
Базальные ганглии образуют прямые связи
Базальными ганглиями называют скопления нейронов в базальных отделах переднего и среднего мозга, которые участвуют в регуляции движения. К базальным ганглиям относят следующие структуры:
• Полосатое тело (хвостатое ядро, скорлупа, прилежащее ядро).
• Бледный шар (часть чечевицеобразного ядра), который имеет наружную (латеральную) и внутреннюю (медиальную) части. Внутренняя часть продолжается к среднему мозгу в виде сетчатой части черной субстанции.
• Субталамическое ядро (СТЯ).
• Пигментная компактная часть черной субстанции.
• Чечевицеобразное ядро, которое образуют скорлупа и бледный шар.
Вставочные нейроны оказывают прямой модулирующий эффект на обе группы проекционных нейронов за счет влияния на пресинаптические терминали и изменения выброса глутамата (корково-стриарный путь) и дофамина (нигростриарный путь). В полосатом теле выделяют отдельные ядра, которые получают нервные волокна от нейронов коры или от ядер таламуса. Полосатое тело также подразделяют на функциональные области: стриосому и матрикс. Нейроны прямого и непрямого путей расположены в матриксе. Нейроны, находящиеся внутри стриосомы, получают нервные импульсы от лимбической коры и отдают проекции к компактной части черной субстанции; эти нейроны образуют путь, через который базальные ганглии оказывают свое влияние на лимбическую систему.
Можно выделить, как минимум, четыре пути, которые формируются в коре, направляются к базальным ганглиям, а затем возвращаются обратно в кору:
1. Двигательная петля — отвечает за выученные двигательные акты.
2. Когнитивная петля — отвечает за намерения и планирование движения.
3. Лимбическая петля — отвечает за эмоциональную окраску движения.
4. Глазодвигательная петля — отвечает за произвольные саккады.
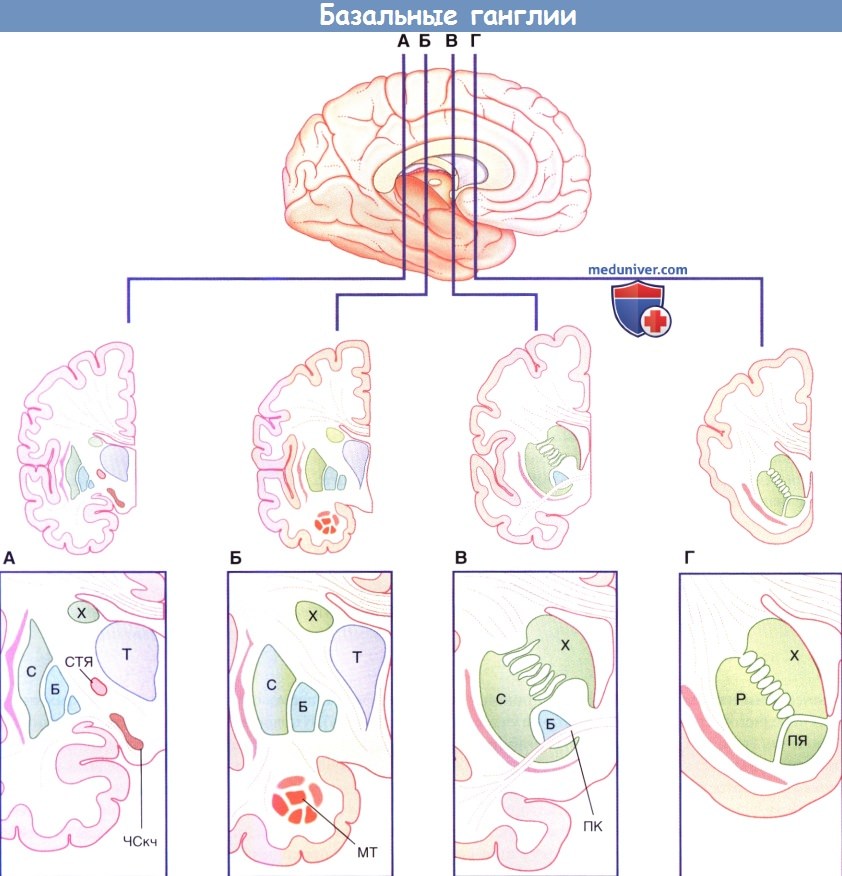
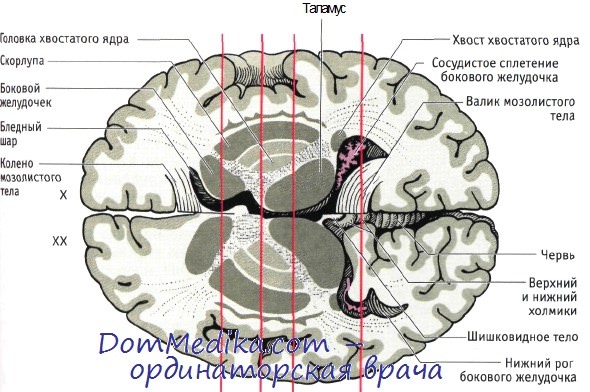
(А-Г). Четыре фронтальных среза головного мозга, вид сзади.
Вентральные отделы на рисунках снизу представлены в увеличенном виде.
ПЯ — прилежащее ядро; ПК — передняя комиссура; МТ — миндалевидное тело; X — хвостатое ядро; Б — бледный шар;
С — скорлупа; ЧСкч — компактная часть черной субстанции; СТЯ — субтапамическое ядро; Т — таламус.
а) Двигательная петля. Двигательная петля начинается в сенсомоторной коре, а возвращается обратно через полосатое тело, таламус и дополнительную моторную область (ДМО).
На рисунке ниже схематично изображено строение проводящих путей двигательной петли, в том числе задней части полосатого тела. Существует два пути. Прямой путь образован ядрами базальных ганглиев, таламуса и проходит через пять последовательных групп нейронов. Непрямой путь также включает СТЯ и состоит из семи групп нейронов. Отдельно от них проходят проекции от ВБШ к таламусу (чечевицеобразная петля и чечевицеобразный пучок), которые показаны на рисунке ниже.
Все проекции, начинающиеся от коры, исходят от пирамидных клеток и являются возбуждающими (глутаматергическими). Это же касается волокон, идущих от таламуса к ДМО. Волокна, начинающиеся от полосатого тела и обеих частей бледного шара, исходят от средних шипиковых нейронов и оказывают тормозящее влияние. Они являются ГАМК-ергическими, а также содержат нейропептиды с неустановленной в настоящее время функцией.
Нигростриарный путь идет от компактной части черной субстанции до полосатого тела, где на проекционных нейронах он образует два вида синапсов. Волокна, образующие синапсы с нейронами прямого пути,— стимулирующие, на дендритных шипиках расположены дофаминергические рецепторы типа 1 (D1). Волокна, образующие синапсы с нейронами непрямого пути,— тормозящие, их действие опосредовано до-фаминергическими рецепторами типа 2 (D2). Холинергические вставочные нейроны полосатого тела возбуждают проекционные нейроны; активность этих вставочных нейронов тормозит допамин.
У здорового человека черная субстанция находится в активном состоянии, постоянно стимулируя работу прямого пути. Стимуляция прямого пути необходима для активации ДМО перед движением и во время него. Возбуждение ДМО, которое возникает сразу перед началом двигательного акта, можно зарегистрировать при помощи электродов, прикрепленных к волосистой части головы. Эту электрическую активность называют (электрическим) потенциалом готовности (происхождение этого потенциала описано и показано на рисунке ниже). Импульсы от ДМО направляются к двигательной коре, откуда мозжечково-та-ламо-корковые волокна избирательно стимулируют пирамидные и корково-сетчатые нейроны за миллисекунды до генерации потенциала действия.
Скорлупа и бледный шар организованы соматотопически. За счет этого возможно селективно стимулировать нейроны, отвечающие, например, за движения рукой (прямой путь), и при этом одновременно тормозить ненужные движения, например ногой (непрямой путь). Для подавления ненужных в данный момент двигательных актов особенно важна работа СТЯ, которое может воздействовать на определенный сегмент ВБШ. Известно, что разрушение СТЯ приводит к тому, что движения конечностей противоположной половины тела становятся размашистыми и неконтролируемыми (см. далее).
Постепенное снижение выработки дофамина в компактном веществе черной субстанции — один из предрасполагающих факторов болезни Паркинсона (БП).
в) Лимбическая петля. На рисунке ниже изображена лимбическая петля базальных ганглиев. Она идет от нижней префронтальной коры через прилежащее ядро (передний конец полосатого тела) и вентральный отдел бледного шара, а затем возвращается в нижнюю префронтальную кору через дорсальное медиальное ядро (ДМЯ) таламуса. Прилежащее ядро и расположенный рядом обонятельный бугорок называют вентральным полосатым телом (вентральный стриатум).
Вероятнее всего, благодаря лимбической петле мы можем выражать свои эмоции в различных движениях, например улыбкой, жестами, принятием агрессивной или покорной позы. У этой петли имеется множество дофаминергических окончаний, которые разрушаются при БП. Из-за этого у таких больных лицо становится маскообразным, исчезает произвольная жестикуляция, через несколько лет развивается деменция.
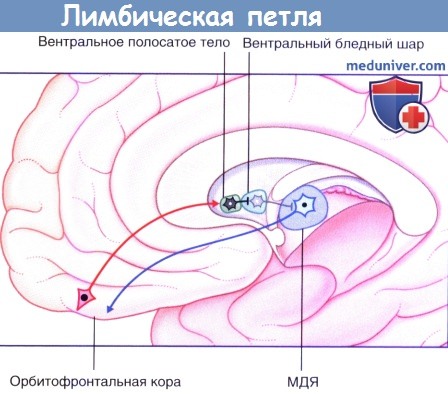
Лимбическая петля базальных ганглиев, правое полушарие.
Активация медиального дорсального ядра (МДЯ) таламуса происходит за счет его дезингибиции.
г) Глазодвигательная петля. Глазодвигательная петля начинается в лобном глазодвигательном поле (поле 8) и задней теменной коре (поле 7). Она проходит через хвостатое ядро и сетчатую часть черной субстанции. Далее через ПВЯ таламуса она возвращается в лобное глазодвигательное поле и префронтальную кору. От СЧЧС отходят тормозящие ГАМК-ергические волокна к верхним холмикам, где они образуют синапсы с нейронами, контролирующими непроизвольные саккады (Глава 23). Эти нейроны непосредственно иннервирует лобное глазодвигательное поле.
При фиксации взора СЧЧС находится в активном состоянии. Когда человек планирует перевести взгляд на какой-либо объект, происходят активация глазодвигательной петли и дезингибиция верхних холмиков (устранение торможения). При отсутствии нормального ингибирования со стороны базальных ганглиев происходит возбуждение верхних холмиков, в результате чего повышается активность прямого пути. Глазные яблоки с максимально высокой скоростью (22 м/с) перемещаются в сторону цели, после чего СЧЧС продолжает выполнять свою контролирующую функцию.
Для выявления глазодвигательной гипокинезии при болезни Паркинсона (БП) существует ряд специальных тестов. Движения глаз в сторону объекта, находящегося на периферическом поле зрения, обычно замедленны и иногда недостаточны. Объяснением этого симптома может служить нарушение дезингибиции верхних холмиков вследствие разрушения нейронов СЧЧС.
д) Резюме. Базальные ганглии представляют собой группу ядер, отвечающих за контроль над двигательной активностью. К ним относят полосатое тело (в том числе прилежащее ядро), бледный шар, СТЯ, черную субстанцию, а также вентральное латеральное (ВЛЯ), ПВЯ и ДМЯ таламуса. Бледный шар имеет наружную (НБШ) и внутреннюю (ВБШ) части, последняя продолжается в средний мозг в виде СЧЧС. Компактная часть черной субстанции лежит вне основных нейронных связей, но влияет на них за счет нигростриарного пути.
Кора больших полушарий возбуждает нейроны полосатого тела и СТЯ. Полосатое тело, в свою очередь, тормозит нейроны бледного шара. Влияние бледного шара на СТЯ и таламус также тормозящее. СТЯ возбуждает нейроны ВБШ.
Прямой путь, полосатое тело => ВБШ, находится в активированном состоянии за счет нормальной тонической активности нигростриарных дофаминергических нейронов. Непрямой путь, полосатое тело => НБШ => СТЯ => ВБШ, ингибирован. Для активации ДМО перед началом движения и во время него в двигательной петле происходит усиление передачи нервных импульсов по прямому пути. Активность ДМО, которую наблюдают непосредственно перед началом двигательного акта, можно зарегистрировать в виде потенциала готовности. Его появление обусловлено торможением нейронов ВБШ с последующим высвобождением (дезингибицией) таламо-корковых нейронов, посылающих нервные импульсы в ДМО. Затем сигнал поступает в двигательную кору, и начинается двигательный акт.
Полосатое тело и бледный шар организованы соматотопически, за счет чего становится возможным избирательный контроль той или иной части тела. Для подавления нежелательных движений особенно важно СТЯ.
Основной функцией двигательной петли, по всей видимости, служит организация правильного порядка выполнения отдельных движений. Это необходимо для реализации выученных двигательных программ. У пациентов с БП разрушение нигростриарных дофаминергических нейронов приводит к тому, что начинает преобладать активность непрямого пути. За этим следует подавление активности ВЛЯ таламуса и ДМО, что клинически проявляется брадикинезией. К основным симптомам БП относят ригидность, тремор и расстройство постуральных рефлексов.
Когнитивная петля начинается в ассоциативной коре, далее через ПВЯ таламуса она возвращается в премоторную и префронтальную кору. Активация когнитивной петли происходит во время изучения новых двигательных программ. Эта петля также отвечает за планирование двигательной активности.
Лимбическая петля начинается в поясной коре и миндалевидном теле, проходит через прилежащее ядро и направляется к ДМО. Вероятно, она отвечает за физическое выражение эмоций.
Глазодвигательная петля дезингибирует СЧЧС и таким образом активирует верхние холмики, которые инициируют саккады.
Гиперкинезы наблюдают при хорее Хантингтона, гемибаллизме и церебральном параличе (ЦП).
Редактор: Искандер Милевски. Дата публикации: 23.11.2018
Термин базальные ядра (или, как их еще называют базальные ганглии) означает, что эти ядра расположены у основания больших полушарий, латеральнее промежуточного мозга. Они составляют примерно 3% от объема полушарий. По общей структуре они подобны стволовым ядрам и разбиты на отдельные группы прослойками белого вещества. Из-за этого они имеют несколько пестрый внешний вид, что отразилось в объединяющем их названии – полосатое тело (corpus striatum).
Кроме полосатого тела в состав базальных ганглиев также входят ограда - тонкая прослойка серого вещества, расположенная латеральнее чечевицеобразного ядра и отделенная от него перегородкой из белого вещества (наружной капсулой) и миндалевидное тело (миндалина) – расположенная под чечевицеобразным ядром в самом переднем отделе височной доли. К базальным ядрам относят также субталамические ядра и черную субстанцию.
Бледный шар состоит из крупных нейронов, напоминающих мотонейроны и мелких – вставочных нейронов. Большие нейроны дают начало быстропроводящим волокнам, которые заканчиваются в красном ядре и черной субстанции среднего мозга, ядрах промежуточного мозга и образуют основной эфферентный выход полосатого тела.
Хвостатое ядро и ограда состоят из мелких клеток и их аксоны направлены, в основном, к бледному шару.
Система этих ядер:
- участвует в образовании экстрапирамидной системы и связана с регуляцией сложных движений и автоматических движений.
- задействована в переработке сенсорной информации, под ее контролем находятся центры гипоталамуса.
- является высшим регуляторным центром вегетативных функций терморегуляции и углеводного обмена.
Хвостатое ядро участвует в формировании условных рефлексов, механизмов памяти, эмоциях. Стриатум не получает прямых входов от спинного мозга и от сенсорных ядер мозгового ствола. Основные входы эта структура получает от:
- коры больших полушарий. Все корковые проекции на стриатум организованы ипсилатерально (на одноименную сторону тела), за исключением полей 4, 6 и 8, которые образуют билатеральные проекции (на обе стороны тела)
- от некоторых ядер таламуса
- от черной субстанции
- от миндалевидного тела.
Бледный шар и черная субстанция являются основными эфферентными структурами стриарной системы. Эти структуры отсылают волокна к таламусу, в область среднего мозга и каудально в область моста. Их синаптические окончания оказывают тормозное синаптическое действие (медиатор – g - аминомасляная кислота).
13. Лимбическая система.
(синонимы: лимбический комплекс, висцеральный мозг, ринэнцефалон, тимэнцефалон) представлена рядом подкорковых структур головного мозга, принадлежащих, в основном, к древней и старой коре. К этой системе относится:
· Области старой коры: поясная, или лимбическая извилина, гиппокамп
· Некоторые образования новой коры: височные и лобные отделы, промежуточная лобно-височная зона
· Подкорковые структуры: бледный шар, хвостатое ядро, скорлупа, миндалевидное тело, перегородка, гипоталамус, ретикулярная формация среднего мозга, неспецифические ядра таламуса.
Функции лимбической системы заключаются в следующем:
· Регуляция работы внутренних органов. При поражении лимбической системы отмечается нарушения деятельности сердечно-сосудистой системы, пищеварительной системы; при поражении миндалевидных ядер – нарушение обменных процессов в миокарде; поражение свода мозга вызывает нарушение кровоснабжения желудочно-кишечного тракта (до язвы)
· Гиппокамп – высший центр обоняния
· Обеспечивает различные формы поведения. Разрушение миндалевидных ядер вызывает нарушения инстинкта продолжения рода
· Отвечает за эмоциональные реакции.
· Обеспечивает различные формы памяти. Поражение гиппокампа вызывает ретроградную амнезию (потеря памяти на предшествующие события).
· При поражении поясной извилины страдает запоминание, выработка практических навыков.
· Способствует проявлению условных рефлексов.
Миндалевидный комплекс представляет собой довольно крупное ядерное образование (у человека - около 10 х 8 х 5 мм), расположенное в глубине передней части височной доли над ростральным отделом нижнего рога бокового желудочка. Миндалина образует связи с гипоталамусом, преимущественно с той его частью, которая участвует в контроле функции гипофиза. На мембране нейронов этой части миндалины есть рецепторы к половым и стероидным гормонам надпочечников. Благодаря этому циркулирующие в крови гормоны контролируют активность этих нейронов, а они, в свою очередь, могут влиять на гипоталамус и, таким образом, на секрецию из гипофиза (обратная связь), а также участвовать в формах поведения, контролируемых этими гормонами.
Миндалина образует также обширные связи с обонятельной луковицей. Благодаря этим связям обоняние у животных участвует в контроле репродуктивного (размножение) поведения. Например, феромоны (видоспецифические химические посредники) влияют на половое поведение через обонятельную систему. Многие виды животных имеют даже дополнительную обонятельную систему (так называемый якобсонов орган), передающую специализированную информацию к структурам лимбической системы, связанную с половым поведением. У человека эта система плохо развита, но полностью отрицать ее существование нельзя. В пользу этого может указывать хотя бы тот факт, что парфюмерия для женщин и мужчин различна.
У приматов, в том числе у человека, повреждения миндалины снижают эмоциональную окраску реакций, кроме того, у них полностью исчезает агрессивность в поведении.
Гиппокамп располагается в медиальной части височной доли. Особое место в системе связей гиппокампа занимает участок новой коры в районе гиппокампа (так называемая энторинальная кора). Этот участок коры получает многочисленные афференты практически от всех областей неокортекса и других отделов головного мозга (миндалины, передних ядер таламуса и др.) и является основным источником афферентов к гиппокампу. Гиппокамп получает также входы от зрительной, обонятельной и слуховой систем. Самой крупной проводящей системой гиппокампа является свод, который связывает гиппокамп с гипоталамусом. Кроме этого, гиппокамп обоих полушарий связаны между собой комиссурой (plasterium).
Повреждение гиппокампа приводит к характерным нарушениям памяти и способности к обучению. В 1887 г. русский психиатр С. С. Корсаков описал грубые расстройства памяти у больных алкоголизмом (синдром Корсакова). Посмертно у них были обнаружены дегенеративные повреждения гиппокампа. Нарушение памяти проявлялось в том, что больной помнил события отдаленного прошлого, в том числе детства, но не помнил о том, что произошло с ним несколько дней или даже минут тому назад. Например, он не мог запомнить своего лечащего врача: если врач выходил из палаты на 5 мин, больной его не узнавал при повторном посещении.
Для объяснения принципов интегративной деятельности лимбической системы выдвинуто представление о циклическом характере процессов возбуждения по замкнутой сети структур, включающих гиппокамп, сосцевидные тела, свод мозга, передние ядра таламуса, поясную извилину – так называемый круг Пейпеца (в 1937 году американский нейроанатом описал замкнутую цепочку структур, начинающуюся и заканчивающуюся в гипоталамусе). Считается, что круг Пейпеца представляет собой центр эмоций. Циркулируя по этому кругу, возбуждение создает длительные эмоциональные состояния, пробегая сквозь центры страха и агрессии, наслаждения и отвращения.
Лимбическая система в мозге человека выполняет очень важную функцию, которая называется мотивационно-эмоциональной. Лимбическая система тесно связана с ретикулярной формацией ствола мозга, как структурно, так и функционально. Вместе они образуют лимбико-ретикулярный комплекс. В лимбическую систему стекается весь поток информации от интеро- и экстерорецепторов, включая рецепторные поля органов чувств. Здесь происходит первичный синтез информации о состоянии внутренней среды организма и воздействующих на него факторов внешней среды. Здесь формируются элементарные потребности (например, в пище, воде, самообороне). Эти потребности представляют собой биологические мотивации (мотив – побуждение) для определенного типа поведения (например, поиск пищи), которое сопровождается определенной эмоциональной окраской. В зависимости от достижения результата эмоции могут быть как положительными, так и отрицательными.
Удовлетворение биологических потребностей направлено на поддержание гомеостаза и, следовательно, на выживание биологической системы. Контроль за состоянием внутренней среды осуществляют вегетативная и эндокринная системы, а лимбическая система – обеспечивает регуляцию вегетативно-висцерально-гуморальных отношений. От состояния лимбической системы зависит уровень сознания, активность двигательных и психических функций, состояния бодрствования и сна.
Базальные ганглии (базальные ядра) – это стриопаллидарная система, состоящая из трёх пар крупных ядер, погружённых в белое вещество конечного мозга в основании больших полушарий, и связывающих сенсорные и ассоциативные зоны коры с двигательной корой.
Филогенетически древняя часть базальных ганглиев – бледный шар, более позднее образование – полосатое тело и наиболее молодая часть – ограда.
Бледный шар состоит из наружного и внутреннего сегментов; полосатое тело – из хвостатого ядра и скорлупы. Ограда расположена между скорлупой и островковой (инсулярной) корой. В функциональном отношении базальные ганглии включают в себя также субталамические ядра и черную субстанцию.
Возбуждающая афферентная импульсация поступает преимущественно в полосатое тело (в хвостатое ядро) в основном из трёх источников:
1) от всех областей коры напрямую и опосредовано через таламус;
2) от неспецифических ядер таламуса;
3) от черной субстанции.
Среди эфферентных связей базальных ганглиев можно отметить три главных выхода:
- от полосатого тела тормозящие пути идут к бледному шару непосредственно и с участием субталамического ядра; от бледного шара начинается самый важный эфферентный путь базальных ядер, идущий преимущественно в двигательные вентральные ядра таламуса, от них возбуждающий путь идет в двигательную кору;
- часть эфферентных волокон из бледного шара и полосатого тела идет к центрам ствола мозга (ретикулярная формация, красное ядро и далее в спинной мозг), а также через нижнюю оливу в мозжечок;
- от полосатого тела тормозящие пути идут к черной субстанции и после переключения – к ядрам таламуса.
Следовательно, базальные ганглии являются промежуточным звеном. Они связывают ассоциативную и, частично, сенсорную кору с двигательной корой. Поэтому в структуре базальных ядер выделяют несколько параллельно действующих функциональных петель, связывающих их с корой больших полушарий.

Рис.1. Схема функциональных петель, проходящих через базальные ядра:
1 – скелетно-моторная петля; 2 – глазодвигательная петля; 3 – сложная петля; ДК – двигательная кора; ПМК – премоторная кора; ССК – соматосенсорная кора; ПФК – префронтальная ассоциативная кора; П8 – поле восьмой фронтальной коры; П7 – поле седьмой теменной коры; ФАК – фронтальная ассоциативная кора; ВЛЯ – вентролатеральное ядро; МДЯ – медиодорсальное ядро; ПВЯ – переднее вентральное ядро; БШ – бледный шар; ЧВ – черное вещество.
Скелетно-моторная петля соединяет премоторную, двигательную и соматосенсорную области коры со скорлупой. Импульсация от нее идет в бледный шар и черное вещество и далее через двигательное вентролатеральное ядро возвращается в премоторную область коры. Считают, что эта петля служит для регуляции таких параметров движения, как амплитуда, сила, направление.
Глазодвигательная петля соединяет области коры, контролирующие направление взгляда, с хвостатым ядром. Оттуда импульсация идет в бледный шар и черное вещество, из которых она проецируется соответственно в ассоциативное медиодорсальное и переднее релейное вентральное ядра таламуса, а из них возвращается в лобное глазодвигательное поле 8. Эта петля участвует в регуляции скачкообразных движений глаз (саккал).
Предполагается существование также сложных петель, по которым импульсация из лобных ассоциативных зон коры поступает в хвостатое ядро, бледный шар и черное вещество. Затем через медиодорсальное и вентральное переднее ядра таламуса возвращается в ассоциативную лобную кору. Считают, что эти петли участвуют в осуществлении высших психофизиологических функций мозга: контроле мотиваций, прогнозировании, когнитивной деятельности.
Функции полосатого тела
Влияние полосатого тела на бледный шар. Влияние осуществляется преимущественно тормозное медиатором ГАМК. Однако часть нейронов бледного шара дают смешанные ответы, а некоторые только ВПСП. То есть полосатое тело оказывает на бледный шар двоякое действие: тормозящее и возбуждающее, с преобладанием тормозящего.
Влияние полосатого тела на черное вещество. Между черным веществом и полосатым телом имеются двусторонние связи. Нейроны полосатого тела оказывают тормозящее влияние на нейроны черного вещества. В свою очередь, нейроны черного вещества оказывают модулирующее влияние на фоновую активность нейронов полосатого тела. Кроме влияния на полосатое тело черное вещество оказывает тормозящее действие на нейроны таламуса.
Влияние полосатого тела на таламус. Раздражение полосатого тела вызывает в таламусе появление высокоамплитудных ритмов, характерных для фазы медленного сна. Разрушение полосатого тела нарушает цикл сон-бодрствование уменьшением длительности сна.
Стимуляция полосатого тела. Стимуляция полосатого тела в различных его участках вызывает различные реакции: поворот головы и туловища в сторону, противоположную раздражению; задержку пищедобывательной деятельности; подавление ощущения боли.
Поражение полосатого тела. Поражение хвостатого ядра полосатого тела приводит к гиперкинезам (избыточным движениям) - хорее и атетозу.
Функции бледного шара
От полосатого тела бледный шар получает преимущественно тормозное и частично возбуждающее влияние. Но на двигательную кору, мозжечок, красное ядро и ретикулярную формацию он оказывает модулирующее влияние. На центр голода и насыщения бледный шар оказывает активирующее влияние. Разрушение бледного шара ведет к адинамии, сонливости, эмоциональной тупости.
Результаты деятельности всех базальных ядер:
- выработка вместе с мозжечком сложных двигательных актов;
- контроль параметров движения (сила, амплитуда, скорость и направление);
- регуляция цикла сон-бодрствоание;
- участие в механизме формирования условных рефлексов, сложных форм восприятия (например, осмысление текста);
- участие в акте торможения агрессивных реакций.
Базальные ганглии являются структурами ядерного типа. Они расположены внутри больших полушарий между лобными долями и промежуточным мозгом. Базальные ганглии относятся к собственно подкорковым образованиям мозга в самом узком смысле этого понятия и включают в себя три парных образования: неостриатум, паллидум (бледный шар) и ограда (claustrum).Неостриатум состоит из двух ядер: хвостатого и скорлупы (n. caudatus, putamen). Неостриатум является филогенетически новой структурой. Наиболее отчетливо он представлен начиная с рептилий. Скорлупа и хвостатое ядро по происхождению, нейронному строению, ходу проводящих путей и нейрохимическому составу являются сходными. Оба ядра, по существу, представляют собой два тяжа серого вещества, разделенных почти на всем протяжении волокнами внутренней капсулы. Паллидум, бледный шар (globus pallidum), в отличие от неостриатума, является филогенетически более древним образованием; его гомолог обнаруживается уже у рыб. Ограда расположена между скорлупой и островковой корой. Филогенетически ограда является самым новым образованием. У ежей и некоторых грызунов ее еще нет.
Морфофункциональные связи базальных ганглиев. Неостриатум образует связи с бледным шаром. Аксоны клеток неостриатума очень тонкие, до 1мкм, поэтому проведение возбуждения от неостриатума к паллидуму медленное. Стриапаллидарные волокна образуют в основном аксо-дендритные синапсы. Неостриатум оказывает двойственное влияние на нейроны паллидума – возбуждающее и тормозное. Неостриатум посылает прямые эфференты не только к паллидуму, но и к черной субстанции. Стрионигральные связи по своей природе моносинаптические и двусторонние. Большой интерес представляет обратная связь – от черной субстанции к неостриатуму. Считается, что аксоны нейронов черной субстанции, которые конвергируют к нейронам хвостатого ядра и к скорлупе, обеспечивают транспорт дофамина, синтезирующийся в нейронах черной субстанции. В неостриатуме он концентрируется в расширенных аксонных терминалях. Скорость транспорта дофамина по аксонам от черной субстанции к хвостатому ядру составляет приблизительно 0,8 мм в 1 час. Содержание дофамина в неостриатуме чрезвычайно велико. Имеются указания на то, что дофамина в неостриатуме млекопитающих в 6 раз больше, чем в паллидуме и передней части больших полушарий, в 19 раз больше, чем в мозжечке. Предполагается медиаторная роль этого амина в данной структуре. Кроме того, высказывается мнение о том, что дофамин активирует тормозные интернейроны неостриатума и таким образом подавляет деятельность его клеток. Выдвигается также предположение о том, что дофамин играет энергетическую роль в неостриатуме: через цАМФ он обеспечивает распад гликогена.
Помимо теоретического интереса в изучении медиаторной и метаболической функции дофамина, особое значение приобретает участие дофамина в патологии. Было установлено, что у больных с двигательными расстройствами резко падает концентрация дофамина в обоих ядрах неостриатума – хвостатом и скорлупе.
Кортикостриарные связи. Сейчас доказано, что почти от всех полей коры к хвостатому ядру и скорлупе конвергируют прямые волокна в составе внутренней капсулы и подмозолистого пучка. Наибольшее количество волокон идет к скорлупе и хвостатому ядру от передних отделов коры. Кортикостриарные волокна отличаются пространственной организацией. Топографически это проявляется в том, что передние области коры больших полушарий представлены в головке хвостатого ядра, а задние – в каудальном отделе хвостатого ядра (рис. 2.8).

Рис. 2.8. Базальные ганглии и структуры, связанные с ними
Скелетомоторная петля имеет входы от премоторной, моторной и соматосенсорной областей коры мозга. Основной поток информации идет через скорлупу, внутреннюю часть бледного шара или каудолатеральную область ретикулярной формации черной субстанции, затем через двигательные ядра таламуса и назад к шестому слою коры больших полушарий.
При регистрации активности индивидуальных клеток скорлупы и бледного шара у обезьян, которые были обучены стандартным движениям, обнаружены четкие корреляции между этими движениями и активностью определенных нейронов. Наблюдается четкая топографическая организация: активность нейронов строго определенной области базальных ганглиев всегда соответствует специфическим движениям конкретных частей тела. Кроме того, во многих случаях наблюдается корреляция с особыми параметрами движения: силой, амплитудой или направлением движения. Регистрация активности клеток показала, что путь от стриатума через латеральную область ретикулярной формации черной субстанции управляет главным образом движением лица и рта.
Окуломоторная (глазодвигательная) петля специализируется, вероятно, на регуляции движения глаз. Входные сигналы поступают от областей коры, контролирующих направление взгляда: фронтального глазного поля (поле 8) и каудальной части поля 7 теменной коры. Затем путь продолжается через хвостатое тело к дорсомедиальному сектору внутренней части бледного шара или к вентролатеральной области ретикулярной части черной субстанции. Затем идут связи к ядрам таламуса, которые дают проекции к фронтальному глазному полю. Аксоны нейронов сетчатой части черной субстанции раздваиваются, и одна ветвь идет к верхнему двухолмию среднего мозга, которое связано с движением глаз. Наблюдается положительная корреляция между активностью этих нейронов и саккадами (резкий перевод взгляда с одной точки на другую). Частота импульсации резко падает перед саккадой, что обусловлено тормозной стрианигральной связью (связью полосатого тела с черной субстанцией). Такое отключение тормозного выхода черной субстанции ведет к фазической активности таламуса или верхнего двухолмия. О полном пространственном разделении скелетомоторной и окуломоторной петель свидетельствует корреляция нейронной активности ретикулярной части черной субстанции с движениями либо глаз, либо рта, но никогда с теми и другими одновременно.
Медиаторная система базальных ганглиев. Прохождение информации в описанных выше множественных параллельных трансстриальных функциональных петлях может облегчаться или подавляться модулирующими системами. Описано несколько модулирующих систем. Особого внимания среди них заслуживает дофаминергическая система. Дофаминергические нигростриальные пути (черная субстанция – полосатое тело) начинаются в сетчатой части черной субстанции. Содержащие дофамин нейроны обнаружены также поодиночке или группами вне черной субстанции, но поблизости от нее.
Очень тонкие дофаминергические аксоны сильно ветвятся, образуя по всему стриатуму относительно диффузную сеть. Вдоль этих волокон находится множество небольших, заметных в световой микроскоп утолщений, называемых варикозами. На электронных микрофотографиях они идентифицируются как пресинаптические элементы. У нейронов сетчатой части черной субстанции довольно регулярная импульсация с частотой 1 Гц. Таким образом, каждую секунду импульс одной дофаминергической клетки вызывает высвобождение дофамина в многочисленных рассеянных по полосатому телу синапсах.
Аналогичное модулирующее влияние на базальные ганглии, возможно, оказывают серотонинергические волокна от ядер шва, норадренергические от голубого пятна,а также волокна с неизвестным медиатором от интраламинарных ядер таламуса и от миндалины; все они идут к полосатому телу. Кроме того, в базальных ганглиях содержится множество местных нейронов (интернейронов), модулирующих поток информации в трансстриатных петлях. К ним относятся холинергические нейроны полосатого тела и различные пептидергические нейроны.
В течение длительного времени полосатое тело рассматривали как крупную однородную массу клеток, и лишь недавно была обнаружена его модульная организация. Окончания двух обширных систем афферентных волокон от коры больших полушарий и от ламинарных ядер таламуса образует здесь небольшие четко ограниченные центры. Анатомические эксперименты с дифференциальным окрашиванием волокон, относящихся к разным системам, показали, что в хвостатом ядре перемешаны скопления нервных окончаний от лобной и височной ассоциативной коры. Гистохимические методы дают аналогичную картину: разные медиаторы (глутамат, ГАМК, ацетилхолин, различные пептиды) обнаруживаются в пределах мелких, четко очерченных участков. Сейчас эти центры считаются независимыми компартментами, или микромодулями. Удалось проследить топографическую организацию в виде продольных колонок, идущих через весь стриатум. Таким же образом организованы проекции лобной и височной ассоциативной коры. С помощью микроэлектродного тестирования выявлены соматотопические продольные колонки, относящиеся к скелетомоторной петле. Например, в колонке верхней конечности, вероятно, собираются сигналы от премоторной, моторной и соматосенсорной областей коры. Нейроны в такой колонке объединены по сходству их соматотопических свойств.
Читайте также: