
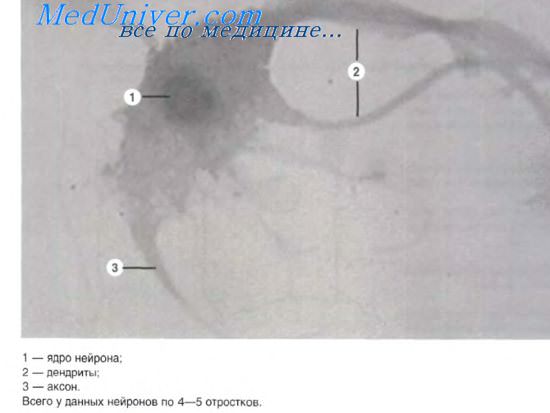
Нервное волокно окончания нервных волокон
Нервные волокна – это отростки нейронов, окруженные оболочками. Различают два вида нервных волокон:
Безмиелиновыенервные волокна имеют следующие компоненты:
− осевые цилиндры (отростки нейронов). Их в составе одного волокна несколько,
− шванновская оболочка, которая образована глиальными (шванновскими) клетками.
Под электронным микроскопом можно заметить, что осевые цилиндры располагаются в складках цитолеммы шванновских клеток, т.е. вне клеток. В связи с этим следует знать термин – мезаксон. Это сближенные края цитолеммы шванновских клеток при образовании складок. Место расположения безмиелиновых нервных волокон – это постганглионарные волокна вегетативной нервной системы. Скорость проведения нервных импульсов по безмиелиновым нервным волокнам – от 0,5 до 2 м/сек.
Миелиновые нервные волокна
Под световым микроскопом миелиновые нервные волокна представляют из себя однородные тяжи, которые на определенном расстоянии друг от друга имеют сужения (перехваты Ранвье) (Рис. 17).
Компоненты миелинового нервного волокна:
− миелиновая оболочка и
Осевой цилиндр (отросток нейрона) в составе миелинового нервного волокна – один. Как указывалось ранее, он имеет оболочку (аксолемма) и цитоплазм (аксоплазма).
Миелиновая оболочка. Чтобы понять строение миелиновой оболочки следует проследить развитие миелиновой оболочки. Вначале осевой цилиндр погружается в складку цитолеммы шванновских клеток, которые затем начинают закручиваться вокруг осевого цилиндра. При этом мезаксон (сдвоенная цитолемма шванновских клеток) ложится в виде слоев вокруг осевого цилиндра, образуя миелиновую оболочку. Таким образом, миелиновая оболочка – это наслоенные друг на друга листки мезаксона, т.е. наслоенные друг на друга липопротеидные мембраны.

- Осевой цилиндр.
- Миелиновый слой
б. Перехват Ранвье
в. Насечка миелина
Шванновская оболочка. Это наружная оболочка нервного волокна, которая содержит цитоплазму и ядра шванновских клеток.
Перехват Ранвье это место стыка (соединения) двух соседних шванновских клеток. Эти участки имеют особенности строения:
− в них отсутствует миелиновая оболочка; осевой цилиндр здесь покрыт только шванновской оболочкой,
− в этом месте отмечается скопление митохондрий.
Перехваты Ранвье – это функционально активные участки миелинового нервного волокна, а именно, передача нервных импульсов в нем совершается скачкообразно, от одного перехвата до другого.
Скорость проведения нервных импульсов в миелиновых нервных волокнах – от 3 до 100 м/сек.
Миелиновые нервные волокна – это основной вид волокон. Все нервные волокна в составе центральной и периферической нервной системы (за исключением постганглионарных волокон вегетативной нервной системы) по строению миелиновые.
Периферические нервы
Состоят из миелиновых (в основном) нервных волокон. Миелиновые нервные волокна, кроме миелиновой и шванновской, имеют оболочку – неврилемма, которая имеет в своем составе: базальную мембрану и сеть коллагеновых волокон. Между отдельными нервными волокнами прослойки рыхлой соединительной ткани – это эндоневрий. Соединительнотканные прослойки между пучками нервных волокон – периневрий и общая соединительнотканная оболочка нервного ствола – эпиневрий.
Нервные окончания
− чувствительные нервные окончания,
− двигательные нервные окончания и
Чувствительные нервные окончания (рецепторы). Определения:
1. Рецептор – это специализированный нервный аппарат, воспринимающий действия раздражителей,
2. (по И.П. Павлову), рецепторы – это трансформаторы энергии раздражителей в нервные импульсы.
Физиологическая классификация рецепторов:
− механорецепторы – стимулируются механическими движениями органов или их частей,
− барорецепторы – воспринимают изменения давления,
− хеморецепторы – стимулируются различными химическими веществами,
− терморецепторы – воспринимают колебания температуры.
Морфологическая классификация чувствительных нервных окончаний. По строению их делят на: свободные и несвободные. Несвободные в свою очередь делят на: неинкапсулированные и инкапсулированные.
Свободные нервные окончания – это голые ветвления осевого цилиндра, т.е. они (ветвления) свободны от оболочек (миелиновой и шванновской) (Рис. 18). Отличаются большой чувствительностью. Их много в составе эпителия, особенно в составе разновидностей многослойного эпителия. Пример. Роговица глаза отличается высокой чувствительностью, из-за наличия между клетками эпителия большого количества свободных чувствительных нервных окончаний.

- Слой шиповатых клеток
- Рыхлая соединительная ткань
- Терминали
- Нервные волокна
Несвободные нервные окончания. В них концевые ветвления осевого цилиндра покрыты оболочками.
Несвободные неинкапсулированныенервные окончания – когда концевые ветвления осевого цилиндра покрыты оболочкой из клеток нейроглии. Таких рецепторов много в составе эпителия и соединительных тканей.
Несвободные инкапсулированные нервные окончания. Их разновидности: колбы Краузе, тельца Мейснера, тельца Фатер – Пачини. По строению они сходны. Рассмотрим их строение на примере телец Фатер – Пачини (Рис. 19). В средне части тельца располагается концевое ветвление осевого цилиндра. Оно окружено видоизмененными глиальными клетками, образующими внутреннюю капсулу тельце. Внутренняя капсула окружена наружной капсулой, состоящей из наслоенных друг на друга соединительнотканных пластинок. Тельца Фатер – Пачини воспринимают давление на орган, т.е. функционально механорецепторы.

1.Секреторные отделы железы
2, 3. Пластинка соединительной ткани
б. Осевой цилиндр
4. Миелиновое нервное волокно.
Двигательные нервные окончания передают нервные импульсы на рабочие органы и ткани. В скелетных мышцах они называются нервно-мышечные окончания.
Синапс. Это место контакта двух нейронов. В зависимости от того, какие части двух нейронов контактируют, различают следующие виды синапсов:
− аксо-соматические, когда аксон первого нейрона контактирует с телом второго,
− аксо-дендритические, когда аксон первого нейрона контактирует с дендритов второго,
− аксо-аксональные, когда аксон первого нейрона контактирует с аксоном второго.
Синапсы имеют две части:
− пресинаптический, полюс и
Пресинаптический полюс – это концевое расширение аксона первого нейрона; содержит скопления митохондрий и пресинаптические пузырьки, имеющие в своем составе медиатор (ацетилхолин или норадреналин) и пресинаптическую мембрану (часть аксолеммы концевого расширения).
Постсинаптический полюс – это прилежащая часть второго нейрона. Имеет постсинаптическую мембрану.
Между пре- и постсинаптической мембраной располагается синаптическая щель. Основное свойство синапса – одностороннее проведение нервных импульсов от пресинаптического полюса в постсинаптический.
Нервные волокна – это отростки нервных клеток, покрытые снаружи глиальной оболочкой и осуществляющие проведение нервных импульсов. Отросток нервной клетки (аксон или дендрит), расположенный в центре нервного волокна, называют осевым цилиндром. Осевой цилиндр представляет собой вырост нейроплазмы тела нервной клетки с содержащимися в ней органеллами, покрытый оболочкой – аксолеммой.
В зависимости от наличия или отсутствия в составе глиальной оболочки миелина различают два вида нервных волокон – миелиновые и безмиелиновые. В миелиновых волокнах глиальная оболочка толще и составляет на поперечном разрезе 1/2–2/3 диаметра всего нервного волокна. Содержащийся в миелиновых волокнах миелин придает им белый цвет.
Миелиновые волокна по диаметру делят на три группы – толстые (12–20 мкм), средние (6–12 мкм) и тонкие (1–6 мкм). Через каждые 1–3 мм нервное волокно резко истончается, образуются узловые перехваты (перехваты Ранвье) шириной 1 мм. В области перехватов миелиновый слой отсутствует – эго место соединения соседних глиальных (шванновских) клеток. В зависимости от диаметра волокна различается скорость проведения нервного импульса. В толстых миелиновых волокнах она составляет примерно 80–120 м/с, в средних – 30–80 м/с, в тонких – 10–30 м/с. При этом скорость прохождения импульсов в определенной группе волокон нс зависит от силы раздражения.
Установлено, что толстые миелиновые волокна являются преимущественно двигательными, средние по диаметру волокна проводят импульсы тактильной и температурной чувствительности, а тонкие – болевой. Таким образом, по составу волокон можно дать функциональную характеристику нерва (двигательный, чувствительный, смешанный).
Миелиновая оболочка предотвращает распространение идущих по волокну нервных импульсов на соседние ткани, т.е. она выполняет роль диэлектрика (изолятора). Миелинизация нервных волокон начинается на 4–5-м месяце внутриутробного развития и имеет неодинаковую продолжительность в различных отделах нервной системы. Завершение процесса миелинизации нервных волокон свидетельствует о зрелости нервных структур. Так, нервные волокна полушарий большого мозга, ответственные за эмоционально-психические функции, миелинизируются только к 12–13 годам.
Безмиелиновые волокна имеют небольшой диаметр – 1–4 мкм и проводят нервные импульсы со скоростью 1–2 м/с. Причем, в отличие от миелиновых волокон, импульсы в них проводятся не скачкообразно, а непрерывно. Безмиелиновые нервные волокна являются эфферентными волокнами вегетативной нервной системы. Они обеспечивают иннервацию внутренних органов, желез и сосудов.
В зависимости от направления проведения нервного импульса по отношению к центральной нервной системе различают две группы волокон – центростремительные и центробежные. Центростремительные волокна направляются к спинному или головному мозгу и функционально являются афферентными (восходящими). Центробежные волокна идут от головного или спинного мозга к рабочим органам (мышца, сосуд, железа) и называются эффекторными.
Нервные волокна, расположенные в пределах центральной нервной системы, составляют белое вещество спинного и головного мозга.
Нервные окончания – это концевые отделы нервных волокон. В зависимости от выполняемой функции различают три вида окончаний: рецепторы, эффекторы и межнейронные контакты – синапсы.
Рецепторы – это нервные окончания периферических отростков чувствительных (рецепторных) нейронов, обеспечивающие восприятие специфических раздражений из внешней или внутренней среды и трансформацию энергии раздражения в нервный импульс.
По локализации рецепторы делят на три группы: экстероцепторы, проприоцепторы и интероцепторы (рис. 1.2).
- 1. Экстероцепторы располагаются в коже и слизистых оболочках полости рта, носа и органа зрения (в конъюнктиве). Они воспринимают тактильные, температурные и болевые раздражения из внешней среды.
- 2. Интероцепторы находятся во внутренних органах. Адекватными раздражителями для них являются преимущественно химические вещества и механические воздействия. Интероцепторы воспринимают химический состав определенных веществ, степень наполнения органов или болевые ощущения.
- 3. Проприоцепторы, или глубокие рецепторы, локализуются в мышцах, сухожилиях, фасциях, надкостнице, связках и суставных капсулах. Они воспринимают такие раздражения, как чувство веса, давления, вибрации, положение частей тела, степень напряжения мышц.
Экстеро-, интеро- и проприоцепторы являются преимущественно рецепторами общей чувствительности.
Рецепторы специализированных органов чувств (орган зрения, слуха, равновесия, обоняния и вкуса) составляют группу рецепторов специальных видов чувствительности. Они воспринимают зрительные (свет и цвет), слуховые (звук и шум), вестибулярные (угловые и вертикальные ускорения), обонятельные (запахи) и вкусовые раздражения.
Рецепторы, воспринимающие раздражения путем непосредственного контакта с раздражителем, называются контактными (например, болевые, температурные, вкусовые). Рецепторы, воспринимающие раздражения на значительном удалении от организма, являются дистантными (например, зрительные, слуховые).

Рис. 1.2. Основные типы рецепторов соматической нервной системы
По строению рецепторы делят на три группы – свободные нервные окончания, инкапсулированные нервные окончания и нервные окончания, представленные первично чувствующими клетками. Свободные нервные окончания воспринимают боль; инкапсулированные – тактильные, температурные и проприоцептивные раздражения; первично чувствующие клетки – зрительные, слуховые, вестибулярные и вкусовые раздражения.
Синапс – это специализированное морфофункциональное образование, предназначенное для передачи нервного импульса контактным способом с одного нейрона на другой или с нейрона на рабочий орган.
По локализации синапсы могут быть межнейронными и нейротканевыми. В первой группе в зависимости от контактирующих частей нейрона выделяют аксо-соматические (аксон – тело), аксо-дендритические (аксон – дендрит), аксо-аксональные (аксон – аксон) синапсы. На одном нейроне может находиться до 10 тыс. синаптических образований. Аксо-соматические и аксо-дендритические синапсы обеспечивают распространение импульса с одной нервной клетки на другую. Аксо-аксональные синапсы обеспечивают торможение импульсов.
Нейротканевые синапсы по расположению делят на нервно-мышечные и нервно-секреторные.
По механизму передачи нервного импульса различают три группы синаптических структур:
- 1) синапсы с химической (медиаторной или трансмиттерной) передачей импульса;
- 2) синапсы с электрической передачей нервного импульса (эфапсы);
- 3) синапсы со смешанной передачей нервного импульса.
Морфологически синапс представляет собой утолщение в виде пуговок, бляшек, колбочек или нитей. На ультраструктурном уровне в нем выделяют пресинаптическую часть, синаптическую щель и постсинаптическую часть (рис. 1.3). Пресинаптическая часть для синапсов с химической передачей обычно образована терминальным аппаратом аксона и содержит скопление синаптических пузырьков и митохондрий. Синаптические пузырьки наполнены медиатором. В качестве медиатора чаще выступают такие вещества, как ацетилхолин, норадреналин, гамма-аминомасляная кислота (ГАМК), гистамин, дофамин, серотонин и т.д. По величине синаптических пузырьков можно судить о виде медиатора: ацетилхолин находится в мелких пузырьках диаметром 30–50 нм; норадреналин – в пузырьках средней величины – 50–90 нм; ГАМК – в крупных пузырьках – 100–120 нм. В момент поступления нервного импульса в пресинаптическую часть медиатор освобождается из связанного состояния и выбрасывается в виде пузырьков в синаптическую щель. В одном пузырьке содержится до 10 тыс. молекул медиатора.
Синаптическая щель имеет ширину 10–20 нм и заполнена гелем (межклеточным веществом). Более широкая синаптическая щель характерна для синапсов с химической передачей и узкая (до 10 нм) – для эфапсов.

Рис. 1.3. Строение синапса:
1 – пресинаптическая мембрана; 2 – молекулы медиатора; 3 – синаптическая щель; 4 – постсинаптическая мембрана; 5 – обратный транспорт медиатора; 6 – синаптические пузырьки с медиатором
Пройдя синаптическую щель, медиатор связывается с хеморецептором (белковая структура) на постсинаптической мембране. В зависимости от химической природы медиатора различают следующие основные виды хеморецепторов; α-, β-адренорецепторы; Μ-, Н-холинорецепторы; пуринорецепторы, ГАМК-рецепторы и т.д. Альфа-, бета- адренорецепторы реагируют с такими медиаторами, как адреналин, норадреналин, дофамин, т.е. с катехоламинами; М-, Н-холинорецепторы – с ацетилхолином; пуринорецепторы – с пуриновыми основаниями и ГАМК-рецепторы – с гамма-аминомасляной кислотой.
Прореагировав с хеморецептором, медиатор разрушается (инактивируется) имеющимися в хеморецепторе веществами (ацетилхолин – ацетилхолинэстеразой, норадреналин – моноаминооксидазой и т.д.). Инактивированные молекулы медиатора обратно всасываются через пресинаптическую мембрану, где подвергаются восстановлению.
Таким образом, при химической передаче нервных импульсов последовательно проходит четыре этапа: синтез медиатора, проникновение медиатора через пресинаптическую мембрану, взаимодействие с хеморецепторами постсинаптической мембраны, инактивация.
Ультраструктурные особенности строения синапса определяют следующие закономерности его функционирования:
- 1) односторонность проведения нервного импульса (закон динамической поляризации синапса), обусловленная возможностями синтеза, проникновения и взаимодействия медиатора;
- 2) синаптическая задержка, связанная с затратой времени на диффузию медиатора и реакцию взаимодействия с хеморецептором (0,08 с);
- 3) высокая избирательная чувствительность хеморецепторов (они взаимодействуют только со специфичным медиатором);
- 4) утомляемость, вызванная расходом медиатора.
Электрические синапсы – беспузырьковые, характеризуются узкой синаптической щелью и отсутствием специфических хеморецепторов. Они обеспечивают передачу нервных импульсов без синаптической задержки в обоих направлениях, т.е. закон динамической поляризации синапса на них не распространяется.
По функции синапсы делят на возбуждающие и тормозные. Химические синапсы обеспечивают проведение как возбуждающих, так и тормозных нервных импульсов. Электрические синапсы проводят только возбуждающие импульсы.
Эффекторы – это нейротканевые синапсы, осуществляющие передачу нервного импульса с нейрона на ткани рабочего органа.
В поперечнополосатых, или скелетных, мышцах эффекторы представлены моторными бляшками (рис. 1.4). Мякотное нервное волокно вблизи моторной бляшки теряет миелиновый слой и распадается на терминальные ветви. Последние погружаются в складки сарколеммы мышечного волокна. В нервно-мышечном синапсе между терминалью аксона и сарколеммой мышечного волокна имеется синаптическая щель, ширина которой составляет от 10 до 20 нм.
Медиатором в этих синапсах, как правило, является ацетилхолин, который действует возбуждающе. В нейро- ткансвых синапсах вегетативной нервной системы нервный импульс передается с помощью медиаторов, в качестве которых могут выступать различные химически активные вещества, чаще ацетилхолин, норадреналин, аденозинтрифосфорная кислота и др. Именно медиаторы определяют конкретную реакцию на раздражение и ее продолжительность.

Рис. 1.4. Строение моторной бляшки:
1 – миофибрилла; 2 – синаптическая щель; 3 – митохондрии; 4 – сарколемма; 5 – нервные волокна; 6 – миелиновая оболочка; 7 – пресинантическая мембрана; 8 – постсинаптическая мембрана
Взаимодействия между глиальными и нервными клетками отчетливо проявляются в процессах развития и структурной организации нервных волокон. Нервным волокном называется отросток нервной клетки, окруженный глиальной оболочкой.
Непосредственно сам отросток называют еще осевым цилиндром, а клетки глиальной оболочки — нейролеммоцитами. Различают миелиновые (мякотные) и безмиелиновые (безмякотные) нервные волокна.
В безмиелиновых нервных волокнах отростки нервных клеток погружены в углубления на поверхности нейролеммоцитов, имеющих вид желоба. Погруженный в тело глиальной клетки нервный отросток ограничен как собственной плазмолеммой, так и внешней мембраной нейролеммоцита. Он как бы подвешен на двухлистковой ее складке. Эти складки мембран (своеобразные ультраструктурные "брыжейки") называют мезаксонами. Безмиелиновые волокна могут включать несколько осевых цилиндров.
Миелиновое нервное волокно состоит из нервного отростка и нейролеммоцитов (шванновских клеток). Осевой цилиндр не просто погружен в цитоплазму нейролеммоцита, а окружен спиральной слоистой оболочкой (миелином), образованной наматыванием мезаксонов нейролеммоцитов при их вращении вокруг отростка нервной клетки. В миелиновой оболочке обнаружены липиды, щелочной белок миелина, маркерный белок S100 и др.
Высокое содержание липидов (почти 2/3 массы миелина) выявляется при обработке препаратов четырехокисью осмия, окрашивающей миелиновую оболочку в темно-коричневый цвет. По ходу миелинового волокна имеются сужения — узловые перехваты (перехваты Ранвье). Они соответствуют границе смежных нейролеммоцитов. Каждый межузловой сегмент оболочки волокна представлен одним нейролеммоцптом. Миелиновые волокна толще безмиелиновых. Скорость проведения нервного импульса по ним составляет 5-120 м/с, тогда как по безмиелиновым волокнам импульс проводится со скоростью 1-2 м/с.
Сложные взаимоотношения между нервными и глиальными клетками складываются при формировании чувствительных нервных окончаний (рецепторов) и двигательных нервных окончаний (эффекторов).
Нервные окончания — концевой аппарат нервных волокон, формирует межнейрональные контакты, или синапсы, рецепторные (чувствительные) окончания и двигательные (эффекторные) окончания.
Синапс (от synapsis — соединение) — специализированный для передачи нервных импульсов контакт между двумя нейронами или между нейроном и эффектором. Процессы возбуждения нейронов, возникновение импульсов и распространение их по отросткам связаны с изменениями в плазмолемме. Она является структурной основой возникновения и передачи потенциалов действия. Плазмолемма имеет существенные особенности строения и функции в участках, входящих в состав синапсов.
Межнейрональные синапсы бывают нескольких видов: аксосоматические (между аксоном одного нейрона и телом другого нейрона); аксодендритические (между аксоном одного нейрона и дендритом другого нейрона); аксоаксональные (между аксонами двух нейронов). Описаны также синапсы соматосоматические, дендродендритические и др.
Все синапсы по механизму передачи импульсов между нервными клетками подразделяются на 3 типа: синапсы с химической передачей, электротонические и смешанные синапсы. Типичный синапс с химической передачей состоит из пресинаптической и постсинаптической частей, а также синаптической щели. Пресинаптическая часть включает концевое расширение аксона, ограниченное пресинаптической мембраной. Специфическими структурами этой части являются синоптические пузырьки, содержащие нейромедиаторы. Пузырьки бывают со светлым и электронно-плотным содержимым и называются в связи с этим агранулярными и гранулярными.
По форме они подразделяются на круглые и уплощенные. На внутренней поверхности пресинаптической мембраны расположены конусовидные электронно-плотные образования — пресинаптические уплотнения. В цитоплазме пресинаптической части имеются митохондрии. Синаптическая щель размером 20-30 нм содержит филаменты, связывающие наружные слои плазмолеммы контактирующих нейронов.
Постсинаптическая часть в составе плазмолеммы второго нейрона имеет рецепторы к медиатору, который выделяется в синаптическую щель при деполяризации мембраны первого нейрона. Внутренняя поверхность постсинаптической мембраны характеризуется наличием электронно-плотного слоя цитоплазмы — постсинаптические уплотнения.
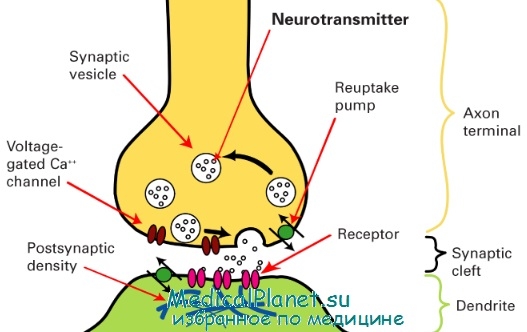
Схема строения синапса
Нервная ткань развивается из эктодермы, является основным компонентом нервной системы. Основными свойствами нервной ткани являются возбудимость и проводимость.
Нервная ткань состоит из нервных клеток (нейронов) и межклеточного вещества (нейроглии). Нейроны способны воспринимать, анализировать раздражение, приходить в состояние возбуждения, генерировать нервные импульсы и передавать их другим нейронам либо рабочим органам.
Нейроны представляют собой отростчатые клетки, размеры которых колеблются в значительных пределах. По форме нервные клетки также различны. Отростки являются проводниками нервных импульсов. Различают два вида отростков:
· аксон – длинный отросток, обеспечивает проведение импульса от нервной клетки к рабочему органу или другой клетке; каждая нервная клетка имеет только один аксон;
· дендрит – короткий, древовидно-ветвящийся отросток, воспринимает импульсы и проводит к телу нейрона; количество дендритов у разных нейронов различное.
Нейрон имеет типичное клеточное строение. В цитоплазме клеток присутствуют специфические органеллы:
· нейрофибриллы – участвуют в проведении нервного импульса;
· тигроидное (базофильное) вещество – представляет собой зернистость, образующую нерезко отграниченные глыбки, лежащие в теле клетки и дендритах. Оно меняется в зависимости от функционального состояния клетки. В условиях перенапряжения, травмы (перерезка отростков, отравление, кислородное голодание и др.) глыбки распадаются и исчезают. Этот процесс получил название хроматолиза, или тигролиза, т.е. растворения тигроидного вещества. По морфологическим изменениям базофильного вещества можно судить о состоянии нервных клеток в условиях патологии и эксперимента.
Нейрон является структурно-функциональной единицей нервной ткани. С помощью своих отростков он взаимодействует с другими нейронами, образуя рефлекторные дуги – нейронные цепи, из которых построена нервная система.
В организме человека нервный импульс обычно передается от одного нейрона к другому либо на рабочий орган не напрямую, а через химический посредник – медиатор.
Нейроны классифицируют по трем основным группам признаков: морфологическим, функциональным и биохимическим.
Морфологическая классификация (по особенностям строения):
ü по количеству отростков нейроны делятся на:
- униполярные (с одним отростком) – встречаются в эмбриогенезе;
- биполярные (с двумя отростками) – некоторые нейроны сетчатки глаза, нейроны спирального и вестибулярного ганглиев;
- псевдоуниполярные (ложно униполярные) – к ним относятся все рецепторные нейроны спинальных и краниальных ганглиев. Аксон и дендрит начинается от общего выроста тела клетки с последующим Т-образным делением;
- мультиполярные (имеют три и более отростка) – преобладают во всех отделах ЦНС и в вегетативных ганглиях периферической нервной системы;
ü по форме – описано до 80 вариантов нейронов (звездчатые, пирамидальные, грушевидные, веретеновидные и др.).
Функциональная классификация (в зависимости от выполняемой функции и места в рефлекторной дуге различают нейроны):
- рецепторные (чувствительные, афферентные) – с помощью дендритов воспринимают воздействия внешней или внутренней среды, генерируют нервный импульс и передают его другим типам нейронов; встречаются только в спинальных ганглиях и чувствительных ядрах черепно-мозговых нервов;
- эффекторные (эфферентные) – передают возбуждение на рабочие органы (мышцы или железы); располагаются в передних рогах спинного мозга и вегетативных нервных ганглиях;
- вставочные (ассоциативные) – располагаются между рецепторными и эффекторными нейронами; по количеству их больше всего, особенно в ЦНС;
- секреторные (нейроэндокриноциты) – специализированные нейроны, по своей функции напоминающие эндокринные клетки. Они синтезируют и выделяют в кровь нейрогормоны, расположены в гипоталамической области головного мозга; регулируют деятельность гипофиза, а через него и многие периферические эндокринные железы.
Медиаторная классификация (по химической природе выделяемого медиатора):
- холинергические (медиатор ацетилхолин);
- аминергические (медиаторы – биогенные амины, например, норадреналин, серотонин, гистамин);
- ГАМК-эргические (медиатор – гамма-аминомасляная кислота);
- пептидергические (медиаторы – пептиды, например, опиодные пептиды, субстанция Р, холецистокинин и др.);
- пуринергические (медиаторы – пуриновые нуклеотиды, например, аденозин) и др., а также нейроны, которые в качестве медиатора используют аминокислоты (глицин, глутамат, аспартат).
Нейроглия(межклеточное вещество) органически связана с нервными клетками, имеет клеточное строение и осуществляет трофическую, секреторную, защитную, разграничительную и опорную функцию. Она поддерживает постоянство среды вокруг нейронов. Клетки нейроглии делятся на две группы: макроглию и микроглию.
Макроглия. Клетки макроглии бывают трех типов:
· эпендимоциты – выстилают каналы и желудочки спинного и головного мозга, по которым циркулирует спинномозговая жидкость (ликвор). В желудочках мозга находятся сосудистые сплетения. Они покрыты специализированными секреторными эпендимоцитами, участвующими в образовании ликвора.
· астроциты – различают протоплазматические и волокнистые астроциты. Протоплазматические астроциты имеют короткие толстые отростки. Они расположены в сером веществе мозга, выполняют разграничительную и трофическую функции. Волокнистые астроциты находятся в белом веществе, имеют многочисленные тонкие длинные отростки, которые оплетают кровеносные сосуды мозга, образуя периваскулярные глиальные пограничные мембраны. Их отростки также изолируют синапсы. Таким образом, они изолируют нейроны и кровеносные сосуды и участвуют в образовании гематоэнцефалического барьера, обеспечивают обмен веществ между кровью и нейронами. Они также участвуют в образовании оболочек мозга и выполняют опорную функцию (образуют каркас мозга).
· олигодендроциты – имеют мало отростков, окружают нейроны, выполняя трофическую (участие в питании нейронов) и разграничительную функции. Олигодендроциты, расположенные вокруг тел нейронов, называются мантийными глиоцитами. Олигодендроциты, расположенные в периферической нервной системе и образующие оболочки вокруг отростков нейронов, называют леммоцитами (шванновскими клетками).
Нервные волокна – это отростки нейронов, покрытые глиальными оболочками. Отростки нейронов лежат внутри нервных волокон и называются осевыми цилиндрами. Их окружают глиальные клетки – олигодендроциты, которые здесь называются леммоцитами (оболочечными клетками), или шванновскими клетками.
По гистологическому строению нервные волокна бывают миелиновые (мякотные) и безмиелиновые (безмякотные).
Миелиновые нервные волокна имеют оболочку из двух слоев: внутренний называется миелиновым (мякотным) и представлен липопротеидным веществом – миелином; наружный – шванновскими клетками и называется нейролеммой. Миелин служит для защиты, питания и изоляции нервных волокон. Через равные промежутки миелиновая оболочка прерывается, образуя перехваты Ранвье. Такие волокна образуют белое вещество спинного и головного мозга, входят в периферические нервы.
Безмиелиновые (безмякотные) нервные волокна преимущественно входят в состав вегетативной нервной системы. Оболочка состоит из клеток нейроглии – шванновских клеток, плотно прилегающих друг к другу.
По функции нервные волокна бывают двигательные и чувствительные.
Нервные волокна заканчиваются нервными окончаниями. По функции нервные окончания делятся на:
· рецепторы – чувствительные нервные окончания образованы концевыми разветвлениями дендритов чувствительных нейронов. Они воспринимают раздражения из внешней среды – экстерорецепторы и из внутренних органов – интерорецепторы.
· эффекторы – двигательные нервные окончания являются концевыми разветвлениями аксонов двигательных клеток, посредством которых импульс передается на ткани рабочих органов. Двигательные нервные окончания скелетных мышц называются моторными бляшками.
Особую группу нервных окончаний образуют соединения (контакты) между нервными клетками – межнейрональные синапсы.
Читайте также: