
Проведение нервных импульсов у животных
Раздражимость, или чувствительность — характерная черта всех живых организмов, означающая их способность реагировать на сигналы, или раздражители. Сигнал воспринимается рецептороми передается с помощью нервов и(или) гормонов к эффектору, который осуществляет специфическую реакцию, или ответ.
Животные в отличие от растений имеют две взаимосвязанные системы координации функций — нервную и эндокринную (гормональную). Сравнение их представлено в табл. 17.1. Разница между нервной и гормональной регуляцией хорошо прослеживается на примере регуляции секреции различных веществ в пищеварительном тракте (разд. 8.4). У животных и нервная, и гормональная системы развивались параллельно. У растений имеется только химическая координация функций, аналогичная эндокринной. Логично предположить, что дополнительное существование у животных нервной системы связано с особенностями их питания, а именно с потребностью в активном добывании пищи. Для этого же необходимы сенсорные и локомоторные органы, работа которых контролируется нервной системой.
Таблица 17.1. Сравнение нервной и гормональной регуляции у животных
Электрическое и химическое проведение (нервные импульсы и нейромедиаторы в синапсах)
Химическое проведение (гормоны) по кровеносной системе
Быстрые проведение и ответ
Более медленное проведение и отсроченный ответ (исключением является адреналин)
В основном кратковременные изменения
В основном долговременные изменения
Специфический путь распространения сигнала (по нейронам)
Неспецифический путь распространения (с кровью по всему телу) к специфической мишени
Ответ часто узко локализован (например, одна мышца)
Ответ может быть крайне генерализованным (например, рост)
Нервная система состоит из высокоспециализированных клеток со следующими функциями:
1) восприятие средовых сигналов. У многоклеточных организмов для этого существуют модифицированные нервные клетки — рецепторы. Их структура и функция описаны в разд. 17.5;
2) преобразование сигналов в электрические импульсы — так называемая трансдукция;
3) проведение импульсов, часто на значительные расстояния, к другим специализированным клеткам — эффекторам, которые, получив сигнал, способны дать адекватный ответ. Структура и функция эффекторов вкратце описаны в разд. 17.5 и гл. 18.
Связь между рецепторами и эффекторами осуществляют особые клетки — нейроны. Они являются основными структурными и функциональными элементами нервной системы и их разветвленные отростки пронизывают все тело животного, образуя сложную сеть передачи сигналов.
Типы нейронов - резюме
Строение различных типов нервных клеток описано в разд. 6.6. Речь идет о сенсорных (афферентных), двигательных (эфферентных) и вставочных нейронах (рис. 6.27). Двигательные нейроны называют также мотонейронами, а вставочные нейроны — интернейронами. Во всех нервных клетках можно различить компактное тело (перикарион), содержащее ядро, и отходящие от тела тонкие отростки. Отросток, по которому нервные импульсы идут от перикариона, называется аксоном. Он оканчивается синаптическим вздутием, так называемой синаптической бляшкой. К перикариону импульсы поступают по дендрону, разделенному на тонкие ветви — дендриты, которые получают сигналы от других клеток нервной системы 1 . Многие аксоны окружены белково-липидной миелиновой оболочкой и называются миелинизированными. Иногда дендрон и аксон образуют единое волокно, отходящее от перикариона с одного бока. Другой особый тип нейронов — биполярный — описан в этой главе ниже. У него дендрон и аксон отходят от тела клетки с противоположных ее сторон.
Измерение электрической активности нейронов
То, что сигналы, вызывающие мышечное сокращение и секреторную активность желез, передаются по нейронам в форме электрических импульсов, известно уже более 200 лет. Однако механизм распространения этих импульсов был выяснен лишь в последние полвека, после того как у кальмара обнаружили аксоны толщиной около миллиметра. Такая толщина этих гигантских аксонов (они проводят импульсы, связанные с реакцией бегства) позволила вводить в них электроды и регистрировать импульсы, т. е. проводить электрофизиологические эксперименты.
На рис. 17.1 изображен прибор, используемый в настоящее время для изучения электрической активности нейронов. Его важнейшую часть составляет микроэлектрод — стеклянная трубочка, вытянутая на конце в тончайший капилляр диаметром 0,5 мкм и заполненная раствором электролита (например 3М КС1). Этот микроэлектрод вводят в аксон, а второй электрод, имеющий вид металлической пластинки, помещают в физиологический (солевой) раствор, омывающий нейрон. Оба электрода подсоединены к усилителю, замыкающему цепь. Сигнал, усиленный примерно в 1000 раз, передается на двухлучевой катодный осциллограф. Микроэлектрод перемещают с помощью микроманипулятора — специального устройства, снабженного винтами, напоминающими винты микроскопа. Вращая их, можно с большой точностью регулировать положение микроэлектрода.
Когда кончик микроэлектрода проходит сквозь плазматическую мембрану аксона, лучи осциллографа раздвигаются. Расстояние между лучами на экране показывает разность потенциалов между электродами в цепи. Разность потенциалов покоящейся клетки называется потенциалом покоя (см. ниже). В сенсорных клетках, нейронах и мышечных клетках измеряемая описанным способом величина меняется в зависимости от их активности, поэтому такие клетки называют возбудимыми. На мембранах всех остальных живых клеток тоже существует подобная разность потенциалов, известная как мембранный потенциал, но в этих клетках она остается постоянной, поэтому их называют невозбудимыми клетками.

Рис. 17.1. Схема установки для регистрации электрической активности аксона изолированного нейрона. Датчик сигналов дает ток, генерирующий в аксоне потенциал действия, который воспринимается введенным в аксон микроэлектродом. Возникающий сигнал передается от него на двухлучевой катодный осциллограф.
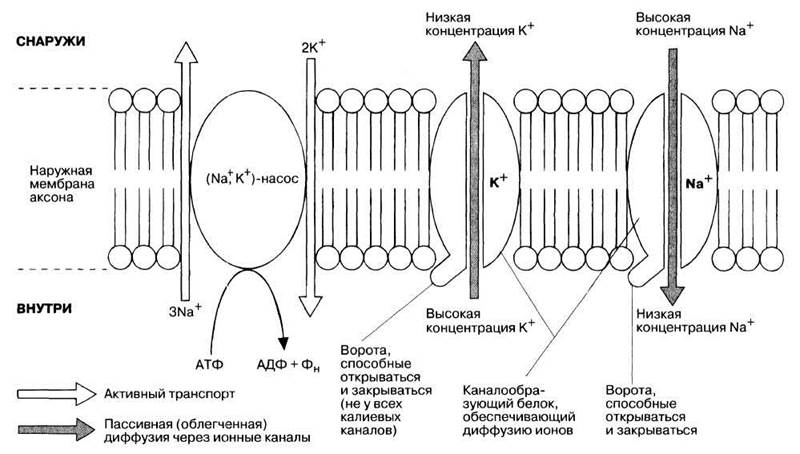
Разность потенциалов между наружной и внутренней поверхностями клеточной мембраны (трансмембранный потенциал) обычно, в том числе и у нейронов в покое, устанавливается таким образом, что внутренняя часть клетки заряжена отрицательно относительно окружающей ее среды. Этот феномен называют поляризацией мембраны. Потенциал покоя нейрона составляет примерно —70 мВ. Он поддерживается благодаря активному транспорту и диффузии ионов через мембрану, что было продемонстрировано Кертисом и Коулом в США и Ходжкином и Хаксли в Англии в конце тридцатых годов прошлого века на аксонах кальмара (табл. 17.2).
Таблица 17.2. Концентрация ионов (ммоль/л) во внеклеточной среде и внутриклеточной жидкости гигантского аксона кальмара (приблизительные значения из Hodgkin , 1958)
А - (органические анионы)
В цитоплазме аксона (аксоплазме) больше ионов калия (К + ) и меньше — натрия ( Na + ). В окружающей аксон среде наблюдается обратная картина, т. е. низкая концентрация К + и высокая — Na + . (Распределение хлорид-ионов и других ионов в последующем описании не учитывается, поскольку оно не играет существенной роли в нервном проведении.) Таким образом, существуют трансмембранные электрохимические градиенты ионов, обусловленные их электрическими и химическими свойствами. Электрические свойства ионов определяются их зарядами: одноименные заряды отталкиваются, противоположные — притягиваются. На движение ионов влияет также их концентрация в растворе. Ион имеет тенденцию перемещаться по градиенту концентрации, т. е. туда, где она ниже. Результирующее движение зависит от соотношения зарядов и концентраций по обе стороны мембраны.
Активному транспорту ионов противостоит их пассивная трансмембранная диффузия по электрохимическому градиенту через специфические ионные каналы, образованные особыми белками (см. рис. 17.2). Скорость этой диффузии определяется проницаемостью мембраны для данного иона. У аксона она в 20 раз выше для К + , чем для Na + . Следовательно, пассивная потеря аксоном К + идет быстрее, чем пассивное поступление Na + за счет диффузии. В результате в аксоне становится меньше катионов и создается отрицательный заряд. Это и есть потенциал покоя, величина которого определяется главным образом электрохимическим градиентом К + .
Теперь рассмотрим, как эти клетки генерируют нервные импульсы. В принципе нервные импульсы возникают при изменении проницаемости мембраны нервных клеток для К + и Na + , что приводит к изменению разности потенциалов на мембране и к появлению так называемого потенциала действия.
При стимуляции аксона электрическим током (рис. 17.3) потенциал на внутренней поверхности мембраны меняется с —70 на +40 мВ. Это изменение полярности носит название потенциаладействия и регистрируется на двухлучевом осциллографе в виде кривой, представленной на рис. 17.3.

Рис. 17.3. Типичный потенциал действия в аксоне кальмара.
Причина такого явления — изменение натриевых каналов. Они (как и некоторые калиевые) снабжены воротами, т. е. могут открываться и закрываться особыми полипептидными цепями. При закрытых воротах проницаемость мембраны для ионов, проходящих через соответствующие каналы, падает, а при открытых — увеличивается. Когда клетка находится в состоянии покоя, натриевые ворота закрыты, поэтому ее мембрана в 20 раз более проницаема для ионов калия.
Потенциал действия возникает в результате внезапного кратковременного открывания натриевых ворот. Это происходит в ответ на сигнал, приводящий к пороговой деполяризации мембраны аксона, т. е. к уменьшению его трансмембранной разности потенциалов на небольшую, но достаточную для данной реакции величину. Открывание ворот увеличивает проницаемость мембраны для ионов натрия, и они поступают в клетку путем диффузии. Это увеличивает в ней количество положительных ионов, т. е. приводит к еще большей деполяризации: трансмембранный потенциал меняется с отрицательного до нулевого (полностью деполяризованное состояние), а затем становится положительным. Поскольку натриевые ворота чувствительны к деполяризации, в ходе нее их открывается все больше. Это в свою очередь усиливает поступление в клетку натрия и ведет к дальнейшей деполяризации. Таким образом, описанные два процесса стимулируют друг друга и развиваются по механизму так называемой положительной обратной связи. Происходит взрывообразное ускорение потока натрия внутрь аксона (важно помнить, что все эти события разворачиваются в течение тысячных долей секунды). Максимальная трансмембранная разность потенциалов достигает +40 мВ, как показано на рис. 17.4, А. Она соответствует максимальной концентрации ионов натрия внутри клетки. Амплитуда изменения разности потенциалов составляет от—70 до +40 мВ, т. е. равна 110 мВ. Как показывают расчеты, для этого нужно, чтобы в аксон поступило совсем небольшое количество ионов натрия — порядка одной миллионной доли уже находившихся внутри.
Вскоре после открывания натриевых ворот деполяризация мембраны заставляет открываться и калиевые ворота. В результате усиливается диффузия ионов калия из клетки (рис. 17.4, Б), а поскольку у них такой же заряд, как у ионов натрия, их выход делает внутреннюю среду аксона менее положительной — начинается процесс реполяризации, т. е. возвращения к исходному потенциалу покоя.
Из сказанного выше следует, что потенциал покоя определяется в основном ионами калия, а потенциал действия — ионами натрия (рис. 17.4).

Рис. 17.4. Изменения различных свойств наружной мембраны аксона при распространении по нему нервного импульса (потенциала действия). А. Изменения мембранного потенциала, т. е. изменения электрических свойств мембраны. Б. Изменения ионной проводимости. В. Суммарное распределение зарядов по обеим сторонам мембраны аксона во время возникновения потенциала действия.
Нервные импульсы представляют собой потенциалы действия, распространяющиеся вдоль аксона в виде волны деполяризации. Наружная поверхность аксона в месте возникновения потенциала действия заряжена отрицательно.
17.1. Назовите две причины стремительного поступления Na + в аксон при повышении проницаемости мембраны аксона для Na + .
17.2. Если бы проницаемость мембраны для ионов натрия и калия увеличивалась одновременно, то как это сказалось бы на потенциале действия?
17.3. В 1949 г. Ходжкин и Катц исследовали влияние ионов натрия на возникновение потенциала действия в аксонах кальмара. Внутриклеточные микроэлектроды регистрировали потенциалы действия в аксонах, помещенных в изотонические растворы морской воды, содержащие разные концентрации ионов натрия. Результаты экспериментов приведены на рис. 17.5. Какие кривые соответствуют обычной морской воде и морской воде в разведениях 1:2 и 1:3? Объясните влияние этих растворов на потенциалы действия.

Рис. 17.5. Мембранные потенциалы в аксонах кальмара, помещенных в морскую воду с различной концентрацией ионов натрия.
Особенности потенциалов действия
РАСПРОСТРАНЕНИЕ (ПРОВЕДЕНИЕ) НЕРВНЫХ ИМПУЛЬСОВ. Нервный импульс представляет собой волну деполяризации, распространяющуюся по поверхности нейрона. Распространение происходит вследствие самогенерирования потенциалов действия за счет поступающих в аксон ионов натрия. Поступившие ионы натрия создают зону положительного заряда внутри клетки, что приводит к возникновению локальной электрической цепи, по которой течет местный ток между этой и соседней отрицательно заряженной зоной. Местный ток снижает мембранный потенциал в этой зоне, и в результате деполяризации здесь повышается проницаемость мембраны для натрия и в свою очередь генерируется потенциал действия. Последовательная деполяризация все новых и новых участков мембраны приводит к тому, что потенциал действия распространяется по аксону все дальше. Теоретически потенциалы действия могут передаваться на любые расстояния, иными словами, они не затухают. Причина этого кроется в том, что локальное изменение концентрации ионов в каждой точке обусловливает независимое самогенерирование потенциала действия. До тех пор пока снаружи и внутри аксона существует необходимая разница в концентрации ионов, потенциал действия в одной точке мембраны будет порождать потенциал действия в соседнем его участке.

Как же на самом деле организм различает силу раздражителя (сигнала)? Оказывается, что интенсивность сигнала влияет на число потенциалов действия, возникающих в единицу времени, т. е. чем сильнее сигнал (в определенных пределах), тем чаще генерируются нервные импульсы. Таким образом, речь идет о своего рода частотном коде.
Например, частота 10 имп/с в соответствующем аксоне поддерживает тонус (состояние частичного напряжения — готовности к быстрому действию) двуглавой мышцы (бицепса). Примерно 50 имп/с необходимы для ее нормального сокращения при движении руки. Большинство аксонов могут проводить сигналы в диапазоне 10—100 имп/с, хотя иногда в естественных условиях эта частота достигает 500 имп/с, а в экспериментальных условиях бывает еще больше.
СКОРОСТЬ ПРОВЕДЕНИЯ. У позвоночных подавляющая часть нервных волокон, особенно в спинномозговых и черепных нервах, окружены миелиновой оболочкой, образованной шванновскими клетками (рис. 6.30 и разд. 6.6.1). Миелин — это материал белково-липидной природы, обладающий высоким электрическим сопротивлением и действующий как изолятор, подобно резиновому или пластиковому покрытию электрического провода. Суммарное сопротивление мембраны аксона и миелиновой оболочки очень велико, но там, где в миелиновой оболочке имеются разрывы, называемые перехватами Ранвье, сопротивление току между аксоплазмой и внеклеточной жидкостью меньше. Только в этих участках замыкаются местные цепи, и именно здесь через мембрану аксона проходит ток, генерирующий следующий потенциал действия. В результате импульс перескакивает от одного перехвата Ранвье к другому и пробегает по миелинизированному аксону быстрее, чем серия меньших по величине местных токов в немиелинизированном нервном волокне. Такой способ распространения потенциала действия, называемый сальтаторным (от лат. saltare — прыгать), может обеспечивать проведение импульса со скоростью 120 м/с (рис. 17.7).
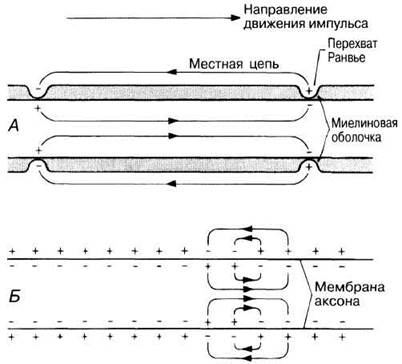
В немиелинизированных аксонах, типичных для беспозвоночных, скорость распространения потенциалов действия зависит от сопротивления аксоплазмы. Это сопротивление в свою очередь зависит от диаметра аксона — чем меньше диаметр, тем больше сопротивление. В тонких аксонах (
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Одностороннее проведение возбуждения. В нервных центрах прохождение импульсов возбуждения по цепи нейронов осуществляется только в одном направлении: от чувствительного нерва через промежуточные к двигательному, а от двигательного — к органу-исполнителю. Это обусловлено односторонней синаптической передачей импульсов от одной нервной клетки к другой с помощью медиатора, который выделяется концевым аппаратом аксона и содержится лишь в пресинаптической щели. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление. Это свойство обеспечивает координирующую роль центральной нервной системы и способствует замыкательной функции условно-рефлекторных связей.
Замедление проведения возбуждения.
Это свойство называется центральной задержкой, или латентным (скрытым) периодом рефлекса. Центральная задержка обусловлена более медленным проведением нервных импульсов через синапсы.
Интервал от начала раздражения рецептора до появления ответной реакции — 0,2–0,5 секунды. Чем сложнее рефлекс, тем длительнее центральная задержка. Дрессировщику необходимо учитывать скрытый период при выработке условного рефлекса и подкрепляющий раздражитель надо применять не раньше 0,5 секунды после воздействия сигнального раздражителя. Скрытый период рефлекса увеличивается при утомлении и заболевании животного.
Присуще всем нервным центрам и характеризуется тем, что на короткое раздражение проявляется длительная ответная реакция, т. е. рефлекторный акт длится еще некоторое время после того, как раздражение рецептора прекращено. Такое последействие объясняется тем, что нервные импульсы от рецепторов поступают к двигательным центрам по различным нервным путям неодновременно: по коротким быстрее, чем по длинным. Запаздывающие импульсы поддерживают возбужденное состояние соответствующего нервного центра. Остаточное возбуждение в нервных центрах может сохраняться до 2 секунд и тем самым способствовать лучшей замыкательной функции при образовании условных рефлексов. Кроме того, в практике дрессировки нужно помнить, что после окончания действия любого раздражителя требуется некоторое время для спада остаточного возбуждения нервного центра и освобождения рефлекторных путей для новой рефлекторной деятельности.
Происходит путем накопления слабых допороговых раздражений до критического потенциала, способного вызвать возбуждение нервного центра. Различают пространственную и временную суммацию. Пространственная суммация наблюдается в случае одновременного воздействия раздражителей допороговой силы на несколько чувствительных нейронов. Поступившие импульсы слабой силы суммируются в нервном центре и вызывают возбуждение. Суммация во времени происходит при активизации одного и того же чувствительного нейрона серией последовательных раздражений допороговой силы. Слабые импульсы от предыдущих раздражений накладываются друг на друга, суммируются и вызывают рефлекс. Суммация в пространстве и во времени в нервных центрах происходит одновременно. Они дополняют и усиливают друг друга и, например, увеличивают обонятельную и слуховую чувствительность у собаки, что так необходимо при работе по чутью, когда исключительно слабые раздражения запаховых частиц (1 молекула в одном литре воздуха) вызывают запаховые ощущения и соответствующие ответные реакции.
Свойство нервных центров изменять частоту и силу передающихся импульсов, т. е. трансформировать. Проявляется в активизации и перестройке нейронов на более высокий или более низкий ритм и изменении их лабильности, обеспечивает взаимодействие и установление связи между различными нервными центрами и другими отделами нервной системы, что имеет важное значение в замыкании условно-рефлекторных связей при дрессировке собак.
Свойство нервных центров обеспечивать высокую возбудимость и эффективность рефлекторной деятельности нервной системы. Сущность облегчения заключается в том, что после каждого раздражения в нервном центре повышается возбудимость к повторным раздражениям, следующим через небольшие промежутки времени. Один поток импульсов как бы облегчает действие другого и способствует образованию условного рефлекса.
Способность одних нервных центров повышать возбудимость других и вступать во взаимодействие через обмен и передачу импульсов возбуждения между центрами по нейронным путям. Процесс образования временной связи рассматривается как результат взаимодействия двух очагов возбуждения в коре, следствием чего является проторение пути между этими корковыми пунктами. Проходимость пути обусловлена функциональными и морфологическими изменениями в синапсах.
При сильном и длительном раздражении импульсы, поступившие в нервную систему, не ограничиваются возбуждением одного рефлекторного центра, а распространяются на другие центры. Чем сильнее раздражение, тем больше нервных центров охватывает иррадиация. Возбуждение большого количества различных нервных центров позволяет отобрать из них наиболее нужные и установить между ними новые функциональные связи — условные рефлексы. Большинство двигательных условных рефлексов формируется благодаря иррадиации возбуждения. Излишняя иррадиация возбуждения центральной нервной системы нарушает уравновешенность поведенческих реакций, приводит к расстройству рефлекторной деятельности. Иррадиация возбуждения ограничивается и уравновешивается торможением.
Тормозной процесс — необходимое условие в координации нервной деятельности. Торможение возникает в определенных нервных структурах под влиянием волны возбуждения, которая подавляет другое возбуждение. При торможении выключается деятельность ненужных в данный момент органов и предохраняется от чрезмерного перенапряжения работа нервных центров. В нервной системе вырабатываются тормозные условные рефлексы: прекращения нежелательных действий, выдержки, дифференцировки и др.
Преобладание активности одних нервных центров над активностью других.
Доминирующий (господствующий) очаг возникает при определенном функциональном состоянии нервных центров. Одно из условий его образования — повышенная возбудимость нервных клеток одного или нескольких нервных центров под влиянием определенных нервных и гуморальных факторов. Возбудимость доминирующего очага усиливается суммированием импульсов, поступающих из других нервных центров, рефлекторная деятельность которых подавляется, а рефлекторная деятельность доминирующего очага заметно усиливается. Установившееся доминирование при инстинктах может быть продолжительным состоянием, которое определяет поведение животного на тот или иной срок.
Доминанту можно наблюдать и в условно-рефлекторной деятельности. Доминирующий очаг возбуждения в коре головного мозга притягивает к себе импульсы нервных центров других очагов возбуждения и тем самым способствует суммации, облегчению, проторению и замыканию условно-рефлекторных связей на сигналы дрессировщика. Доминанта по теории академика И.П. Павлова имеет главенствующее значение в механизме образования и проявления условных рефлексов. У собаки всегда сильно проявляются стойко выработанные навыки, доминирующие над другими рефлексами и обеспечивающие безотказное управление собакой при отвлекающих раздражителям. Дрессировщику необходимо учитывать свойство доминанты при подготовке собак.
Схождение поступающих по чувствительным путям импульсов возбуждения в одном промежуточном или двигательном центре. В центральной нервной системе в 4–5 раз больше чувствительных путей, чем двигательных. Поэтому к одному и тому же двигательному центру могут подходить импульсы возбуждения по многим путям. Эта особенность прохождения возбуждения по нервным центрам противоположна иррадиации и является основой для концентрации возбуждения в отдельных пунктах коры головного мозга. Конвергенция обеспечивает специализацию условного рефлекса и формирование навыка на сложные и комплексные раздражители.
Проявляется при сочетании особенно сильных раздражителей, дающих эффект меньше суммы величин этих реакций на каждый раздражитель в отдельности. Это свойство противоположно суммации. Окклюзия и пространственная суммация постоянно взаимодействуют между собой. При слабом возбуждении проявляется суммация, при сильных раздражениях — окклюзия. В дрессировке собак ошибочно применение сильных раздражителей для ускорения первоначального условного рефлекса. На громкие команды и сильные подкрепления вырабатываются, как правило, слабые условные рефлексы.
Способность перестройки функций нервных центров. Сформировавшиеся в процессе эволюции рефлекторные акты, реакции поведения и инстинкты могут перестраиваться, а нервные центры изменять свои функции. Опытами установлено, что перестройка функций нервных центров у животных происходит под регулирующим влиянием коры полушарий головного мозга. Благодаря этому свойству при изменении условий жизни меняется поведение животных, их привычки и навыки: собака и кошка могут мирно уживаться, инстинкт преследования собакой диких животных по следам заменяется реакцией поиска человека по его запаховому следу, вредные привычки, нежелательные связи у собаки могут быть исправлены систематической дрессировкой. Пластичность нервных центров позволяет при дрессировке вырабатывать сложные навыки в виде динамических стереотипов.
Нервные центры обладают свойством переходить в состояние возбуждения только при относительно длительном раздражении. Возбудившись, они сохраняют это состояние возбуждения в течение определенного времени. Такое явление И. П. Павлов назвал инертностью. Способность нервных центров длительно сохранять в себе следы возбуждения и торможения, сильно выражена в нервных клетках коры полушарий головного мозга. И. П. Павлов говорил, что если бы у нервных клеток не было инертности, у нас не было бы никакой памяти, никакой выучки, не существовало бы никаких привычек. У животных существует два вида памяти: кратковременная и долговременная. Оба вида памяти обеспечивают возможность выработки у животных условных рефлексов и формирования стойких навыков. Кратковременная память у собаки проявляется в течение нескольких минут, долговременная, или долгосрочная, память — через много дней, месяцев и даже лет. Оба вида памяти у животных выражаются в репродукции образа воспринимаемых отдельных раздражителей или целостного объекта.
Нервные центры обладают свойством постоянно находиться в состоянии незначительного возбуждения при относительном рефлекторном покое. Они постоянно посылают импульсы, обеспечивающие тоническое сокращение скелетной мускулатуры. Тонус нервных центров поддерживается действием гуморальных веществ и непрерывным потоком импульсов, поступающих от рецепторов. Огромное значение в поддержании мышечного тонуса имеют нервные центры продолговатого, среднего и промежуточного мозга. Тоническое состояние нервных центров и мышечной системы обеспечивает устойчивую выработку условных рефлексов и хорошую работоспособность собаки. Состояние рабочего тонуса поддерживается правильно организованной дрессировкой, систематической тренировкой и регулярным использованием собаки на службе.
Перечисленные свойства нервных центров обеспечивают их функциональное назначение в нервной системе и организме.
из книги Арасланов Филимон, Алексеев Алексей,
Шигорин Валерий "Дрессировка собак"
при использовании материалов
ссылка на ресурс wolcha.ru
Новость отредактировал: maugli - 11-10-2019, 07:19
О том, что нервные сигналы передаются по нейронам в виде электрических импульсов, влияющих на сокращение мышц и секреторную активность желез, было известно уже более 200 лет назад. Однако механизм возникновения и распространения этих импульсов был выяснен только в последние 40 лет, после того как у кальмара были открыты гигантские аксоны толщиной около миллиметра. Они иннервируют мускулатуру мантии и вызывают ее быстрое сокращение, когда животному нужно спасаться от врага. Большая толщина этих аксонов позволила провести на них некоторые из самых ранних электрофизиологических исследований.
На рис. 16.1 изображен прибор, применяемый в настоящее время для изучения электрической активности нейронов. Его важнейшую часть составляет микроэлектрод - стеклянная трубочка, вытянутая на конце в капилляр диаметром 0,5 мкм и заполненная раствором, проводящим ток, например 3 М КС1. Этот микроэлектрод вводят в аксон, а второй электрод, имеющий вид маленькой металлической пластинки, помещают в солевой раствор, омывающий исследуемый нейрон. Электроды подсоединены к усилителю, замыкающему цепь. Сигнал, усиленный примерно в 1000 раз, передается на двухлучевой осциллограф. Все перемещения микроэлектрода осуществляются с помощью микроманипулятора - специального устройства, позволяющего с большой точностью регулировать положение микроэлектрода. Когда кончик микроэлектрода проходит сквозь плазматическую мембрану аксона, лучи осциллографа раздвигаются. Расстояние между лучами по-казывает разность потенциалов между двумя электродами. Эта разность называется потенциалом покоя аксона и составляет у всех исследованных видов приблизительно - 65 мВ. Таким образом, мембрана аксона поляризована, а минус перед потенциалом покоя означает, что с внутренней стороны она заряжена отрицательно по отношению к наружной поверхности. В сенсорных клетках, нейронах и мышечных волокнах эта величина во время активности изменяется, поэтому такие клетки называются возбудимыми. На мембранах всех остальных живых клеток тоже существует подобная разность потенциалов, известная как мембранный потенциал, но в этих клетках она остается постоянной, поэтому их называют невозбудимыми клетками.
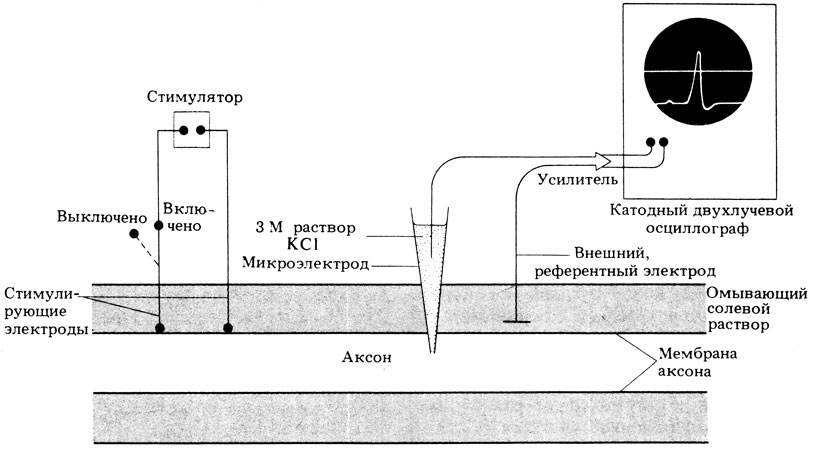
Рис. 16.1. Схема аппаратуры для регистрации электрической активности аксона отдельного нейрона. Стимулятор подает ток, возбуждающий потенциал действия, который отмечается и записывается с помощью внутриклеточных микроэлектродов и двулучевого катодного осциллографа
У большинства нейронов млекопитающих потенциал покоя остается постоянным до тех пор, пока клетка находится в неактивном состоянии из-за отсутствия стимула. Кертис и Коул в США и Ходжкин и Хаксли в Англии в конце 30-х годовустановили, что потенциал покоя имеет физико-химическую природу и обусловлен разностью ионных концентраций по обе стороны мембраны аксона и избирательной проницаемостью мембраны для ионов. Анализ жидкости, находящейся внутри аксона, и омывающей аксон морской воды показал, что между обеими жидкостями существуют электрохимические ионные градиенты (табл. 16.1).

Таблица 16.1. Концентрации ионов во внеклеточной среде и внутриклеточной жидкости аксона кальмара (приближенные величины; выражены в миллимолях на 1 кг Н2О; из Hodgkin, 1958)
В аксоплазме, находящейся внутри аксона, больше ионов К + и меньше Na + , тогда как в жидкости, омывающей аксон, наоборот, больше ионов Na + и меньше К + (распределение ионов Сl - в последующем описании не учитывается, так как оно не играет существенной роли в интересующих нас явлениях).
Указанные градиенты поддерживаются за счет активного переноса ионов против их электрохимических градиентов, который осуществляют определенные участки мембраны, называемые катионными или натриевыми насосами. Эти непрерывно действующие транспортные механизмы работают за счет энергии, освобождающейся при гидролизе АТФ; при этом происходит выведение из аксона Na+, сопряженное с поглощением К + (рис. 16.2, А).
Активному транспорту этих ионов противостоит их пассивная диффузия, поскольку они постоянно перемещаются "вниз" по электрохимическим градиентам, как показано на рис. 16.2, Б. Скорость диффузии определяется проницаемостью мембраны аксона для данного иона. Ионы К + более подвижны, и проницаемость мембраны для них в 20 раз больше, чем для Na + , поэтому К + легче выходит из аксона, чем Na + входит в него, и в результате в аксоне становится меньше катионов и создается отрицательный заряд. Величина потенциала покоя определяется главным образом электрохимическим градиентом К + .
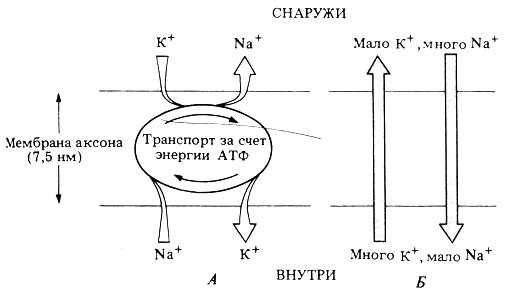
Рис. 16.2. Активное (А) и пассивное (Б) перемещение ионов, связанное с созданием отрицательного потенциала внутри аксона. Натриево-калиевый насос осуществляет активный перенос ионов (А), которые в то же время проходят через мембрану путем пассивной диффузии в направлении их электрохимических градиентов (Б)
Изменение проницаемости мембраны возбудимых клеток для ионов К + и Na + приводит к изменению разности потенциалов на мембране, к возникновению потенциалов действия и распространению нервных импульсов по аксону.
При стимуляции аксона электрическим током (рис. 16.3) потенциал на внутренней поверхности мембраны меняется с - 70 мВ до + 40 мВ. Это изменение полярности носит название потенциала действия (спайка) и регистрируется на двухлучевом осциллографе в виде кривой, представленной на рис. 16.3.
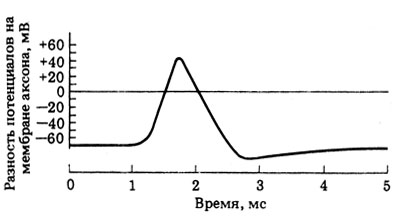
Рис. 16.3. Типичный потенциал действия в аксоне кальмара
Потенциал действия возникает в результате внезапного кратковременного повышения проницаемости мембраны аксона для ионов Na + и входа последних в аксон. Вследствие увеличения проводимости (электрический эквивалент проницаемости) для натрия число положительно заряженных ионов внутри аксона возрастает, и мембранный потенциал снижается по сравнению с величиной покоя, составляющей около - 70 мВ. Такое изменение мембранного потенциала называется деполяризацией. Повышение натриевой проводимости и деполяризация влияют друг на друга по принципу положительной обратной связи, т.е. взаимно усиливают друг друга, и в результате возникает фаза крутого подъема потенциала действия. Как показали расчеты, при деполяризации порядка 110 мВ, происходящей при развитии потенциала действия, в аксон входит сравнительно немного ионов Na + (около 10-6% от общего количества Na + , присутствующего в клетке, в зависимости от диаметра аксона). На пике потенциала действия проницаемость для натрия начинает падать (инактивация натриевых каналов), а примерно через 0,5 мс после начала деполяризации повышается проницаемость для К + , и калий выходит из аксона. По мере выхода ионов К + положительный заряд с внутренней стороны мембраны меняется на отрицательный. Эта фаза реполяризации мембраны, представленная нисходящей областью пика потенциала действия, приводит к восстановлению исходного мембранного потенциала.
Из приведенного объяснения видно, что если потенциал покоя определяется главным образом ионами К + , то потенциал действия зависит в основном от ионов Na + (рис. 16.4).
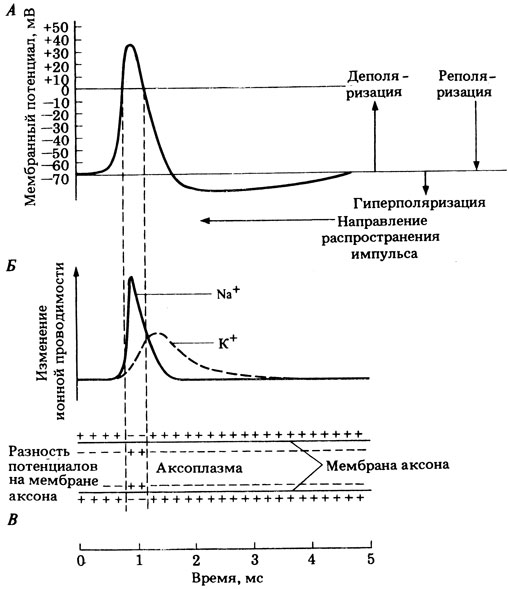
Рис. 16.4. Схема взаимоотношений между мембранным потенциалом (А), ионной проводимостью (Б) и распределением потенциалов на обеих сторонах мембраны аксона (В) во время потенциала действия
16.1. Приведите две причины резкого увеличения входа Na + в аксон при повышении проницаемости мембраны аксона для Na + .
16.2. Как изменился бы потенциал действия, если бы одновременно повысилась проницаемость мембраны для инов Na + и К + ?
16.3. В 1949 г. Ходжкин и Катц исследовали влияние ионов Na + на потенциалы действия в аксонах кальмара. С помощью внутриклеточных микроэлектродов их регистрировали в аксонах, омываемых изотоничной морской водой и той же водой в разведениях 1:2 и 1:3. Результаты представлены на рис. 16.5. Какой среде соответствует каждая из приведенных трех кривых? Объясните влияние различных растворов на потенциалы действия.
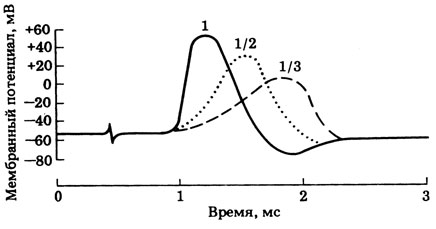
Рис. 16.5. Запись мембранных потенциалов в аксоне кальмара, омываемом обычной и разведенной морской водой
Возникновение потенциала действия. Стимуляция сенсорных клеток приводит к деполяризации их мембран, и если эта деполяризация достигнет определенной пороговой величины, она вызовет потенциал действия. Амплитуда потенциала действия (рис. 16.3) постоянна для каждого нейрона, и увеличение силы или частоты стимуляции на нее не влияет; поэтому говорят, что потенциал действия подчиняется закону "всё или ничего".
Передача нервных импульсов. В нервной системе информация передается как серия нервных импульсов - распространяющихся потенциалов действия. Нервный импульс пробегает по аксону в виде волны деполяризации. Такое распространение потенциала действия происходит благодаря натриевому току: когда ионы Na + входят в аксон, в данном участке аксона создается область положительного заряда и в местной цепи возникает ток между этим активным участком и отрицательно заряженной областью, находящейся непосредственно перед ним. Возникающий в местной цепи ток снижает мембранный потенциал в этой области, и в результате деполяризации здесь повышается проницаемость мембраны для Na + и в свою очередь возникает потенциал действия, развивающийся по принципу "все или ничего". Последовательная деполяризация все новых и новых участков мембраны ведет к тому, что потенциал действия распространяется по аксону все дальше без изменения амплитуды. Таким образом потенциалы действия могут передаваться на любые расстояния, т. е. носят незатухающий характер (бездекрементное проведение). Причина этого заключается в том, что изменение в каждой точке аксона происходит за счет локального запаса энергии в виде ионного градиента, хотя и запускается изменением в соседней точке. До тех пор пока существует необходимая разность ионных концентраций внутри и вне аксона, потенциал действия, возникающий в одном участке, будет генерировать потенциал действия в соседнем участке.
Период рефрактерности. Нервные импульсы проходят по аксону только в одном направлении. Это связано с тем, что участок, бывший ранее активным, переходит в восстановительную фазу. Во время этой фазы мембрана аксона не может отвечать на деполяризацию изменением натриевой проводимости даже в том случае, если интенсивность стимула увеличилась. Эта фаза носит название периода абсолютной рефрактерности и продолжается около 1 мс. За нею следует период относительной рефрактерности, продолжающийся 5-10 мс, в течение которого сильные стимулы могут вызывать деполяризацию. Период рефрактерности ограничивает возможную частоту нервных импульсов.
16.4. Опишите изменения, происходящие в аксоне во время периода рефрактерности.
Читайте также: