Темновые реакции фотосинтеза. Опыты Кальвина.
Добавил пользователь Евгений Кузнецов Обновлено: 30.01.2026
Фотосистемы. Биохимия фотосинтеза. Источник кислорода.
Молекулы хлорофилла и вспомогательных пигментов расположены в фотосистемах, подразделяемых на два типа: фотосистемы I и II (ФС1 и ФС11). Эти фотосистемы можно обнаружить в виде частиц в составе тилакоидных мембран. Каждая из таких частиц содержит пигментные молекулы, организованные в так называемый антенный, или светособирающий комплекс.
В состав светособирающего комплекса входят 200-300 пигментных молекул, которые накапливают световую энергию, как это показано на рисунке. Различные пигменты улавливают свет с различной длиной волны, что делает этот процесс более эффективным. Вся энергия передается от молекулы к молекуле и, в конце концов, на специализированную форму хлорофилла о, известную как Р700 в ФСl и Р680 в ФСl. Р700 и Р680 — это пигменты (Р), у которых максимумы пиков в спектрах поглощения составляют соответственно 700 и 680 нм (оба пика в красной области спектра).
В результате поглощения энергии молекулы хлорофиллов Р700 и Р680 переходят в «возбужденное» состояние и становятся источниками электронов, обладающих высокой энергией (как описывалось выше). Судьба этих электронов будет обсуждаться ниже. Теперь мы можем рассмотреть суммарный процесс фотосинтеза.
Биохимия фотосинтеза
Для описания процесса фотосинтеза обычно используется следующее уравнение:
В таком виде уравнение удобно использовать, если надо показать образование одной молекулы сахара, однако это лишь суммарное отображение многих событий. Более подходящей формой записи является уравнение:
Соединения СН2О не существует в природе, это просто условное обозначение любого углевода.
Источник кислорода
Посмотрев на суммарное уравнение фотосинтеза, мы вправе задаться вопросом: какое сосдинение — диоксид углерода или вода — служит источником кислорода? Наиболее очевидным кажется ответ, что таким источником является диоксид углерода. Тогда для образования углевода оставшемуся углероду следует только присоединиться к воде. Точный ответ удалось получить в сороковых годах XX в., когда в распоряжении биологов оказались изотопы.
Обычный изотоп кислорода имеет массовое число 16 и обозначается как 16 O (8 протонов, 8 нейтронов). Существует еще редкий изотоп с массовым числом 18 ( 18 O). Это стабильный изотоп, но из-за большей, чем у 16 O , массы его можно обнаружить, используя масс-спектрометр, аналитический прибор, позволяющий выявлять различия между атомами и молекулами на основе значений их масс. В 1941 г. был проведен эксперимент, результаты которого суммированы в следующем уравнении:
Иными словами, источником кислорода является вода. В итоге уравновешенное уравнение выглядит как:
Это наиболее точное выражение процесса фотосинтеза, которое, кроме того, наглядно показывает, что вода не только используется при фотосинтезе, но и является одним из его продуктов. Данный эксперимент позволил заглянуть глубоко внутрь природы фотосинтеза, показав, что фотосинтез протекает в две стадии, первая из которых состоит в образовании водорода в результате расшепления воды на водород и кислород. Для этого требуется энергия, которую дает свет (поэтому процесс называют фотолизом: photos — свет; lysis — расщепление). Кислород высвобождается как побочный продукт. На второй стадии водород взаимодействует с диоксидом углерода, образуя сахар. Присоединение водорода — это пример химической реакции восстановления.
Тот факт, что фотосинтез является двухстадийным процессом, был впервые установлен в двадцатых—тридцатых годах XX в. Реакции первой стадии нуждаются в свете, поэтому они называются световыми реакциями. Реакции второй стадии света не требуют, поэтому они носят название темновых реакций, хотя и протекают на свету! Установлено, что световые реакции протекают на мембранах хлоропластов, а темновые реакции — в строме хлоропластов.
После того как установили, что темновые реакции фотосинтеза протекают вслед за световыми, в 1950-х годах оставалось только выявить природу этих реакций.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Фотосинтез
Фотосинтез — это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них — каротиноиды и фикобилины.
В природе распространены два пути фотосинтеза растений: C3 и С4. У других организмов есть своя специфика реакций. Все, что объединяет эти разные процессы под термином «фотосинтез», – во всех них в общей сложности происходит преобразование энергии фотонов в химическую связь. Для сравнения: при хемосинтезе происходит преобразование энергии химической связи одних соединений (неорганических) в другие — органические.
Выделяют две фазы фотосинтеза — световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы — наиболее распространенного продукта фотосинтеза:
Атомы кислорода, входящие в молекулу O2, берутся не из углекислого газа, а из воды. Углекислый газ – источник углерода, что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.
Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент — бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) - CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
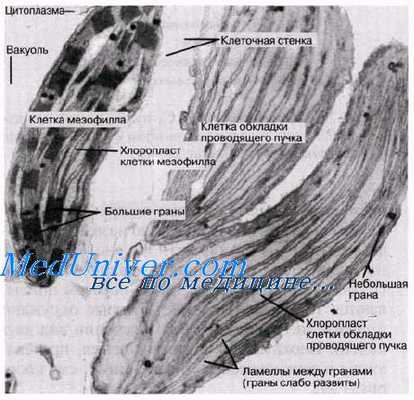
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Темновые реакции фотосинтеза. Опыты Кальвина.
С4-фотосинтез. Путь Хэтча-Слэка.
В 1965 г. было обнаружено, что первыми продуктами фотосинтеза у сахарного тростника (растение тропиков) являются органические кислоты, в состав которых входят 4 атома углерода (яблочная, щевелевоуксусная и аспарагиновая), а не 3С-кислота (фосфоглицериновая), как у большинства растений умеренного климата. С тех пор выявили множество растений, в основном тропических и субтропических, и имеющих порой важное экономическое значение, у которых фотосинтез протекает по такому же пути. Они получили название С4-растений. Примерами могут служить кукуруза, сорго, сахарный тростник и просо. Растения, в которых первым продуктом фотосинтеза является трехуглеродная фосфоглицериновая кислота, носят название С3-растения.
Биохимию именно С3-растений мы рассматривали до сих пор в этой главе.
В 1966 г. австралийские исследователи Хэтч и Слэк (Hatch, Slack) показали, что С4-растения значительно эффективнее, чем Сграстения, поглощают диоксид углерода: они способны удалять СО: из экспериментальной атмосферы вплоть до концентрации 0,1 части на миллион, тогда как для С3-растений это значение составляет 50—100 Ч*млн -1 . Хэтч и Слэк описали новый путь метаболизма углерода у Сграсгений, получивший название путь Хэтча—Слэка. Далее мы изучим этот процесс на примере типичного Сграстения — кукурузы.
У С4-растений листья имеют характерную особенность строения: вокруг каждого проводящего пучка у них расположены два ряда клеток. У клеток внутреннего кольца — обкладки проводящего пучка — хлоропласты по своей форме отличаются от хлоропластов клеток мезофилла внешнего кольца. Различие в строении хлоро-пластов у С4-растений получило название диморфизма (наличие двух форм). На рисeyrt, Ли Б показана так называемая «кранц-анатомия» (Kranz — корона, ореол; таким образом описываются те два ряда клеток, которые окружают проводящие пучки и имеют в сечении вид двух колец). Биохимические превращения, протекающие в этих клетках, описаны ниже; см. также рис. 7.22.
Путь Хэтча-Слэка
Путь Хэтча—Слэка предназначен для транспортировки диоксида углерода и водорода из клеток мезофилла в клетки обкладки проводящих пучков. Из клеток обкладки проводящих пучков диоксид углерода высвобождается и поступает в обычный С3-путь фотосинтетических превращений.
Захват (фиксация) диоксида углерода в клетках мезофилла
Диоксид углерода фиксируется в цитоплазме клеток мезофилла. Механизм этого процесса приведен в следующем уравнении:
Акцептором диоксида углерода служит фосфоенолпируват (ФЕП) вместо РиБФ у С3-растений, а вместо фермента РиБФ-карбоксилазы у С4-растений участвует фермент ФЕП-карбоксилаза. Фермент ФЕП-карбоксилаза работает значительно более эффективно, чем фермент С3-растений по двум причинам. Во-первых, ФЕП-карбоксилаза обладает большим сродством к диоксиду углерода, а во-вторых, ее работа не подвергается конкурентному ингибированию кислородом. Образовавшаяся щавелево-уксусная кислота далее превращается в малат, 4С-кислоту.
Малатный обходной путь (шунт)
Пройдя через плазмодесмы в клеточных стенках, малат попадает в хлоропласты клеток обкладки проводящего пучка, где он, соединяясь с диоксидом углерода, превращается в пируват (ЗС-кислоту). При этом выделяется водород, который используется для восстановления НАДФ. Обратите внимание, что в клетки мезофилла диоксид углерода и водород поступают извне, а затем в клетках обкладки проводящего пучка они вновь удаляются. Суммарным эффектом этих процессов является перемещение диоксида углерода и водорода из клеток мезофилла в клетки обкладки проводящего пучка.
Регенерация акцептора диоксида углерода
Пируват возвращается в клетки мезофилла, где в результате присоединения фосфатной группы от АТФ используется для регенерации ФЕП. На это расходуется энергия двух высокоэнергетических фосфатных связей.
- Вернуться в оглавление раздела "Биология."
Темновые реакции протекают в строме хлоропластов, не требуют света и контролируются ферментами. В результате этих реакций происходит восстановление диоксида углерода с использованием энергии (АТФ) и восстанавливающей способности (восстановленный НАДФ), произведенных в ходе световых реакций. Последовательность темновых реакций была определена в США Кальвином, Бенсоном и Бессемом в 1946—1953 годах. За эту работу в 1961 г. Кальвин был удостоен Нобелевской премии.
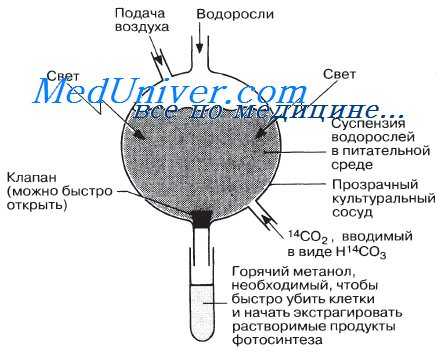
Схема, показывающая принцип устройства аппарата Кальвина типа «леденец на палочке». Он состоит из тонкого прозрачного сосуда, в котором выращивается культура одноклеточных водорослей. В экспериментах по определению пути, проходимому углеродом в процессе фотосинтеза, через суспензию водорослей продували диоксид углерода, содержащий радиоактивный углерод.
Опыты Кальвина
В своей работе Кальвин использовал изотоп углерода 14 С (период полураспада 5570 лет, см. приложение 1), который стал доступен для исследователей только в 1945 г.
Кроме того, Кальвин использовал бумажную хроматографию, которая в то время была относительно новым, но редко применяемым методом. Культуры одноклеточных зеленых водорослей Chlorella выращивали в популярном в настоящее время аппарате, по форме напоминающем леденец на палочке. Культуру хлореллы выдерживали с 14 С02 в течение различных промежутков времени, а затем быстро убивали, помещая в горячий метанол. Растворимые продукты фотосинтеза экстрагировали, концентрировали и разделяли при помощи двумерной бумажной хроматографии (рис. 7.16 и приложение 1). Цель данного эксперимента состояла в том, чтобы проследить маршрут, по которому меченый углерод через ряд промежуточных соединений включается в конечный продукт фотосинтеза.
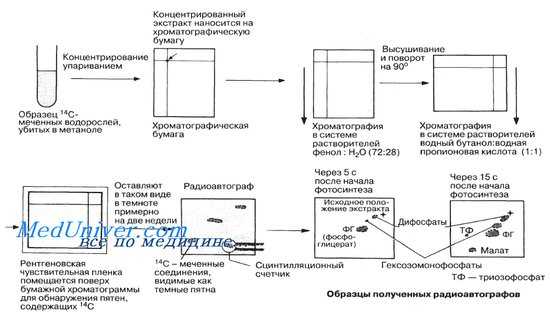
Идентификация продуктов фотосинтеза в водорослях после короткого периода освещения в присутствии радиоактивного диоксида углерода |4С02. Для разделения продуктов используется бумажная хроматография. Расположение соединений на бумаге позволяет их идентифицировать. Присутствие синтезированных веществ на хроматограмме выявляли при помощи фотопленки. Она темнела там, где находились радиоактивные соединения.
Соединения на хроматограмме обнаруживали при помощи радиоавтографии. Для этого использовалась фотопленка, чувствительная к излучению 14 С02 . Фотопленку накладывали на хроматограмму и она темнела там, где находились радиоактивные соединения.
Уже после 1 мин инкубации с 14 С02 синтезировались многие сахара и органические кислоты, в том числе аминокислоты. Используя краткосрочные выдержки (в течение 5 с и менее), Кальвину удалось идентифицировать первый продукт фотосинтеза — кислоту, содержащую 3 атома углерода (3С-кислота) — фосфоглицериновую (ФГ). В дальнейшем исследователь определил всю последовательность промежуточных соединений, в которые включается меченый углерод (стадии этого процесса будут рассмотрены ниже). С тех пор последовательность этих реакций носит название цикл Кальвина.
Темновая фаза фотосинтеза (пути ассимиляции углерода)
Темповая фаза фотосинтеза — это комплекс химических реакций, в результате которых происходит восстановление поглощенного листом С02, а также корнем NO3 и БО^за счет продуктов световой фазы (НАДФН, ФЛН2 и АТФ), и образуются органические вещества. НАДФН, ФЛН2, образовавшиеся в световой фазе и используемые на восстановление, получили название ассимиляционной силы, или восстановительных эквивалентов.
Цикл Кальвина
На рис. 4.15 представлен путь восстановления двуокиси углерода, названный циклом Кальвина по имени открывшего его американского биохимика М. Кальвина, получившего в 1961 г. за эти исследования Нобелевскую премию.
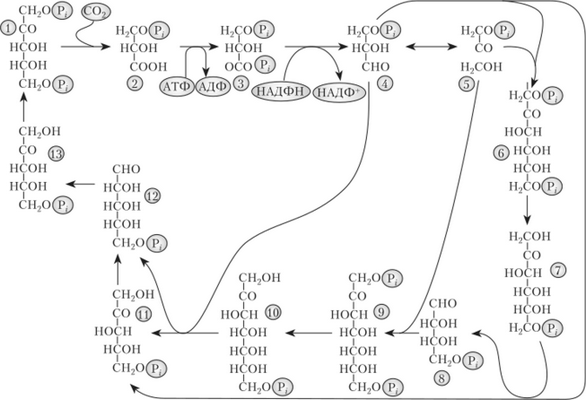
Рис. 4.15. Цикл Кальвина
Цикл начинается с присоединения С02 к рибулозо-1,5-бисфосфату (РуБФ). РуБФ является акцептором С02. Карбоксилирование катализируется рибулозобисфосфаткарбоксилазой (РуБФ-карбоксилазой; РуБисКО; RuBisCO). Это самый распространенный белок на Земле. Продукт реакции, содержащий шесть атомов углерода, распадается на две молекулы 3-фос- фоглицериновой кислоты (ФГК).
ФГК является первичным продуктом ассимиляции углерода. Так как в ее молекуле содержится три атома углерода, то этот цикл получил второе название — С3 -цикл. Для дальнейших превращений ФГК необходимы продукты световой фазы — АТФ и НАДФН. Прежде всего, ФГК фосфорили- руется при участии АТФ, и образуется 1,3-дифосфоглицериновая кислота (ДФГК) (см. рис. 4.15). Реакция катализируется фосфоглицераткиназой.
Эта кислота восстанавливается за счет НАДФН, и образуется 3-фос- фоглицериновый альдегид (ФГА), частично превращающийся с помощью триозофосфатизомеразы в фосфодиоксиацетон (ФДОА). Процесс восстановления ФГК до ФГА катализируется триозофосфатдегидрогеназой.
В результате остальных реакций цикла происходит регенерация РуБФ (см. рис. 4.15). С помощью альдолазы ФДОА соединяется с молекулой ФГА, и возникает молекула фруктозе-1,6-дифосфата (ФДФ).
Фруктозодифосфат дефосфорилируется и превращается во фруктоза - 6-фосфат (Ф-6-Ф), что сопровождается накоплением в строме хлоропласта неорганического фосфата. От фруктозо-б-фосфата отщепляется двухуглеродный фрагмент (— СО — СН2 ОН), который переносится на третью молекулу ФГА. Реакция идет при участии транскетолазы. В результате образуется пентоза — ксилулозо-5-фосфат, а от Ф-6-Ф остается 4-углеродистый сахар — эритрозо-4-фосфат (Э-4-Ф). Эритрозо-4- фосфат соединяется с молекулой ФДОА, и образуется семиуглеродистый сахар — седогептулозо-1,7-дифосфат (СДФ). После отщепления фосфата седогептулозодифосфат превращается в седогептулозофосфат (С-7-Ф). Затем от седогептулозофосфата отщепляется 2-углеродный фрагмент, который переносится натриозу. Образуются еще одна молекула ксилулозо- 5-фосфата и рибозо-5-фосфат (Р-5-Ф). Обе молекулы ксилулозо-5- фосфата и молекула рибозо-5-фосфата с помощью рибозофосфатизо- меразы и рибулозофосфат-3-эпгшеразы превращаются в три молекулы рибулозо-5-фосфата (Py-5-Ф). Последние фосфорилируются с помощью АТФ, образовавшихся во время световой фазы фотосинтеза, и возникают три молекулы рибулозо-1,5-бисфосфата (РуБФ). Эту реакцию катализирует фосфорибулокиназа. Таким образом, расходуются еще три молекулы АТФ. На этом цикл замыкается и может начаться снова. Рибулозо-фосфат и ФГА — конечные продукты этого цикла. Все остальные вещества, участвующие в нем, называют промежуточными. Поскольку в этом цикле образуются пентозы, то он имеет еще одно название — восстановительный пентозофосфатный цикл (ВПФ-цикл).
Цикл Кальвина можно представить в виде следующего суммарного уравнения:
Итак, на каждые три молекулы С02, вошедшие в цикл, образуется одна молекула ФГА и расходуются шесть молекул НАДФН и девять молекул АТФ.
Для простоты этот цикл делят на три этапа: 1) карбоксилирующий; 2) восстановительный и 3) регенерирующий. Первый этап — карбоксили-
рование рибулозобисфосфата и образование двух молекул фосфоглице- риновой кислоты (ФГК). Второй этап — восстановление ФГК с помощью продуктов световой фазы АТФ и НАДФН и образование двух молекул фосфоглицеринового альдегида (ФГА). Эта реакция — главная и является единственной реакцией восстановления во всем цикле. Третий этап включает несколько реакций, в результате которых образуется потраченная молекула рибулозо-1,5-бисфосфата. Для этого этапа также нужна АТФ, образовавшаяся во время световой фазы фотосинтеза.
Если цикл проходит три раза, то образуется шесть молекул ФГА. Пять из шести молекул триозофосфата используются для регенерации трех молекул РуБФ, а шестая молекула ФГА выходит из цикла и включается в синтетические процессы. Например, ФГА может использоваться для образования в хлоропласте первичного крахмала или, выйдя в цитозоль, для образования сахарозы или органических кислот.
Скорость С3-цикла зависит не только от количества образуемых во время световой фазы АТФ и НАДФН, но и от их соотношения. Только соотношение ЗАТФ к 2НАДФ обеспечивает активное восстановление С02 и запасание энергии. Если степень сопряжения работы электронтран- спортпой цени фотосинтеза с фотофосфорилированием мала, то интенсивность фотосинтеза снижается из-за уменьшения количества рибулозо-1,5- бисфосфата, так как в этом случае будет плохо идти фосфорилирование рибулозо-5-фосфата, уменьшается возможность восстановления ФГК до ФГА. В результате одновременно со снижением интенсивности фотосинтеза среди ассимилятов клетки возрастает доля неуглеводных соединений, например органических кислот. Такое явление характерно, например, для растений, выращиваемых при слабой освещенности. Цикл Кальвина происходит в строме хлоропласта.
Ночью в листе эти реакции не идут, так как наиболее важные ферменты цикла (РуБисКО, триозофосфатдегидрогеназа, рибулозо-5-фосфаткиназа, седогептулозо-1,7-бисфосфатаза и фруктозо-1,6-бисфосфатаза) активируются только при освещении. Активность этих ферментов регулируется также конечным продуктом.
Читайте также:
- Значение динамичных микротрубочек для клетки
- Поворотная, гиперкапническая пробы периорбитальной допплерографии. Нарушения мозгового кровообращения
- Лечение индийского лейшманиоза. Профилактика висцерального лейшманиоза
- Диагностика мицетомы. Лечение и профилактика мицетомы
- Полинейропатия: причины, симптомы и лечение