Трансдукция. Неспецифическая трансдукция. Специфическая трансдукция. Абортивная трансдукция. Феномен лизогении.
Добавил пользователь Владимир З. Обновлено: 27.01.2026
Длительное время считалось, что бактерии представляют собой изолированные генетические системы и каждая особь имеет одного (и только одного) родителя, т. е. их изменчивость обусловленалишь мутациями; никто не мог себе представить, что подобно гаметам высших организмов бактерии способны обмениваться генетическим материалом и, по аналогии с половым размножением, давать начало потомству с новыми свойствами.
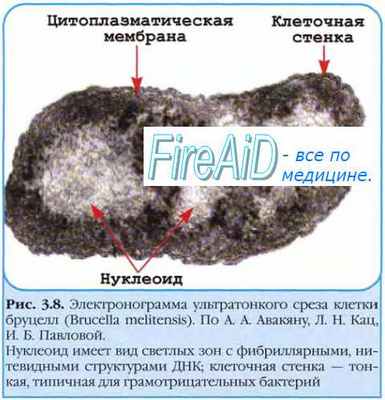
Прокариотам не свойственно половое размножение. Рекомбинация у них происходит в результате внутригеномных перестроек, заключающихся в изменении локализации генов в пределах хромосомы, или при проникновении в клетку реципиента части ДНК донора. Последнее приводит к формированию неполной зиготы - мерозиготы. В результате рекомбинаций в мерозиготе образуется только один рекомбинант, генотип которого представлен в основном генотипом реципиента с включенным в него фрагментом ДНК донора.
Генетические рекомбинации происходят при участии ряда ферментов в пределах отдельных генов или групп сцепленных генов. Существуют специальные rec-гены, детерминирующие рекомбинационную способность бактерий. Передача генетического материала (хромосомных генов) от одних бактерий к другим происходит путем трансформации, трансдукции и конъюгации.
Передача плазмидных генов - путем трансдукции и конъюгации.
Трансформация (изменение одного типа клеток при действии активного начала из другого типа клеток). Феномен открыл Гриффит у Streptococcus pneumoniae (1928); позднее Эвери, Маклеод и Мак Карти (1944) выделили трансформирующее начало пневмококков в форме молекулы ДНК. Это и явилось первым прямым доказательством того, что носителем генетической информации является ДНК. Погибшие бактерии постоянно высвобождают ДНК, которая может быть воспринята другими бактериями. Как правило, любая чужеродная ДНК, попадающая в бактериальную клетку, расщепляется рестрикционными эндонуклеазами; но при некоторых условиях такая ДНК может быть интегрирована в геном бактерий. По происхождению ДНК может быть плазмидной либо хромосомной и нести гены «трансформирующие» реципиента. Подобным путем процессы трансформации могут распространять гены, кодирующие факторы вирулентности, среди бактериальных популяций; однако в обмене генетической информацией трансформация играет незначительную роль.
Трансдукция. Явление открыл Ледерберг и Циндер (1952); в ходе трансдукции ДНК (но не нуклеопротеид) переносится от одной клетки (донор) к другой (реципиент) с помощью бактериофагов; обычно при этом фаг переносит лишь небольшой фрагмент ДНК хозяина. Выделяют 3 типа трансдукции: неспецифическую (общую), специфическую и абортивную. Неспецифическая трансдукция - в клетке, инфицированной бактериофагом, в ходе сборки дочерней популяции в головки некоторых фагов вместе с вирусной ДНК может проникнуть фрагмент бактериальной ДНК или плазмиды. При этом фаговая частица может утратить часть своего генома и стать дефектной. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены. Специфическая трансдукция - характеризуется способностью фага переносить определенные гены от бактерии-донора к бактерии-реципиенту. Это связано с тем, что образование трансдуцирующего бактериофага происходит путем выщепления профага из бактериальной хромосомы вместе с генами, расположенными на хромосоме в клетке-донора рядом с профагом. При взаимодействии трансдуцирующих фагов с клетками реципиентного штамма происходит включение гена бактерии-донора вместе с ДНК дефектного фага в хромосому бактерии-реципиента. Бактерии, лизогенированные дефектным фагом, невосприимчивы, как и все лизогенные клетки, к последующему заражению гомологичным вирулентным фагом. Абортивная трансдукция - принесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому бактерии-реципиента, а располагается в ее цитоплазме и может в таком виде функционировать. Во время деления бактериальной клетки трансдуцированный фрагмент ДНК-донора может передаваться только одной из двух дочерних клеток, т. е. наследоваться однолинейно и постепенно утрачиваться.
Конъюгация - перенос генетического материала их клетки-донора в клетку-реципиента при их скрещивании. Процесс конъюгации у бактерий впервые обнаружен Д. Ледербергом и Э. Тейтумом в 1946 г. Позднее выяснилось, что донорами генетического материала являются клетки, несущие F-плазмиду (половой фактор). При скрещивании F + c F - клеткой половой фактор передается независимо от хромосомы донора, если плазмида находится в автономном состоянии. При этом почти все реципиентные клетки получают F плазмиду и становятся F + клетками. Первым этапом конъюгации является прикрепление клетки-донора к реципиентной клетке с помощью половых ворсинок (sex pili). Затем между обеими клетками образуется конъюгационный мостик, через который из клетки-донора в клетку-реципиент могут передаваться F-фактор и другие плазмиды, находящиеся в цитоплазме бактерии-донора в автономном состоянии.
Интеграция F-плазмиды в состав бактериальной хромосомы приводит к разрыву одной из нитей ДНК, что обеспечивает возможность переноса в реципиентную клетку. Такие штаммы бактерий-доноров получили название Hfr-штаммы (с англ. - высокая частота рекомбинаций). При скрещивании Hfr-штамма с F - бактериями F-фактор, как правило, не передается, поскольку он расположен в дистальной части хромосомы.
Что такое генетическая трансдукция
Генетическая трансдукция — это перенос ДНК бактерий из одной клетки в другую с помощью бактериофага. В этом процессе ведущая роль принадлежит вириону, который является трансдукционной частицей. В результате происходит изменение наследственных признаков.
Генетическая трансдукция — процесс переноса бактериальной ДНК из одной клетки в другую.
История исследования
Процесс перемещения ДНК был установлен американскими исследователями Д. Ледербергом и Н. Циндером в 1952 г. Ученые изучали функции и строение бактерии Salmonella typhimurium и фага Р22. В результате проведенных опытов они пришли к выводу: происходит генетический обмен между микроорганизмами, не способными к синтезу соединений, и прототрофами.
Между встраивающимися генами не наблюдается физического контакта, процесс не идентичен трансформации. Открытие ученых оказалось случайным, один из штаммов был подвергнут состоянию дезорганизации благодаря вмешательству вириона Р22.
В ходе эксперимента в клетке наблюдали вегетативный рост профага. Через некоторое время образовавшиеся частицы фага проникали в ауксотрофный вид и переносили в его клетки гены своего прежнего хозяина.
Реакции фагов
Вирионы, избирательно уничтожающие бактериальные клетки, перемещают генетический материал от донора, в котором размножался возбудитель, к реципиенту, зараженному фагом. Вирус Х переносит лишь избранные гены — gal и Ыо. Носитель вирусных частиц проникает в фазу генома фага, внедренную в хромосомную часть ДНК зараженной клетки.
Вирионы перемещают генетический материал от донора к реципиенту.
Вирусная ДНК отделяется от бактериальной нуклеопротеидной структуры, несущей наследственную информацию. Возможно ошибочное деление вирусной ДНК. В этом случае ее фрагмент сохранится в нуклеопротеиде, но гены основной зиготы будут уничтожены. Главная причина появления неточности — неправильное перераспределение генетического материала.
Фаги уничтожают бактериальную структуру, используя литический или лизогенный метод,
Генетика фагов изучает мутации вирионов Та и Ти, а также физические и химические свойства фагов F1 и I2. Мутанты вирионов чувствительны к колебаниям температуры. К ним принадлежат фаг ts и холодочувствительный вирус es. Они размножаются и разрушают клетки при высокой температуре.
Литическая
Клеточный процесс сложный. Вирионы Т (Т2, Ти, Т6) содержат в ДНК вещество 5-гидроксиметилцитозин. Измененные формы фага способны к репродукции за счет заблокированных фаз. В вирионах-мутантах различные части расположены в четкой последовательности.
Фаги не способны двигаться. В результате взаимодействия вириона и взвеси бактерий частицы фага после столкновения с клетками остаются на их поверхности и вводят в структурную единицу бактерии свою ДНК.
Через несколько часов часть клеток разрушается. Вирусы, участвующие в литической фазе, становятся вирулентными.
На последней стадии литического процесса протеиновые частицы окружают нуклеиновую кислоту, формируя новые фаги.
Лизогенная
Умеренные вирусы взаимодействуют с микроорганизмами. Нуклеиновая кислота, находящаяся внутри бактериальной структурной единицы, внедряется в геном. В течение длительного периода наблюдают сосуществование вируса и клетки, не сопровождающееся гибелью структурной единицы.
Вирусы взаимодействуют с микроорганизмами.
При изменении условий среды вирус проникает в клетку. Начинается развитие активной стадии заболевания. Клетка, содержащая геном вириона, встраивающийся в хромосомную ДНК бактерии, называется лизогенной. Процесс сопровождается мутацией наследственной структурной единицы.
Умеренный фаг обладает способностью к заражению, которая определяется ДНК-связывающими свойствами белка. Лизогения меняет наследственные признаки бактериальной клетки.
Трансдукционное картирование
При переносе бактериальной ДНК вирионы размножаются на чувствительных клетках донора. Для мутагенного ДНК-связывающего белка, содержащего фрагмент нуклеопротеида и несущего наследственную информацию, проводят идентификацию донора и хозяина по нескольким признакам. Для установления сцепления используют отборное скрещивание.
Вирус размножают на штамме, имеющем фенотип Itr+Leu+, и заражают им клетки хозяина, которые нуждаются в аминокислотах.
После обработки вирионом структурные единицы организма помещают в питательную среду, не содержащую треонина. В питательной смеси не происходит рост гибридной нуклеиновой кислоты, но образуются структурные единицы, получившие часть хромосомы донора с многофакторным наследственным материалом.
На этом же участке может располагаться второй ген, встречаются двойные Iht+Leu+. Их можно обнаружить, поместив колонии трансдуктантов в питательную среду, не содержащую лейцина. Аминокислоту используют как неселективный отборный материал, необходимый для установления котрансдукции.
Общая (неспецифическая) трансдукция
Вирус Р1, находящийся в структурной единице хозяина в виде плазмиды, а также фаги Р22 и Ми осуществляют неспецифическое перемещение бактериальной ДНК. Вирусы Р22 и Ми встраиваются в любой участок нуклеопротеидной структуры инфицированной зиготы.
Вирусы осуществляют неспецифическое перемещение бактериальной ДНК.
Усиление генной структуры фага сопровождается проникновением части ДНК бактерии в его структуру. ДНК вириона в капсиде отсутствует. Величина этого участка равна длине стандартной вирусной ДНК.
Проникая в иную клетку, часть ДНК попадает в состав генетического материала после соответствующей рекомбинации. Перенесенные вирусом небольшие молекулы ДНК образуют кольцо и подвергаются репликации в новой бактериальной структурной единице.
События в клетке реципиента
Частицы, появившиеся в результате трансдукции, доставляют ДНК в клетки бактерий. Инфекционный процесс не развивается, т.к. наследственные факторы, отвечающие за воспроизведение фага, отсутствуют.
ДНК, принадлежащая донору, придерживается одного из двух путей развития: прикрепляется к хромосоме или остается в цитоплазме. Если часть ДНК не способна к процессу перекодирования, через некоторое время она разрушается.
Иногда отрезок ДНК удваивается самостоятельно и передается в другие поколения как носитель наследственных признаков, находящихся вне хромосомы.
Вирусы Р22 и Р1 принимают участие в двухниточной замене частей бактериальной хромосомы на вирусную ДНК. В клетку реципиента в процессе трансдукции попадает лишь 5% всей нуклеиновой кислоты.
Вирусы принимают участие в замене частей бактериальной хромосомы.
Использование
Практическое применение вирусов основано на литическом воздействии фагов на бактерии. Вирион используют в следующих случаях:
- для лечения инфекционных болезней;
- с целью идентификации микроорганизмов, вызывающих холеру, сальмонеллез, дифтерию.
Вирионы используют для изучения генетики микроорганизмов. Лечение заболеваний антибиотиками не всегда приводит к желаемому результату, поэтому применяют фаготерапию.
Вирионы применяют в сельском хозяйстве, животноводстве, растениеводстве.
Бактериальные фаги имеют следующие достоинства: не влияют на рост микрофлоры живого организма, уничтожают устойчивые к лечению АБ патогенные микроорганизмы, не оказывают воздействия на формирование эукариотических клеток.
Специфическая трансдукция
Ученые изучили перенос бактериальной ДНК вирусом Л. Фаг располагается на att-участке нуклеопротеидной структуры Е.coli, имеющей одинаковую последовательность фосфорных эфиров.
В период выведения вириона его исчезновение часто сопровождается ошибкой: удаляется часть, по размеру равная фрагменту ДНК. Особенность процесса состоит в том, что начало обломка располагается в другом месте. Некоторые гены вируса исчезают, другие наследственные факторы, несущие информацию, поглощаются фагом.
Некоторые фаги внедряются в любую часть нуклеопротеиновой структуры и переносят наследственную информацию, используя механизм специфического перемещения бактериальной ДНК из одной клетки в другую.
В хромосоме находится пространство, имеющее общие черты с att-участком и ДНК вируса. При разрушении гомологичного ресурса осуществляют внедрения фага в нуклеопротеид и его передачу при специфическом перемещении генов, расположенных рядом.
Фаги переносят наследственную информацию.
Формирование частиц фага лямбда
Вирус принадлежит к умеренным фагам E.coli, обладающим двухцепочной ДНК. Фаг внедряется в генную структуру бактерий в виде профага и передается нескольким поколениям.
Вирион лямбда состоит из следующих элементов: головки, хвоста, боковых нитей, белковой пленки. Внедрение сопровождается генетическим перераспределением материала на участке бактерии. В результате обмена наследственной информацией в присутствии вирусного и бактериального белка формируется форма Холлидея.
Вирус лямбда имеет 2 пути развития — мутагенный и литический. Фаг образует новые частицы, которые попадают во внешнюю среду, а клетка хозяина гибнет. В случае развития лизогенной реакции геном вириона проникает в хромосому E.coli и передается нескольким поколениям.
Вирус лямбда используют как вектор для клонирования, применяют для лечения холеры, дизентерии.
Какие события происходят в клетке реципиента
Вирус Л перемещает хромосомы от клеток донора к реципиенту. Ученые вывели геном фага, внедренного в хромосомную ДНК бактерии в культуре клеток кишечной бактерии К 12. В процессе получили расщепленные вирусом структурные элементы. Ими заражали культуры типа К 12. Затем их высевали в питательные среды и учитывали количество колоний, проверяя наличие аллелей от клеток-доноров, передавшихся мутантным реципиентам.
![]()
Ученые вывели геном фага, внедренного в хромосомную ДНК.
Вирион может переносить часть хромосомы, но процесс специфический, его осуществляет геном gal, находящийся на хромосоме бактериальной структуры. Многие трансдуктанты Cal+ нестабильны, теряют этот признак.
Абортивная трансдукция
При этом виде перемещения часть бактериального наследственного фактора не обменивается. Геном располагается в клетке, но не размножается. В случае деления клетки эта часть переходит в дочерние образования. После нескольких делений все клетки потомства остаются без этого фрагмента.
Процесс абортивного перемещения установлены Дж. Ледербергом в 1953 г. Он доказал, что часть хромосомы, оказавшейся в другой бактериальной зиготе, находится в клетке реципиента в виде непродуктивной частицы и передается 1 дочерней единице, а затем исчезает в потомстве.
Трансдукции клеток млекопитающих с вирусными векторами
Перенос фаговых векторов применяют для модификации наследственных факторов в клетках млекопитающих. В небольших молекулах ДНК гены локализуют в установленных местах вирусные частицы.
Плазмиды внедряют методом фланирования в структуру, образующуюся вместе с другими частями ДНК-конструкции, которые снабжены вирусными частицами, создающими инфекционные полноценные комплексы из нуклеиновой кислоты и капсида.
Для обеспечения безопасности процесса каждая плазмида содержит все данные, необходимые для формирования фага. Одновременное воспроизведение вируса в 2-3 плазмидах приводит к образованию инфекционного вириона.
Векторы, существующие в семействе палочковидных вирусов, могут синтезировать эукариотические протеины в клетках насекомых. Бакуловирус передает наследственную информацию зиготе млекопитающих, он не аутопродуктивен, а векторы, созданные на его основе, безопасны.
Векторы вирусов млекопитающих поражают гены в клетках-мишенях и используются как естественные градиенты для перемещения ДНК в эукариотические клетки. Учитывают такие факторы, как простота использования, регуляция во времени, скорость образования протеинов эукариотов.
Применяют векторы, созданные на основе аденовируса, ретровируса, возбудителя герпеса, лентовирусов.
Система имеет недостатки:
- неадекватность модификаций;
- большая чувствительность бакуловирусов.
Векторы, созданные на их основе, используют в биологии, биомедицине, для вакцинации и в генной терапии.
Трансдукция. Неспецифическая трансдукция. Специфическая трансдукция. Абортивная трансдукция. Феномен лизогении.
Трансдукция. Неспецифическая трансдукция. Специфическая трансдукция. Абортивная трансдукция. Феномен лизогении.
Трансдукция — перенос бактериофагом в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг. Трансдуцирующий бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент).
Выделено три типа трансдукции: неспецифическая (общая), специфическая и абортивная. В клетке, инфицированной бактериофагом, в ходе сборки дочерней популяции в головки некоторых фагов вместе с вирусной ДНК могут проникнуть фрагменты бактериальной ДНК или плазмиды. Вирусы ограничены в объёме генетического материала в соответствии с объёмом головки. Если ДНК бактериальной клетки расщепляется фагом в нетипичном месте, то чтобы освободить пространство для фрагмента хромосомной ДНК, некоторые участки вирусных ДНК «приносятся в жертву», что приводит к утере определённых их функций. При этом фаговая частица может стать дефектной. Количество аномальных фагов может достигать 0,3% всей дочерней популяции.
Образовавшийся фаг и есть частица, вызывающая неспецифическую (общую) трансдукцию. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены.
При неспецифической трансдукции фагом может быть перенесён любой фрагмент ДНК хозяина, а при специфической лишь строго определённые фрагменты ДНК. Наиболее известным примером специфической трансдукции служит трансдукция, осуществляемая фагом. Поскольку этот фаг при переходе в состояние профага включается в хромосому бактерий между генами, кодирующими синтез галактозы и биотина, именно эти гены он может переносить при транедукции. При абортивной трансдукции внесённый фрагмент ДНК донора не встраивается в генофор реципиента, а остаётся в цитоплазме, где его ДНК транскрибируется, но не реплицируется. Это приводит к тому, что при клеточном делении он передаётся только одной из дочерних клеток (то есть наследуется однолинейно) и затем теряется в потомстве.
Свойства трансдуцирующих фаговых частиц следующие:
• Частицы несут лишь часть ДНК фага, то есть не являются функциональными вирусами, а скорее ёмкостями, переносящими фрагменты бактериальной ДНК.
• Подобно прочим дефектным вирусам, частицы не способны к репликации.
• Трансдуцирующие фаги могут содержать какую-либо часть хромосомы хозяина с генами, дающими реципиентной бактерии некоторые преимущества (например, гены устойчивости к антибиотикам или гены, кодирующие способность к синтезу различных веществ). Подобное приобретение бактериями новых свойств получило название феномен лизогении.
• Феномен трансдукции может быть использован для картирования бактериальной хромосомы, если следовать тем же принципам, что и при картировании с использованием феномена трансформации.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Трансдукция
Лизогения бактерий и умеренные фаги. Наряду с конъюгацией и трансформацией у бактерий существует еще один механизм гибридизации — трансдукция. В ней непосредственное участие принимает бактериофаг.
Французские ученые А. Львов и А. Гутман (1950) описали явление лизогении и доказали существование неактивных форм фагов, названных профагами, в некоторых бактериальных клетках. Штаммы таких бактерий, называемых лизогенными, обладают иммунитетом против фагов, переносчиками которых они являются. В случаях, когда профаг переходит в активное или литическое состояние, ее ДНК быстро размножается, образовавшиеся вирусные частицы заполняют клетку и лизируют ее, как в случае обычного заражения вирусными фагами. Индуцировать литическую фазу бактериофага можно под воздействием УФ-облучения или химических реагентов.
Фаги, способные существовать в лизогенных бактериях, называются умеренными, в отличие от вирулентных фагов (например, Т2 или Т4), которые активно размножаются и приводят к гибели клетки-хозяина. В случаях заражения чувствительной (нелизогенной) бактериальной клетки фаг может активно размножаться и привести к лизису клетки либо становится профагом и превращает клетку-хозяина в лизогенную.
При размножении лизогенной бактерии все клетки клона также будут лизогенными, поскольку ДНК профага реплицируется вместе с бактериальной хромосомой. Таким образом, профаг переходит от поколения к поколениям новых бактериальных клеток подобно плазмидам (рис. 7.4) [1] .
Когда Э. Ледерберг обнаружила (1951), что ряд штаммов кишечной палочки Е. coli лизогенны к фагу X, это дало возможность активно изучать структуру и функционирование фага X и он стал самым изученным и полезным для исследований вирусом. Дальнейшие исследования позволили установить локализацию фага X на бактериальной хромосоме — это оказался особый участок между генами метаболизма галактозы (gal) и генами синтеза витамина биотин (bio). Данный участок молекулы ДНК бактериальной клетки имеет ту же последовательность нуклеотидов, что и участок фаговой ДНК, благодаря чему ДНК фага X легко встраивается в бактериальную хромосому. После рекомбинации (кроссинговера) ДНК фага X становится частью бактериальной ДНК Е. coli. Профаг лямбды имеет механизм контроля, благодаря чему, гены фага находятся в неактивном состоянии. Этот же механизм переводит все попадающие в бактериальную клетку вирусы в неактивную форму, поэтому лизогенные бактерии иммунны к большинству фагов того же типа.
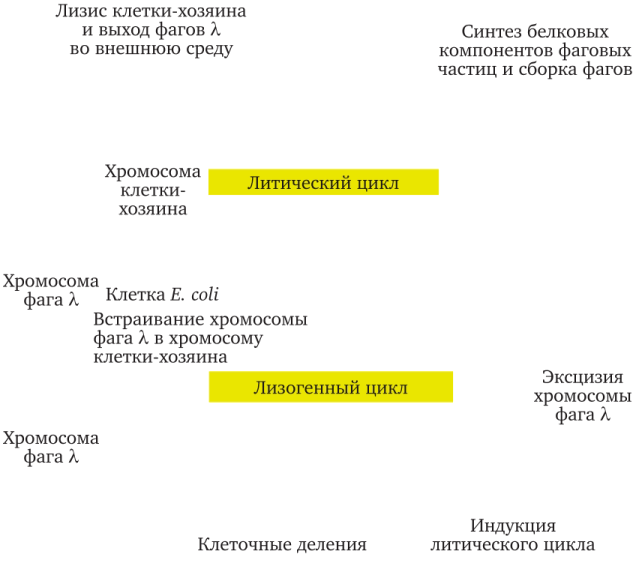
Рис. 7.4. Жизненный цикл умеренного фага на примере фага лямбда (X)
Профаги некоторых других умеренных фагов (например, Р1) не интегрируются в бактериальную хромосому, а находятся в клетке отдельно в виде плазмид. Явление лизогеннии характерно и для вирусов высших животных, при этом фазы активности вируса могу соответствовать стадиям протекания некоторых инфекционных заболеваний.
Явление трансдукции и ее типы. Трансдукция — гибридизация бактерий, совершающаяся в результате переноса умеренным бактериофагом генетических детерминантов из одной клетки, лизировавшейся при образовании фага, в другую. При трансдукции фаг переносит либо фрагмент бактериальной хромосомы, либо эписому.
Трансдукция впервые обнаружена при поисках половой гибридизации у S. typhimurium путем совместного культивирования двух ауксотрофных штаммов с последующим посевом их на минимальную среду для отбора гибридов. В опыте использовались штаммы LA-2, маркированный мутацией his (потребность в гистидине), и LA-22, несший ряд мутантных генов и обладавший, в частности, потребностью в триптофане (trp). На минимальной среде выявлялись рекомбинанты генотипа His, Trp. Дальнейшее изучение обнаружило ряд особенностей трансдукции.
Для получения рекомбинантов у S. typhimurium не требовалось контакта клеток. Они образовывались при помещении родительских штаммов в U-образную трубку, разделенную бактериальным фильтром. Однако рекомбинанты выявлялись только у штамма LA-22 с частотой 1 на 2-10 6 клеток. Таким образом, происходил односторонний перенос генетических факторов, и штамм LA-22 мог быть назван реципиентом, a LA-2 — донором (рис. 7.5) [2] .
Различают специфическую и общую формы трансдукции. Общая форма трансдукции бывает устойчивой и абортивной.
Общая (неспецифическая) трансдукция характерна для фага Р1, который существует в бактериальной клетке в виде плазмиды, а также для фагов Р22 и Ми, которые могут встраиваться в любой участок бактериальной хромосомы. При общей трансдукции после инфицирования при упаковке вирусных ДНК в капсид возможно ошибочное попадание в капсид фрагмента бактериальной хромосомы (приблизительно, с частотой 10~ 5 ). Фермент, контролируемый фагом, разрезает молекулу ДНК в различных участках, при этом получаются участки с длиной, равной нормальной фаговой ДНК. Образовавшиеся участки могут иметь различное происхождение: плазмиды, другие умеренные фаги, любой участок бактериальной хромосомы — и при общей трансдукции они могут включаться в созревающую вирусную частицу вместе с ДНК фага или вместо нее (рис. 7.6) [3] .
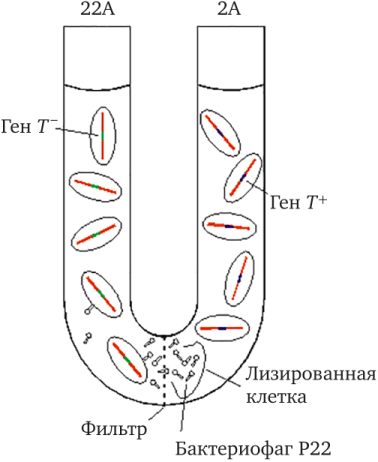
Рис. 7.5. Схема опыта, демонстрирующего явление трансдукции у Salmonella Typhimurium
При дальнейшем инфицировании бактериальных клеток фрагменты ДНК из вирусных частиц могут включаться в бактериальный геном клетки-хозяина путем обычной гомологичной рекомбинации.
Явление абортивной трансдукции заключается в сохранении фрагментов молекулы ДНК в клетке, не включающейся в геном, но сохраняющей за собой способность к транскрипции.
Наиболее изученной является специфическая трансдукция бактериофага X, для которого характерно встраивание только в определенный участок хромосомы Е. coli (att-сайт) с определенной последовательностью нуклеотидов, комплементарной att-сайту фага X.

Рис. 7.6. Схема общей трансдукции между штаммами Е. coli, осуществляемая фагом Р1
Во время индукции его исключение может пройти с ошибкой (вероятность 10" 3 —10" 5 на клетку): вырезается фрагмент тех же размеров, что и ДНК фага, но с началом не в том месте. При этом часть генов фага теряется, а часть генов Е. coli захватывается им. Вероятность переноса гена в этом случае падает при увеличении расстояния от него до att-сайта.
Для каждого специфически встраивающегося в хромосому умеренного фага характерны свой att-сайт и, соответственно, расположенные рядом с ним гены, которые он способен передавать. Ряд фагов может встраиваться в любое место на хромосоме и переносить любые гены по механизму специфической трансдукции. Кроме того, в хромосоме обычно есть последовательности, частично гомологичные аГС-участку ДНК фага. При повреждении полностью гомологичного аЦ-сайта можно добиться включения фага в хромосому по этим последовательностям и передачи в ходе специфической трансдукции генов, соседних уже с ними.
Когда умеренный фаг, несущий бактериальные гены, встраивается в хромосому новой бактерии-хозяина, она содержит уже два одинаковых гена — собственный и принесенный извне. Поскольку фаг лишен части собственных генов, часто он не может индуцироваться и размножиться. Однако при заражении этой же клетки «вспомогательным» фагом того же вида индуцирование дефектного фага становится возможным. Из хромосомы выходят и реплицируются как ДНК нормального «вспомогательного» фага, так и ДНК дефектного вместе с переносимыми им бактериальными генами. Поэтому около 50 % образующихся фаговых частиц несут бактериальную ДНК. Это явление носит название трансдукции с высокой частотой.
Если инфекция трансдуцирующим фагом X производится так, что на одну бактерию приходится в среднем меньше одной частицы (это предотвращает повторное заражение тран-сдуцированной клетки), то клетки оказываются лизогенными по фагу с необычными свойствами.
В результате трансдукции появляются мерозиготы, в которых ко всему генотипу реципиента добавляется лишь фрагмент хромосомы донора. Судьба этого фрагмента иожет быть различной.
Во-первых, если фрагмент встраивается в хромосому реципиента сразу, то образуются стабильные клоны рекомбинантного генотипа. Так дело обычно обстоит при общей устойчивой трансдукции. Аналогичные же стабильные рекомбинантные клоны составляют примерно 30 % трансдуктантов, возникающих при трансдукции фагом X.
Во-вторых, трансдуцированный фрагмент остается соединенным с генетическим материалом фага, а бактерия становится лизогенной по данному профагу. Такое состояние можно рассматривать в отношении трансдуцированного гена как мерозиготное. Оно сохраняется при клеточных делениях неопределенно долго. Клоны же обнаруживают нестабильность. При размножении клеток происходит выщепление тех из них, которые имеют генотип реципиента (за счет потери добавленного фрагмента), или стабильных рекомбинантных, образовавшихся при включении трансдуцированных генов в бактериальную хромосому. Ярким примером описанного типа является трансдукция, характерная для фага A.
В-третьих, перенесенный с помощью трансдукции фрагмент ДНК не встраивается в хромосому клетки-реципиента и не сохраняет связи с фагом. В данном случае он не репродуцируется и при делении клеток сохраняется в одной линии потомков. Это свойственно абортивной трансдукции.
Использование трансдукции для рекомбинационного анализа у бактерий. В ходе гибридологического изучения мутаций с помощью трансдукции можно обнаружить тесное сцепление генов, имеющих родственные биохимические функции. Например, анализ аллельности с использованием абортивной трансдукции ауксотрофных по триптофану мутаций у S. typhimurium позволяет распределить их в четыре группы, соответствущие генам trpA,trpB, trpC, trpD. Биохимический анализ показал, что указанные гены обладают самостоятельными функциями: trpA контролирует образование антраниловой кислоты, trpB — превращение этой кислоты в индолглицеро-фосфат, trpC — превращение последнего в индол, trpD — индола в триптофан. Рекомбинационный анализ при изучении устойчивой трансдукции показал, что эти гены сцеплены как друг с другом, так и с геном cys, и картируются в порядке: cys — trpA — trpB — trpC — trpD.
Метод трансдукции позволяет определить сцепленность близко расположенных генов, поскольку они могут оказаться в одном хромосомном фрагменте, переносимом фагом. Кроме кишечных бактерий, общая трансдукция описана у таких бактерий, как Р. aeruginosa, P. mirabilis, St. aureus, Act. olivaceus, Bac. subtilis.
Трансдукция и трансформация у бактерий
Трансдукция (от лат. transductio — перемещение) — перенос бактериофагом в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг. Трансдуцирующий бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент).
К трансдукции способны как умеренные фаги, так и вирулентные, последние, однако, уничтожают популяцию бактерий, поэтому трансдукция с их помощью не имеет большого значения ни в природе, ни при проведении исследований.
Содержание
Трансдукция у бактерий……………………………………….3
История изучения………………………………………………3
Поведение фагов в бактериальной клетке…………………… 3
Перенос фрагментов ДНК бактерии………………………….. 4
Общая (неспецифическая) трансдукция………………..4
Специфическая трансдукция…………………………… 5
Абортивная трансдукция………………………………. 7
Трансформация у бактерий……………………………………..9
2.1 История изучения………………………………………………..9
2.2 Трансформация у прокариот…………………………………….9
2.3 Стадии трансформации бактерий ………………………………11
Заключение……………………………………………………….12
Литература………………………………………………………..13
Прикрепленные файлы: 1 файл
реферат цитология мо.docx
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
ГОСУДАРСТВЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО
ИРКУТСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Трансдукция и трансформация у бактерий
- Трансдукция у бактерий……………………………………….3
- История изучения………………………………………………3
- Поведение фагов в бактериальной клетке…………………… 3
- Перенос фрагментов ДНК бактерии………………………….. 4
- Общая (неспецифическая) трансдукция………………..4
- Специфическая трансдукция…………………………… 5
- Абортивная трансдукция………………………………. 7
2.2 Трансформация у прокариот…………………………………….9
2.3 Стадии трансформации бактерий ………………………………11
Трансдукция у бактерий
Трансдукция (от лат. transduct io — перемещение) — перенос бактериофагом в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг. Трансдуцирующий бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент).
К трансдукции способны как умеренные фаги, так и вирулентные, последние, однако, уничтожают популяцию бактерий, поэтому трансдукция с их помощью не имеет большого значения ни в природе, ни при проведении исследований.
Эстер Ледерберг была первой учёной, кому удалось выделить бактериофаг лямбда, ДНК вирус, из Escherichia coli K-12 в 1950 году.
Собственно открытие трансдукции связано с именем американского учёного Д жошуа Ледерберга. В 1952 году он совместно с Нортоном Циндером обнаружил общую трансдукцию. В1953 Ледербергом и др. было показано существование абортивной трансдукции, в 1956 — специфической.
Поведение фагов в бактериальной клетке
Фаги способны к реализации двух путей развития в бактериальной клетке:
- Литический — после попадания в бактерию ДНК фага сразу же начинается его репликация, синтез белков и сборка готовых фаговых частиц, после чего происходит лизис клетки. Фаги, развивающиеся только по такому сценарию, называют вирулентными.
- Лизогенный — попавшая в бактериальную клетку ДНК фага встраивается в её хромосому или существует в ней как плазмида, реплицируясь при каждом делении клетки. Такое состояние бактериофага носит название профаг. Система его репликации в этом случае подавлена синтезируемыми им самим репрессорами. При снижении концентрации репрессора профаг индуцируется и переходит к литическому пути развития. Реализующие подобную стратегию бактериофаги называются умеренными. Для некоторых из них стадия профага является обязательной, другие в некоторых случаях способные сразу развиваться по литическому пути.
Перенос фрагментов ДНК бактериями
Перенос участков бактериальной хромосомы фагами был открыт в 1951 г. Ледербергом и Циндером у Salmonella typhimurium. В решающем эксперименте штамм-донор В + инфицировали умеренным бактериофагом Р22. После лизиса клетки-хозяина выделяли свободные фаги и инкубировали их вместе со штаммом-реципиентом В-, который генетически отличался от штамма В + по меньшей мере одним признаком. Авторы нашли, что после высева инкубированных клеток на подходящую среду появлялись рекомбинанты, обладавшие признаками штамма-донора В + .
Процессы, происходящие при таком неспецифическом переносе ДНК, весьма сложны. Во время репродукции фага Р22 в клетках штамма-донора В + в капсиды вместо фаговой ДНК могут включаться фрагменты бактериальной хромосомы. Таким образом, фаголизат содержит смесь нормальных и дефектных фагов. Заражение штамма-реципиента В" нормальным фагом ведет, как правило, к лизису клеток. Однако в некоторые клетки проникают дефектные трансдуцирующие фаги, ДНК которых способна рекомбинироваться с хромосомой реципиента. Происходит обмен гомологичными участками ДНК, что может привести к замене дефектного гена реципиента интактным геном донора.
Так как трансдуцируются лишь небольшие фрагменты ДНК, вероятность рекомбинации, затрагивающей какой-то определенный признак, очень мала: она составляет от 10 -б до 10- 8 . Становится понятно, что с помощью одной частицы фага Р22 Salmonella или неспецифически трансдуцирующего фага PI Escherichia coli в каждом случае может быть трансдуцирован только один ген (или несколько очень близко расположенных генов). Количество бактериальной ДНК, сравнимое с геномом фага, составляет лишь 1-2% всего количества ДНК, содержащегося в бактериальной клетке. Исключение составляет бактериофаг PBS 1 Bacillus subtilis, который может трансдуцировать до 8% генома хозяина.
Наиболее известным примером служит трансдукция, осуществляемая фагом X . Как уже говорилось, этот фаг при переходе в состояние профага включается в определенный участок хромосомы бактерии-хозяина. Отделение фаговой ДНК от бактериальной хромосомы (например, в результате УФ-облучения) может произойти неточно, т.е. какой-то фрагмент ее останется в хромосоме, а близко расположенные гены клетки-хозяина будут захвачены фаговой ДНК. По-видимому, причиной этого может быть неправильная рекомбинация.
В случае заражения трансдуцирующим фагом клеток, дефектных по определенному гену, например gal, может произойти рекомбинация с заменой собственного дефектного гена бактерии интактным трансду-цированным геном; при этом образуются рекомбинанты (трансдук-танты) gal + .
Подобным же образом происходит перенос генов бактериофагом Phi 80. Его ДНК включается в хромосому вблизи генов, кодирующих ферменты, ответственные за биосинтез триптофана. По этой причине Phi 80 особенно пригоден для переноса генов trp.
Предпосылкой успешного переноса генов при специфической транс-дукции (в отличие от неспецифической) является интеграция фага в геном клетки-хозяина.
В некоторых случаях было показано, что трансдуцированный фрагмент ДНК не вступает в рекомбинацию с хромосомой реципиента, а остается вне хромосомы. В этом случае клетка становится гетерозиготной по перенесенным генам. Перенесенная ДНК транскрибируется (на это указывает синтез соответствующего генного продукта), но не реплицируется. Это приводит к тому, что при клеточном делении донорский фрагмент переходит только в одну из дочерних клеток (абортивная трансдукция). Если реципиент ауксотрофный, а перенесенный фрагмент исправляет соответствующий дефект, то расти могут только те клетки, которые унаследовали этот фрагмент; при посеве на агар они образуют мельчайшие колонии.
При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
Свойства трансдуцирующих фаговых частиц следующие:
• Частицы несут лишь часть ДНК фага, то есть не являются функциональными вирусами, а скорее ёмкостями, переносящими фрагменты бактериальной ДНК.
• Подобно прочим дефектным вирусам, частицы не способны к репликации.
• Трансдуцирующие фаги могут содержать какую-либо часть хромосомы хозяина с генами, дающими реципиентной бактерии некоторые преимущества (например, гены устойчивости к антибиотикам или гены, кодирующие способность к синтезу различных веществ). Подобное приобретение бактериями новых свойств получило название феномен лизогении.
• Феномен трансдукции может быть использован для картирования бактериальной хромосомы, если следовать тем же принципам, что и при картировании с использованием феномена трансформации.
Читайте также: