
Центральная нервная система эксперимент
Одностороннее проведение возбуждения
Одним из основных свойств нервного волокна является проведение возбуждения в обе стороны. Между тем в целом организме возбуждение проходит только в одном строго определенном направлении; по одним нервам в центральную нервную систему, а по другим — из центральной нервной системы.
Эта способность центральной нервной системы проводить возбуждение только в одном направлении определяется свойствами синапсов, т. е. местом контакта нервных клеток. Следовательно, центральная нервная система проводит возбуждение только в одном направлении — от центростремительного нейрона к центробежному.
Эту особенность центральной нервной системы можно доказать, если производить запись токов действия в подходящих к спинному мозгу центростремительных и отходящих от него центробежных нервных волокнах. При раздражении центростремительного нервного волокна в центробежном нервном волокне, отходящем от спинного мозга, появляется ток действия. Если же раздражать центробежное волокно, то в центростремительном волокне ток действия не появится. Это объясняется тем, что спинной мозг про водит возбуждение только от центростремительного волокна к центробежному, но не обратно.
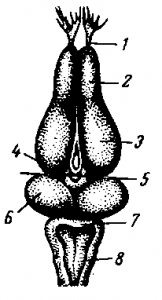
Рис. РАЗРЕЗ ГОЛОВНОГО МОЗГА ЛЯГУШКИ В ОПЫТЕ И. М. СЕЧЕHOBA. 1 — обонятельный нерв; 2 — обонятельная доля; 3 — большие полушария; 4 — зрительный бугор; 5 — линия разреза головного мозга для прикладывания кристаллика поваренной соли; 6 — двухолмие; 7 — мозжечок; 8 — продолговатый мозг.
Помимо нервных центров, односторонней проводимостью обладают также окончания двигательных нервов. Это можно наблюдать, если раздражать мышечные волокна. Возбуждение, которое при этом в них возникает, на нерв не передается.
Скорость проведения возбуждения
Любой рефлекс совершается в течение определенного времени: некоторые протекают быстрее, другие — медленнее. Время, которое протекает от начала раздражения рецепторов до начала ответа, называется временем рефлекса. Время рефлекса складывается из времени, которое необходимо для вызывания возбуждения в рецепторах и проведения возникшего импульса в центральную нервную систему, затем для про хождения возбуждения через центральную нервную систему и для распространения по центробежным нервам, далее для перехода на рабочий орган и, наконец, для скрытого периода возбуждения этого органа. Таким образом, время рефлекса, как мы видим, представляет сумму многих слагаемых.
Специальные исследования и измерения показали,что скорость проведения возбуждения неодинакова в разных отделах рефлекторной дуги. Медленнее всего возбуждение проходит через центральную нервную систему, где происходит передача возбуждения с одного неврона на другой. Поэтому принято говорить о центральной, или синаптической, задержке. Медленное проведение в центральной нервной системе называют задержкой потому, что получается впечатление, как будто возбуждение, дойдя до синапса, встречает какое-то препятствие и поэтому задерживается.
Превращение ритма возбуждения
Центральная нервная система никогда не посылает по центробежным волокнам на периферию к рабочему органу одну волну возбуждения; от нее всегда поступает ряд следующих друг за другом импульсов. Ритм возбуждения, который посылается центральной нервной системой, в значительной мере не зависит от частоты раздражнений, которые наносятся рецепторам. На любой ритм раздражения, даже на наиболее редкий, центральная нервная система все равно ответит рядом импульсов. Ритм импульсов, поступающих из нервных центров, колеблется от 50 до 200 в секунду.
Этой особенностью центральной нервной системы объясняется также то обстоятельство, что все мышечные сокращения в организме являются тетаническими. Самое быстрое и кратковременное сокращение любой мышцы является тетаническим в силу того, что мышца всегда получает ряд следующих друг за другом импульсов.
Суммация в озбуждения в центральной нервной системе
Следующей особенностью центральной нервной системы, впервые открытой И. М. Сеченовым, является ее способность суммировать поступающие возбуждения. Эта особенность заключается в
том, что если нанести аффе рентному волокну одиночное подпороговое раздражение, то центральная нервная система на такое раздражение не ответит и рефлекторной реакции не наступит. Если же нанести быстро несколько следующих друг за другом подпороговых раздражений, то центральная нервная система ответит возбуждением и наступит соответствующий рефлекторный акт.
Чем объяснить, что подпороговые импульсы, каждый из которых не был в состоянии вызвать возбуждение, поступая друг за другом в быстром темпе, вызывают возбуждение? Данное явление объясняется свойствами нервного центра, где каждое поступившее возбуждение вызывает ряд изменений, в частности повышает возбудимость нервного центра.
Последующие раздражения, если они довольно часты, попадают в период повышенной возбудимости и, суммируясь, становятся достаточными для возникновения волны возбуждения. Примером подобной суммации может служить чиханье, которое рефлекторно наступает только в результате длительного раздражения рецепторов, заложенных в слизистой оболочке носа, накопившейся слизью, частицами пыли или другими раздражающими веществами.
Утомляемость центральной нервной системы
От нервного волокна, которое почти не утомляемо, нервный центр отличается большой утомляемостью. Более или менее длительное раздражение центростремительного нерва приводит к постепенному уменьшению, а в дальнейшем — к полному
прекращению рефлекторного ответа. Н. Е. Введенский, раздражая центростремительный нерв, наблюдал ослабление и полное прекращение рефлекторного ответа через 10—40 секунд после начала раздражения. Перенося раздражение на соседний центростремительный нерв, он наблюдал появление рефлекса. Это наблюдение говорит о том, что утомление наступает именно в центральной нервной системе. Следующий эксперимент также дает возможность изучить явление утомления. Если, раздражая центростремительный нерв, добиться прекращения рефлекса, а затем перенести раздражение на центробежный нерв, то мышца ответит сокращением. Этот опыт свидетельствует о том, что утомление наступило именно в центральной нервной системе.
Изменение возбудимости центральной нервной системы
Дальнейшей особенностью центральной нервной системы является ее исключительно большая чувствительность к изменениям, наступающим в организме. В ответ на те или другие изменения меняется ее возбудимость. Малейшие изменения в газообмене и кровообращении сказываются на возбудимости нервных клеток.
Центральная нервная система потребляет кислорода больше всех других органов нашего тела: 100 г головного мозга собаки в одну минуту потребляют 10 мл кислорода, между тем такое же количество печени потребляют в 10 раз меньше, а такое же количество мышц — в 22 раза меньше кислорода. Уменьшение поступления кислорода очень быстро может привести к потере возбудимости, а затем к гибели нервных клеток.
Деятельность головного мозга зависит и от нормального кровообращения. Достаточно на короткий промежуток времени вызвать нарушение кровообращения мозга, как возбудимость его падает или даже полностью исчезает, и человек теряет сознание.
На возбудимость центральной нервной системы влияют некоторые яды, действующие преимущественно на мозг.
Исключительно сильнодействующим ядом является стрихнин. Стрихнин повышает возбудимость центральной нервной системы. Достаточно ввести животному небольшую дозу стрихнина, как оно начинает бурно реагировать даже на слабые раздражения. Если в лимфатический мешок лягушки ввести небольшое количество слабого раствора стрихнина, то стук по столу, на котором она лежит, вызывает у нее судороги. Такую же картину можно наблюдать и у теплокровных животных, у которых после введения небольшой дозы стрихнина появляются судороги на раздражения, на которые до введения стрихнина это животное не реагировало.
В небольших дозах стрихнин иногда применяется с лечебной целью.
На большие полушария головного мозга действуют яды, получившие название наркотиков. К ним относятся хлороформ, эфир, алкоголь и др. Первые два широко применяются в хирургической практике как вещества, вызывающие наркоз. Эти яды вначале вызывают повышение, а затем резкое падение возбудимости нервной системы и глубокий сон. Важным является то обстоятельство, что они действуют на большие полушария головного мозга и почти не влияют на продолговатый мозг, а это имеет очень большое значение для организма. В продолговатом мозгу находятся такие важные центры, как центр дыхания, центр сердечной деятельности и др., угнетение деятельности которых могло бы привести к гибели ор-
Особенно резко меняется возбудимость центральной нервной системы при ее повреждении. Если перерезать спинной мозг, то деятельность нервных центров, находящихся ниже поврежденного участка, угнетается. Такая потеря возбудимости нервных центров, находящихся ниже поврежденного участка, называется спинальным шоком. Через некоторое время шок проходит, и рефлекторная деятельность спинного мозга восстанавливается. Продолжительность шока у разных животных различна: чем выше стоит животное на зоологической лестнице, тем сильнее и продолжительнее шок. У лягушки шок проходит в течение нескольких минут, а у кошек и собак для этого требуются дни и недели. Наиболее тяжелым и длительным бывает шок у обезьян и у человека.
Торможение в центральной нервной системе
До середины прошлого века физиологам был известен только факт торможения работы органов под влиянием периферических нервов.
Впервые в 1862 г. И. М. Сеченов открыл явление центрального торможения. До него существовала точка зрения, что в центральной нервной системе имеет место только процесс возбуждения.
Опыты, в которых И. М. Сеченов открыл явление торможения в центральной нервной системе, были поставлены на лягушке. Для этой цели у лягушки вскрывали головной мозг и производили его поперечную перерезку по верхней границе зрительных бугров. Всю переднюю часть головного мозга удаляли. У приготовленной таким образом лягушки определяли время сгибательного рефлекса. После установления продолжительности времени рефлекса на зрительные бугры накладывали кристаллик поваренной соли. В результате нанесенного химического раздражения происходило торможение сгибательного рефлекса, и время рефлекса резко удлинялось. Схема опыта И. М. Сеченова видна на рис. Этим выдающимся открытием основоположник русской физиологии установил, что в центральной нервной системе одновременно с явлением возбуждения имеет место явление торможения.
Кроме того, было также доказано, что спинномозговые рефлексы находятся под влиянием высших отделов центральной нервной системы и под влиянием этих отделов рефлекторная деятельность спинного мозга может измениться.
Открытие И. М. Сеченова послужило началом целой серии опытов. Эти опыты показали, что торможение может наступить не только в результате непосредственного воздействия на нервные центры, как это было в опыте И. М. Сеченова, но и при одновременном раздражении двух или нескольких групп рецепторов. Если одновременно раздражаются две или несколько групп рецепторов, то в центральную нервную систему поступают возбуждения с разных участков тела, подвергающихся раздражению. Между возбуждениями, поступившими по разным нервам, происходит,борьба, причем более сильное угнетает слабое. В результате наступает торможение рефлекса, который должен был возникнуть на слабое возбуждение.
В итоге всех проведенных исследований было установлено, во-первых, что если к центру рефлекса с различных участков кожи или с разных отделов нервной системы одновременно поступают волны возбуждения, то рефлекс может быть заторможен; во-вторых, торможение является процессом, который, как и возбуждение, может возникнуть в центральной нервной системе при любом рефлекторном акте.
Рассмотрим несколько примеров торможения рефлексов. Если лапку спинальной лягушки опустить в раствор серной кислоты и одновременно пинцетом зажать вторую лапку, то лягушка намного позже выдернет лапку из кислоты, чем до зажатия второй лапки. Произойдет торможение сгибательного рефлекса. При сильных болях, чтобы не совершать защитных движений, часто стискивают зубы, прикусывают язык, чтобы не смеяться при щекотании, и т. п.
Торможение многих рефлексов может наступить под влиянием головного мозга. Так, например, можно задержать мочеиспускание, моргание и другие рефлексы, в которых принимают участие произвольные мышцы. Возбуждение и торможение— тесно связанные друг с другом процессы,
Открытое И. М. Сеченовым торможение в центральной нервной системе в дальнейшем дало возможность изучить такое сложное явление в организме, как координацию движений.
Статья на тему Основные свойства центральной нервной системы
В организме человека имеется 12 систем: центральная нервная система (ЦНС), дыхательная система, сердечно-сосудистая, кроветворная, пищеварительная, выделительная (включая мочевыделительную систему и кожу), репродуктивная система, эндокринная, костно-мышечная, лимфатическая, иммунная, периферическая нервная система. Не существует важных или неважных систем. Каждая нужна и каждая важна. Если в организме страдает одна из них, то через время, в процесс вовлекуться все остальные.
Центральная нервная система

Центральная нервная система — это система, которая обеспечивает в организме человека контроль над всей его жизнедеятельностью. В нашем организме находятся миллиарды, триллионы нервных клеток, которые существуют сами по себе и, к сожалению, не размножаются. За 3-4 года человек может полностью (в смысле здоровья клеток и их полноценной функции) восстановить печень, сердце, наша кровь обновляется через 4 месяца. Но с клетками мозга все по-другому. С течением жизни их не становиться больше, эти клетки возможно только полноценно питать и улучшать межклеточное пространство между ними, по возможности очищать от разного рода токсинов. Поэтому, если мы ребенка уморим с детства экологическими ядами, то это на всю оставшуюся жизнь. Например: приведем в садик, где красят стены, или в школу, которую не доремонтировали. Или отправим 3 раза в неделю плавать в бассейне с хлорированной водой. Никто не воспринимает нейроны как живую клетку. Ведь очень важно, чтобы мы понимали: нами руководит мозг. Пример: мы думаем, неплохо было бы купить творог. На самом деле — в организме дефицит кальция, а нейроны без кальция жить не могут, вот и посылают Вас в магазин за ним.
Для того чтобы понять, что необходимо нейрону, нужно изучить жизнь клетки. Для ее жизнедеятельности необходимы: 28 аминокислот, 15 минералов, 12 витаминов, жирные кислоты, ферменты, вода и кислород.
В медицине нервную систему разделили между двумя врачами: одной частью владеют невропатологи, другой -психиатры. Как будто поведение — это нечто особенное. Поведение — это жизнь нейрона. Чувство страха — это ничто иное как дефицит кислорода, который испытывает нейрон. Стресс, выделился адреналин, спазмировались сосуды, возник дефицит кровоснабжения мозга, отсюда нехватка кислорода. Формируется чувство страха.
Почему может заболеть головной мозг? Вспомним 12 причин возникновения заболеваний.
1. Психология и центральная нервная система
Сегодня биологической медициной доказано, что негативные эмоции, а больше всего — обида, оказывают самое разрушительное воздействие на тело человека и, в первую очередь, на центральную нервную систему. Человек с обидой в душе потенциальный онкобольной. Понимание этих механизмов дарит большую надежду человеку. Для меня это направление мыслей 10 лет назад открыли книги Луизы Хей и Лиз Бурбо. Многие серьезные результаты моих пациентов также были получены с применением этих знаний.
2. Питание и центральная нервная система
3. Вода и центральная нервная система
Ещё раз повторим: мозг на 90% состоит из воды. Мы не имеем права не пить 1,5 литра воды в день, потому что мы выделяем 1,0 литр мочи. А с дыханием и потом — и того больше.
4. Паразиты, вирусы и центральная нервная система
5. Медицина и центральная нервная система
Может ли она явиться причиной заболевания центральной нервной системы? 100% может. Практически все лекарственные препараты обладают нейротоксическим действием в той или иной степени: нтигистаминные, жаропонижающие, снотворные препараты, транквилизаторы.
6. Наследственность и центральная нервная система
Задумайтесь: цитомегаловирус, вирус герпеса, токсоплазма тоже передаются от матери к ребенку.
7. Травмы и центральная нервная система
Дорогие родители, оцените, каким видом спорта занимается ваш ребенок и насколько он травматичен. 2-3 удара по голове мячом могут привести к эпилепсии, а в лучшем случае к вегето-сосудистой дистонии.
8. Движение и центральная нервная система
Отсутствие движения напрямую не связано с нарушением работы центральной нервной системы, но это общий образ жизни, и косвенно он также является причиной заболеваний.
9. Биоэнергетика и центральная нервная система
Центральная нервная система первая отреагирует на любое энергетическое воздействие извне.
10. Экология и центральная нервная система
Может ли быть нормальным мозг маляра, работающего всю жизнь с маслами и красками, или шахтера? Даже теоретически не может. Что ему делать? Необходимо одно: очищать организм каждые три месяца, пить воду и правильно питаться. А кто позаботиться о нервной системе педагогов? У людей этой профессии она находиться в постоянном напряжении. Если бы удалось достучаться до врачей, которые занимаются профпатологией, то для каждой профессии можно было бы выбрать сберегающий фактор для центральной нервной системы.
11. Вредные привычки и центральная нервная система
Безусловно. Мозг реагирует на все, начиная от неправильного сна и чтения в общественном транспорте, до воздействия никотином и смолами при курении.
Поддержание центральной нервной системы
Вот что должен получать головной мозг для оптимальной работы.
Мальцева М.В. – Философия здоровья
Введение. Проблема физиологической и репаративной регенерации нервной системы всегда была в центре внимания нейробиологов и неврологов. В конце ХХ века большое количество исследований было посвящено исследованию трансплантации нервной ткани. (Полежаев и др., 1993; Семченко и др., 2000). Однако, несмотря на определенные достижения, полноценного приживления нервной ткани и восстановления функциональных связей при разного видах алло- и ксенотрансплантациях достигнуть не удалось.
Открытие нейрональных стволовых клеток (НСК), их обнаружение во взрослой нервной системе позвоночных животных и человека и развитие клеточных технологий позволило по новому взглянуть на эту проблему (Семченко и др. 2012; К.Н. Ярыгин, В.Н. Ярыгин, 2012; Ярыгин и др., 2015). В данной работе приводится краткий обзор собственных и имеющихся в литературе данных по регенерации нервной ткани в норме и в условиях эксперимента.
Нейрональные стволовые клетки (НСК) относятся к группе тканеспецифичных или региональных стволовых клеток. Они обладают характеристиками самоподдерживающейся популяции клеток, которые при дифференцировке способны давать нейроны, астроциты и олигодендроциты в развивающемся и взрослом мозге. Впервые они были обнаружены в ЦНС человека в 1995 году при анализе посмертных срезов мозга, окрашенных иммунногистохимически на BrdU (бромдезоксиуридин) (Gage et al., 1995). В настоящее время найден целый ряд нейрональных маркеров, которые позволяют более или менее надежно идентифицировать НСК и их потомки. Среди них следует отметить: ядерный антиген нервных клеток – NeuN; маркер нейробластов – даблкортин (DCX); нейрон-специфическую энолазу – NSE; молекулы адгезии нервных клеток – PSA-NCAM; цитоскелетные белки – нестин, β-тубулин III; транскрипционные факторы – Sox-1, Sox-2, Dlx2, Pax 6 и ряд других. Часть из этих маркеров специфична для клеток нервной ткани, другие направлены на выявление свойств, характерных для разных популяций стволовых клеток. (Гомазков, 2014; Коржевский и др., 2010, 2015).
Обнаружение НСК как в развивающемся, так и во взрослом мозге поставило вопрос об их происхождении. В ранний период эмбрионального развития НСК происходят из клеток нейроэпителия, которые путем симметричного и асимметричного деления дают начало нейронам, глиальным клеткам и клеткам-предшественникам, которые включаются в процессы нейрогенеза на более поздних этапах развития, включая постнатальный период. Характер деления клеток – один из механизмов выбора НСК путей развития. В случае симметричного митоза образуются две одинаковые дочерние клетки, которые либо сохраняют пролиферативный потенциал – т.е. остаются стволовыми, либо могут уйти на путь нейрогенеза или глиогенеза – опять же обе. При втором варианте симметричного деления популяция стволовых клеток может потерять способность к самообновлению и истощится. При ассиметричном делении одна клетка остается пролиферативной (стволовой), другая выходит в дифференцировку. При этом пул НСК сохраняется.
Таким образом, во взрослом мозге позвоночных животных и человека сохраняются группы клеток-предшественников, которые обладают свойствами НСК и способны в течение длительного периода обновлять популяции нейронов и глии.
Особо следует отметить, что сами НСК способны синтезировать и секретировать подобные вещества, действующие в данном случае по типу пара- или аутокринной регуляции.
Организация пролиферативных зон в мозге млекопитающих. Зоны взрослого нейрогенеза у млекопитающих, включая приматов, обнаружены в субвентрикулярной зоне (SVZ) латеральных мозговых желудочков конечного мозга и в субгранулярной зоне (SGZ) зубчатой фасции гиппокампа. Наличие подобных зон в других отделах ЦНС млекопитающих в настоящее время не доказано, а имеющиеся данные носят крайне противоречивый характер. (Ярыгин и др., 2014; Ярыгин, Ярыгин, 2012). Субвентрикулярная зона (SVZ) образована несколькими слоями клеток (от двух до пяти), в составе которых выделяют несколько типов клеток. Скорость увеличения числа новых клеток в зубчатой извилине гиппокампа (SGZ) взрослого мозга определяется как 9000 единиц в течение суток, что составляет примерно 6 % от ощего количества нейронов в зубчатой фасции гиппокампа крысы.или около 250 тысяч в месяц. (Cameron, McKay, 2001). Вновь образованные нервные клетки мигрируют на места своей локализации в данной структуре мозга, формируют систему отростков и синапсов и встраиваются в функциональные нейронные сети. Cледует отметить, что, хотя факт интеграции новых нейронов в существующие нейронные сети доказан, функциональные аспекты этого процесса во многом еще неясны.
В этом плане весьма интересным является обнаружение подобных пролиферативных зон в разных отделах головного мозга у представителей других групп позвоночных животных (рыб, амфибий, птиц). Взрослый нейрогенез у этих животных идет более интенсивно и дольше, чем у млекопитающих. (Обухов и др., 2015; Puschina et al., 2014; Grandel, Brand, 2013).
Взрослый нейрогенез и перспективы репаративной регенерации нервной ткани. Исследование последствий ишемии мозга показали, что она сопровождается усилением нейрогенеза в пролиферативных зонах и миграцией молодых клеток в зону повреждения. (Гомазков, 2014; Solway et al., 1998). Эти данные вызвали целую серию работ, направленных на изучение возможности использования НСК и их потомков для трансплантации в поврежденный мозг, а также поиска модельных объектов для экспериментальных работ (Григорьян, Кругляков, 2008; Семченко и др., 2012). Одной из удачных моделей явились рыбы разных видов. В серии работ с помощью иммуногистохимического маркирования PCNA (пролиферативного ядерного антигена), ядерного маркера нейрональной дифференцировки (HuCD), транскриптационного фактора Pax6 и серии других маркеров, в разных отделах мозга рыб был идентифицирован ряд пролиферативных зон (ПВЗ), свидетельствующих о наличии постоянного постнатального нейрогенеза в ЦНС рыб. (Обухов и др., 2015; Zupanc, 2009; Zupanc, Sîrbulescu, 2013; Puschina, Obukhov, 2012; Pushchina et al., 2014 a, b). Однако, в настоящее время неизвестно как этот процесс связан с нейрогенезом во взрослом мозге, и какие элементы матричных зон мозга рыб участвуют в репаративном нейрогенезе.
Были поставлены эксперименты на молоди нескольких видов рыб, которым наносили механическую травму в разные структуры мозга (зрительный нерв, крыша среднего мозга, полушария конечного мозга). При механическом повреждении разных отделов мозга молоди рыб (сетчатки, среднего мозга и мозжечка) было выявлено усиление пролиферативной активности как в традиционных пролиферативных зонах нейрогенеза (перивентрикулярные области), так зафиксировано появление новых нейрогенных участков. Процесс репарации после нанесения механической травмы глаза начинается с апоптоза поврежденных элементов. Апоптотический ответ наблюдается уже через полчаса после нанесения повреждающего воздействия и продолжается до 21 дня после нанесения травмы. Эти данные подтверждены результатами маркирования TUNEL-позитивных фрагментов ДНК в зоне повреждения (зрительного нерва), а также данными электронно-микроскопического анализа. Ультраструктурные изменения ядра свидетельствуют о различных стадиях процесса апоптоза в поврежденных клетках. Апоптоз, как механизм элиминации поврежденных в результате травмы клеток мозга рыб существенно отличается от такового у млекопитающих. У последних, основным способом элиминации поврежденных клеток в зоне травмы является некроз. Апоптоз же затрагивает незначительный объем клеток в прилегающих к травме областях. Наличие некроза в зоне травмы млекопитающих является одной из причин развития последующего вторичного воспаления в зоне повреждения, что в свою очередь вызывает дальнейшее нарастание некротического ответа в области травмы, в результате которого формируются большие полости, лишённые клеток. Эти полости, как правило, ограничены зоной реактивных астроцитов, создающих как механический, так и биохимический барьеры, затрудняющие рост нервных волокон и миграцию клеток в зону повреждения. В отличие от некроза при апоптозе отсутствуют признаки воспалительной реакции, а сами клетки впоследствии уничтожаются с помощью макрофагов/микроглии. Прижизненный мониторинг клеток в зоне повреждения с помощью мультифотонной конфокальной микроскопии показал, что уже через час после повреждающего воздействия наблюдается физиологический ответ со стороны макрофагов и микроглии, которые мигрируют в область нанесения механической травмы и активно участвуют в элиминации поврежденных клеток с помощью фагоцитоза. Дифференцировка клеток в нейрональном направлении, обнаруженная при помощи маркирования клеток антителами против белка HuC/D, происходила в пролиферативных зонах теленцефалона, зрительного тектума, мозжечка и продолговатого мозга форели уже через 2 дня после травмы (Пущина и др., 2016; Puschina et al., 2014).
Таким образом, показано, что после механической травмы в мозге экспериментальных животных источником новых нейронов являются появляющиеся в пролифератиных областях мозга новые зоны индуцированного нейрогенеза: нейрогенные ниши и участки вторичного нейрогенеза. (Пущина и др., 2016). Полученные данные послужат основой для дальнейших исследований особенностей постнатального нейрогенеза в ЦНС животных и человека в норме и при патологии.











- Физиология
![]()
- История физиологии
![]()
- Методы физиологии



Методы исследования центральной нервной системы
Частная физиология центральной нервной системы — раздел физиологии, изучающий функции структур головного и спинного мозга, а также механизмы их осуществления.
К методам исследования функций центральной нервной системы относятся нижеперечисленные.

Электроэнцефалография — метод регистрации биопотенциалов, генерируемых нейронами головного мозга, при отведении их от поверхности кожи головы. Величина таких биопотенциалов составляет 1-300 мкВ. Они отводятся с помощью электродов, накладываемых на поверхность кожи головы в стандартных точках, над всеми долями мозга и некоторыми их областями. Биопотенциалы подаются на вход прибора электроэнцефалографа, который их усиливает и регистрирует в виде электроэнцефалограммы (ЭЭГ) — графической кривой непрерывных изменений (волн) биопотенциалов мозга. Частота и амплитуда электроэнцефалографических волн отражают уровень активности нервных центров. С учетом величин амплитуды и частоты волн выделяют четыре основных ритма ЭЭГ (рис. 1).
Альфа-ритм имеет частоту 8-13 Гц и амплитуду 30- 70 мкВ. Это относительно регулярный, синхронизированный ритм, регистрируемый у человека, находящегося в состоянии бодрствования и покоя. Он выявляется приблизительно у 90% людей, находящихся в спокойной обстановке, при максимальном расслаблении мышц, с закрытыми глазами или в темноте. Альфа-ритм наиболее выражен в затылочных и теменных долях мозга.
Бета-ритм характеризуется нерегулярными волнами с частотой 14-35 Гц и амплитудой 15-20 мкВ. Этот ритм регистрируется у бодрствующего человека в лобных и теменных областях коры, при открытии глаз, действии звука, света, обращении к испытуемому, выполнении им физических действий. Он свидетельствует о переходе нервных процессов к более активному, деятельному состоянию и повышению функциональной активности мозга. Смену альфа-ритма или других электроэнцефалографических ритмов мозга на бета-ритм называют реакцией десинхронизации, или активации.

Рис. 1. Схема основных ритмов биопотенциалов головного мозга (ЭЭГ) человека: а — ритмы, регистрируемые с поверхности кожи головы в покос; 6 — действие света вызывает реакцию десинхронизации (смену α-ритма на β-ритм)
Тета-ритм имеет частоту 4-7 Гц и амплитуду до 150 мкВ. Он проявляется при поздних стадиях засыпания человека и развитии наркоза.
Дельта-ритм характеризуется частотой 0,5-3,5 Гц и большой (до 300 мкВ) амплитудой воли. Он регистрируется над всей поверхностью мозга во время глубокого сна или наркоза.
Основную роль в происхождении ЭЭГ отводят постсинаптическим потенциалам нейронов коры мозга. Считается, что на характер ЭЭГ-ритмов оказывает наибольшее влияние ритмическая активность пейсмекерных нейронов таламуса и ретикулярной формации ствола мозга. При этом таламус индуцирует в коре высокочастотные, а ретикулярная формация ствола мозга — низкочастотные ритмы (тета и дельта).
Метод ЭЭГ широко используется для регистрации нейронной активности в состояниях сна и бодрствования; для выявления очагов повышенной активности в мозге, например при эпилепсии; для исследования влияния лекарственных и наркотических веществ и решения других задач.
Метод вызванных потенциалов позволяет регистрировать изменение электрических потенциалов коры и других структур мозга, вызываемых стимуляцией различных рецепторных полей или проводящих путей, связанных с этими структурами мозга. Возникающие в ответ на одномоментное раздражение биопотенциалы коры носят волнообразный характер, длятся до 300 мс. Для выделения вызванных потенциалов из спонтанных электроэнцефалогических волн применяют сложную компьютерную обработку ЭЭГ. Эта методика используется в эксперименте и в клинике для определения функционального состояния рецепторной, проводниковой и центральной частей сенсорных систем.
Микроэлектродный метод позволяет с помощью тончайших электродов, вводимых в клетку или подводимых к нейронам, расположенным в определенной области мозга, регистрировать клеточную или внеклеточную электрическую активность нейронов, нервных центров, а также оказывать на них воздействие электрическими токами.
Стереотаксический метод позволяет вводить в заданные структуры мозга зонды, электроды с лечебной и диагностической целью. Их введение осуществляется с учетом трехмерных пространственных координат расположения интересующей структуры мозга, которые описаны в стереотаксических атласах. В атласах указывается под каким углом и на какую глубину относительно характерных анатомических точек черепа должны вводиться электрод или зонд для достижения интересующей структуры мозга. При этом голова больного фиксируется в специальном держателе.
Метод раздражения. Раздражение различных структур мозга чаще всего проводится с помощью слабого электрического тока. Такое раздражение легко дозируется, не вызывает повреждений нервных клеток и может наноситься многократно. В качестве раздражителей используются также различные биологически активные вещества.
Методы перерезок, экстирпации (удаления) и функциональной блокады нервных структур. Удаление структур мозга и их перерезки широко использовались в эксперименте в начальный период накопления знаний о мозге. В настоящее время сведения о физиологической роли различных структур ЦНС пополняются клиническими наблюдениями за изменением состояния функций мозга или других органов у больных, подвергшихся удалению или разрушению отдельных структур нервной системы (при опухолях, кровоизлияниях, травмах).
При функциональной блокаде производят временное выключение функций нервных структур путем введения веществ тормозного действия, воздействий специальных электрических токов, охлаждения.
Реоэнцефалография. Представляет собой методику исследования пульсовых изменений кровенаполнения мозговых сосудов. Она основана на измерении сопротивления нервной ткани электрическому току, которое зависит от степени их кровенаполнения.
Эхоэнцефалография. Позволяет определять локализацию и размеры уплотнений и полостей в мозге и костях черепной коробки. Эта методика основывается на регистрации ультразвуковых волн, отраженных от тканей головы.
Методы компьютерной томографии (визуализации). Основаны на регистрации сигналов от проникших в ткани мозга короткоживущих изотопов с помощью магниторезонансной, позитронно-эмиссионной томографии и регистрации поглощения проходящих через ткани рентгеновских лучей. Обеспечивают получение четкого послойного и трехмерного изображения структур мозга.
Методы исследования условных рефлексов и поведенческих реакций. Позволяют изучать интегративные функции высших отделов мозга. Эти методы подробнее рассмотрены в разделе интегративные функции мозга.
Электроэнцефалография (ЭЭГ) — регистрация электромагнитных волн, возникающих в коре головного мозга при быстром изменении потенциалов корковых полей.
Магнитоэнцефалография (МЭГ) — регистрация магнитных полей в коре головного мозга; преимущество МЭГ над ЭЭГ связано с тем, что МЭГ не испытывает искажений от тканей, покрывающих мозг, не требует индифферентного электрода и отражает только источники активности, параллельные черепу.
Позитивно-эмиссионная томография (ПЭТ) — метод, позволяющий с помощью соответствующих изотопов, введенных в кровь, оценить структуры мозга, а по скорости их перемещения — функциональную активность нервной ткани.
Магнитно-резонансная томография (МРТ) — основана на том, что различные вещества, обладающие парамагнитными свойствами, способны в магнитном ноле поляризоваться и резонировать с ним.
Термоэнцефалоскопия — измеряет локальный метаболизм и кровоток мозга по его теплопродукции (недостатком его является то, что он требует открытой поверхности мозга, применяется в нейрохирургии).
Читайте также: