентного пути парасимпатической нервной системы. Но вместе с тем к ним поступают нервные клетки от внутрішньоорганних рецепторов, есть здесь и вставные нейроны, следовательно, уже в самом ганглии возможна передача воздействий от рецепторного нейрона до эфферентного. Весомые аргументы в пользу наличия рецепторных нейронов в периферических нервных ганглиях выявлены факты сохранения афферентных, вставочных и эфферентных нейронов и нервных волокон, идущих от них, а также местной внутрішньоорганної рефлекторной регуляции в пересадженому сердце. Если бы эти рецепторы, нервные клетки и нервные волокна принадлежали к нейронам, тела которых находятся в ЦНС, то есть за пределами пересаженного сердца, должно произойти их перерождение.
Строение интрамуральных ганглиев напоминает типичные нервные центры. Каждый нейрон окружен большим количеством клеток нейроглії. К тому же здесь есть структуры, которые избирательно пропускают к нейрону из крови лишь определенные вещества, которые по своей функции напоминают ГЭБ. Таким образом, нейроны ганглия, подобно нейронам мозга, защищены от непосредственного воздействия веществ, циркулирующих в крови
Среди структур метасимпатичного отдела вегетативной нервной системы является пейсмекерні клетки, что обладают способностью к самопроизвольной деполяризации, которая обеспечивает ритм активности сокращение всех непосмугованих мышечных клеток органа. Эта активность корректируется импульсами собственной афферентации в зависимости от состояния органа, его отдельных частей.
"Местные" периферические рефлексы, которые выполняют интрамуральные вегетативные ганглии, регулирующие работу сердца, перистальтику кишечника, осуществляют взаимосвязь различных отделов желудка и некоторых других органов. Нейроны, входящие в эти ганглии, их отростки, синапсы и окончания формируют внутриорганные рефлекторные структуры, регулирующие работу органа внутрішньоорганними периферическими рефлексами.
Влияние парасимпатических нервных центров на метасимпатичні рефлексы.
Импульсы, поступающие к органу прегангліонарними волокнами парасимпатических нервов, вступают во взаимодействие с импульсами, осуществляющими процессы внутрішньоорганної рефлекторной регуляции. Характер ответной реакции органа определяет результат указанного взаимодействия. Поэтому эффект раздражения преганглионарных волокон не бывает однозначным. На органы, в которых обнаружены интрамуральные рефлекторные механизмы регуляции, прегангліонарні парасимпатические волокна могут оказывать (в зависимости от функционального состояния органа, который иннервируется) как возбуждающий, так и тормозное влияние.
Противоположные влияния парасимпатических волокон отнюдь не "парадоксальные". Это естественное проявление разнонаправленных воздействий, необходимых для обеспечения нормальной функции органов и тканей. Парасимпатичний отдел - это система, способная осуществлять текущую регуляцию физиологических процессов и обеспечивать в полном объеме поддержание постоянства внутренней среды организма. Количество интрамуральных нейронов, приходящихся на 1 см2 поверхности кишечника, может достигать 20 000. Как следствие, только одна часть метасимпатичної системы, что находится в кишках, содержит примерно такое же количество нейронов, что и весь спинной мозг.
Таким образом, импульсы, поступающие к органу прегангліонарними волокнами парасимпатических нервов, вступают во взаимодействие с импульсами, осуществляющими процессы внутрішньоорганної рефлекторной регуляции. В зависимости от текущего состояния физиологических процессов в этом органе или системе они могут включать или выключать, усиливать или ослаблять ту или иную функцию органа, осуществляя разнообразные регуляторные влияния, необходимые для поддержания нормальной текущей деятельности и гомеостаза.
Физиологическое значение "местных" рефлексов.
Эфферентные интрамуральные нейроны - это общий конечный путь для импульсов внутриорганного и екстраорганного (центрального) происхождения. Наличие "местных" механизмов нервной регуляции функций внутренних органов, что осуществляется с помощью периферических рефлексов ганглиями вегетативной нервной системы, внутрішньоорганними и позаорганними, имеет большое физиологическое значение. В результате этого ЦНС высвобождается от необходимости перерабатывать избыточную информацию, поступающую от внутренних органов. Кроме того, периферические рефлексы увеличивают надежность регуляции физиологических функций этих органов. Такая регуляция, будучи базовой, направленная на сохранение гомеостаза. В то же время она при необходимости может легко корректироваться высшими уровнями вегетативной нервной системы и гуморальными механизмами. К тому же эта регуляция может осуществляться и после выключения связи органов с ЦНС.
Спинальные рефлексы
На уровне спинного мозга замыкаются рефлекторные дуги многих вегетативных рефлексов (рис. 58).
Характер рефлекторного ответа во многом определяется наличием нервных центров симпатического (тораколюмбального) и парасимпатического (сакрального) отделов вегетативной нервной системы. Спинномозговой отдел симпатической нервной системы имеет признаки сегментарной (метамерної) организации. Это выражается в том, что четкое переключение чувствительных входов на эфферентные происходит в пределах конкретного сегмента. Хотя случаются и зоны перекрытия смежных сегментов, но в таком случае ответ на раздражение смежных корешков выражена в меньшей степени. Самые показательные в этом плане рефлексы сердечно-сосудистой системы и выделительных органов (кардио-кардиальные, желудочно-кишечные, евакуаторні рефлексы).
Інтернейронний аппарат спинного мозга обеспечивает взаимодействие рефлекторных путей как внутри вегетативной нервной системы, так и между ней и соматической нервной системой. Вследствие этого обеспечивается широкое привлечение различных внутренних органов в рефлекторную ответ. Важно и то, что рефлекс может запускаться с рецепторов одного, а заканчиваться эффекторами другого отдела нервной системы.
Спинальные центры регуляции вегетативных функций.
На уровне последнего шейного и двух верхних грудных сегментов спинного мозга находятся нейроны, иннервирующие три непосмуговані мышцы глаза: мышцу, расширяющую зрачок, глазничную часть круговой мышцы глаза и одну из мышц верхнего века.
В верхних грудных сегментах спинного мозга расположены нейроны, входящие в состав центра, который регулирует работу сердца и состояние кровеносных сосудов (см. разд. 3). Здесь есть нейроны, иннервирующие бронхи.
Во всех грудных и верхних поясничных сегментах спинного мозга расположены нейроны, иннервирующие потовые железы. Поражения отдельных сег
Рис. 58. Взаимодействие вегетативных и соматических путей спинномозговых рефлексов (по ногам): афферентные пути каждого нерва соматической нервной системы (1). вегетативного нерва (2), соматического рефлекса (3), вегетативного рефлекса (4)
ментов вызывает прекращение потоотделения в участках тела, что потеряли симпатическую иннервацию.
В крестцовом отделе спинного мозга находятся спинальные центры рефлексов мочеиспускания, дефекации, эрекции и эякуляции. Разрушение указанных центров вызывает половое бессилие, недержание мочи и кала. Нарушения мочеиспускания и дефекации происходит вследствие паралича мышц-замыкателей мочевого пузыря и прямой кишки.
Вегетативные рефлексы можно разделить на: вісцеро-висцеральные, вісцеродермальні и дерматовісцеральні.
Вісцеро-висцеральные рефлексы вызываются раздражением рецепторов, расположенных во внутренних органах, и заканчиваются изменением деятельности также внутренних органов. К тому же эти рефлексы могут начинаться и заканчиваться в органах одной функциональной системы (например сердечно-сосудистой) или быть межсистемными. К вісцеро-висцеральных рефлексов относятся рефлекторные изменения сердечной деятельности, тонуса сосудов, кровенаполнения селезенки вследствие повышения или снижения давления в аорте, каротидном синусе или легочных сосудах, рефлекторная остановка сердца при раздражении органов брюшной полости и др.
Вісцеродермальні рефлексы возникают при раздражении внутренних органов и проявляются в изменении потоотделения, электрического сопротивления (электропроводимости) кожи и кожной чувствительности на ограниченных участках поверхности тела, топография которых разнообразна в зависимости от того, какой орган раздражается.
Дерматовісцеральні рефлексы выражаются в том, что при раздражении некоторых участков кожи наступают сосудистые реакции и изменение деятельности определенных внутренних органов.
Многие из указанных вегетативных рефлексов применяют в практической медицине, к тому же применение их многоплановое.
Пример использования дерматовісцерального рефлекса в клинике - применение грелок или, наоборот, пузырей со льдом для воздействия на патологический очаг во внутренних органах. Лечебное воздействие разного типа акупунктур также основывается на подобных рефлексах. Вісцеродермальні рефлексы часто применяют в диагностике патологии внутренних органов. Так, развитие патологического очага в каком-либо внутреннем органе может усилить чувствительность определенных участков кожи, что проявляется их болезненностью при легком прикосновении или даже без раздражителя (отраженная боль в зонах Геда-Захарьина) (рис. 59). Подобный рефлекс может начаться с інтероцепторів, а ефектором могут стать скелетные мышцы: во время "пожара" в брюшной полости повы
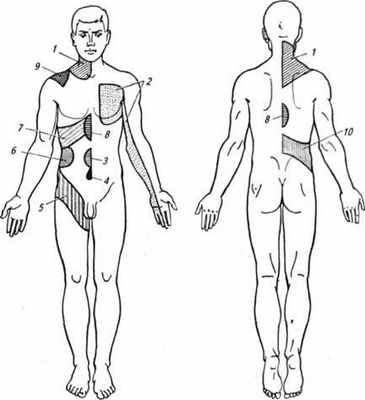
Рис. 59. Участка Геда-Захарьина, где отмечают повышенную чувствительность или отраженная боль при заболеваниях внутренних органов; 1-участок легких и бронхов; 2-область сердца; С - участок кишок; 4,5 - участок мочевого пузыря; б - участок почек; 7,9-участок печени; 8 - участок желудка и поджелудочной железы; 10 - участок мочевых и половых органов
щується тонус мышц-сгибателей (человек сворачивается "калачиком"), напрягаются мышцы определенных участков брюшной стенки.
Спинальный шок.
Указанные рефлексы спинного мозга в целостном организме координируются расположенными выше отделами ЦНС. Это четко проявляется после разрыва связь головного и спинного мозга. Вследствие такого повреждения, так же как и в соматической нервной системе, появляется спинальный шок - временное исчезновение вегетативных рефлексов спинного мозга. Исчезнувшие рефлексы постепенно, в течение 1-6 мес. восстанавливаются, причем даже такие сложные, как опорожнение мочевого пузыря и толстой кишки, половые.
Восстановление спинномозговых рефлексов после спинального шока может быть связано с активизацией бывших или формированием новых синапсов на вставочных преганглионарных и двигательных нейронах.
В этой ситуации дуги парасимпатических (вагусных) рефлексов не повреждаются.
Рефлексы ствола мозга
Вегетативные центры ствола мозга участвуют в регуляции функций сердечно-сосудистой, пищеварительной систем, осуществляющих евакуаторні рефлексы, управляют репродуктивными органами, контролируя иннервацию их вегетативными нервами. Здесь происходит объединение спинномозговых центров, отвечающих за отдельные вегетативные функции, в функциональные комплексы.
В продолговатом мозге размещен бульварный отдел вазомоторного центра, регулирующего функцию сердца и состояние сосудов. Здесь также содержатся центры, возбуждающие слезоотделение и секрецию слюнных и желудочных желез, поджелудочной железы, вызывают выделение желчи из желчного пузыря и желчного протока, стимулируют моторику желудка и тонкой кишки.
В среднем мозга (в передних буграх чотиригорбкової пластинки) содержатся нервные центры зрачкового рефлекса и аккомодации глаза. В передний участок среднего мозга располагается один из центров, которые участвуют в опорожнении мочевого пузыря. Указанные центры относятся к парасимпатического отдела. Но в целостном организме для выполнения рефлекторной функции многие из них (особенно это ярко проявляется на примере вазомоторного центра) тесно взаимодействуют с другими отделами ЦНС. Так, вазомоторный центр продолговатого мозга функционирует вместе с симпатическим отделом торакальной участка, а евакуаторні рефлексы осуществляются при взаимодействии центров ствола с крижовими центрами парасимпатической нервной системы. (Подробнее указанные рефлексы рассмотрены при изложении соответствующих разделов.)
Рефлекторная регуляция функций нервными центрами ствола осуществляется при непосредственном участии інтернейронних механизмов, которые отвечают за міжцентральну взаимодействие различных отделов ЦНС: симпатического, парасимпатического вегетативной и соматической нервной системы. Наглядный пример - дыхательно-сердечный рефлекс, или так называемая дыхательная аритмия: замедление сердечных сокращений в конце выдоха перед началом следующего вдоха.
Естественно, что все рефлексы ствола мозга находятся под контролем высших отделов ЦНС. К примеру, указанные выше евакуаторні рефлексы контролируются корой полушарий большого мозга.
В медицинской практике используются вегетативные рефлексы ствола мозга. Так, например, некоторые рефлексы, замыкающиеся здесь, дают возможность определить состояние вегетативной нервной системы (вегетативные функциональные пробы). К ним относятся: а) окосерцевий рефлекс, или рефлекс Даніні-Ашнера (кратковременное замедление сердцебиений при надавливании на глазные яблоки); б) ортостатическая реакция (учащение сердечных сокращений и повышение АД во время смены положения лежа на положение стоя) и др.
Читайте также: