
Проведення імпульсу по нерву
Аксони та дендрити разом з оболонками, що входять до складу периферичних нервів, є нервовими волокнами. Нервові волокна, що мають мієлінову оболонку, називають мієліновими (рис. 2.22). Мієлінова оболонка створюється внаслідок того, що мієлоцит (шваннівська клітина) багаторазово обгортає осьовий циліндр, шари її зливаються і створюють щільний жировий футляр. Мієлінова оболонка через проміжки рівної довжини розривається і залишає, таким чином,

РИС. 2.22. Будова мієлінових волокон. 1 - аксон, 2 - мієлінова оболонка, 3 - шваннівська клітина, 4 - перехват Ранв'є, 5 - ядро шваннівської клітини
відкритими ділянки мембрани шириною близько 1 мкм. Ці ділянки одержали назву перехватів Ранв'є. Довжина міжперехватних ділянок пропорційна діаметру волокна.
Безмієлінові волокна не мають мієлінової оболонки і відокремлюються одне від одного тільки шваннівськими клітинами. Усередині волокна міститься осьовий циліндр з нейрофібрилами. Нейрофібрили складаються з мікротрубочок (діаметр до 30 нм) і нейрофіламентів (до 10 нм). Вони забезпечують транспортування різних речовин і деяких органел по нервових волокнах від тіла нейрона до нервових закінчень. На периферію транспортуються білки, які формують іонні канали й насоси, нейропептиди, медіатори, мітохондрії, і це явище називається аксоплазматичним транспортом, який забезпечується енергозалежним білком кінезином.
Розрізняють швидкий (близько 40 см за добу) і повільний (близько 0,5 см за добу) аксонний транспорт. Деякі речовини транспортуються у зворотному напрямку – ретроградний транспорт. До них належать ацетилхолінестераза, що бере участь у синтезі білків у тілі нейрона, токсини бактерій і вірусів, що утворюються на периферії і потрапляють до аксона, а по ньому переміщаються до тіла нейрона. За таким принципом діє правцевий токсин, що ушкоджує ЦНС, спричиняючи м'язові судоми.
Нервові волокна входять до складу нервів, які іннервують органи чуття і скелетні м'язи, внутрішні органи та судини.
Вони не можуть існувати без зв'язку з тілом нервової клітини: перерізка нерва призводить до загибелі волокон, дегенераційних змін в ефекторах. Регенерація нервів проходить повільно – зі швидкістю 0,5-4,5 мм за добу.
У процесах виникнення і проведення нервових імпульсів головну роль відіграє поверхнева мембрана осьового циліндра. Мієлінова оболонка виконує трофічну функцію, а також є електричним ізолятором. Завдяки існуванню мієлінової оболонки потенціал дії, тобто збудження, виникає тільки в перехватах Ранв'є, що має важливе значення для розповсюдження нервового імпульсу вздовж волокна.
Механізми проведення збудження нервовими волокнами – це спосіб передачі інформації на відстань. ПД, який генерується у певній точці мембрани нервового волокна, діє як стимул на сусідню (ще не збуджену) ділянку цього ж волокна завдяки місцевим кільцевим електричним струмам, що виникають між ними, і в цій новій ділянці мембрани утворюється ідентичний ПД. Період рефрактерності, що виникає під час розвитку ПД, упереджує повернення збудження до початкової ділянки мембрани та обмежує частоту генерації ПД під час поширення збудження по нервовому волокну. Таким чином, механізм проведення збудження є електричним.
Характеристика нервових волокон.
Нервові волокна- це відростки нервових клітин. До них належать аксони (ті, що передають інформацію від до центру клітини до виконавчих органів) та дендрити (ті, що передають інформацію від рецепторів до центру клітини).
Види нервових волокон:
1. Безм'якотні (безмієлінові).
2. М'якотні (мієлінові).
М'якотні нервові волокна складаються з осьового циліндру та мієлінової оболонки. У плазмі нервових волокон знаходяться численні нейрофібрили та мікротубули, через які здійснюється транспорт БАР, органічних речовин і які підтримують структуру нервових волокон.
Мієлінова оболонка створюється мієлоцитами (шванівськими клітинами), які багаторазово огортають циліндр нервового волокна, зливаються і утворюють щільний жировий футляр. Мієлінова оболонка через проміжки рівної довжини переривається, залишаючи відкритими ділянки мембрани до 1 мкм, які назвали перехватами Ранв'є. Саме у цих ділянках відбувається зміна заряду на поверхні мієлінового волокна і через ці ділянки передається струм по волокну.
Безм'якотні нервові волокна не мають мієлінової оболонки, а віддалені одне від одного шванівськими клітинами.
Сукупність нервових волокон утворює нерви. З мієлінових волокон складаються чутливі, рухові нерви, нерви органів чуття і скелетних м'язів тощо. Безмієлінові нервові волокна належать, в основному, до симпатичної нервової системи.
Нерви зазвичай складаються як з м'якотних , так і безм'якотних волокон, але з різним їх співвідношенням. Наприклад, у блукаючому нерві, що іннервує всі органи грудної і частину органів черевної порожнини, кількість безм’якотних волокон сягає 80-95%. У нервах скелетних м'язів навпаки.
Властивості нервових волокон:
2. Двобічна провідність.
3. Цілісність, без якої неможливе проведення імпульсу. Цілісність порушуєть-ся при: перерізуванні нервового волокна (анатомічне порушення), знеболенні (фізіологічне порушення), перегріванні, переохолодженні нерва тощо.
4. Ізольоване проведення імпульсів (не переходить з одного волокна на інше).
5. Збудження проходить бездекрементно (без затухання).
6. Нервове волокно майже не стомлюється проводити імпульси.
7. Швидкість проведення по нервових волокнах залежить від діаметру волокна (чим більший діаметр, тим більша швидкість).
Проведення збудження по нервовому волокну
Проведення збудження вздовж нервових та м'язових волокон здійснюється за допомогою місцевих струмів, що виникають між збудженими та спокійними ділянками волокна. Напрямок місцевого струму такий, що він деполяризує сусідню з активною (А) спокійну (В) ділянку мембрани. Деполяризація швидко досягає критичного рівня та породжує ПД, який активує сусідню спокійну ділянку. Завдяки такому естафетному механізму збудження поширюється вздовж всього волокна.
Теорія проведення збудження за допомогою місцевого струму вперше була висунута Германом у 1899 р. На даний час вона отримала багато експерименталь-них підтверджень.
Фактори, від яких залежить швидкість проведення імпульсу по нервовому волокну:
1). Опір середовища навколо волокна . Якщо нервове волокно помістити у середовище без іонів (розчин сахарози), то проведення струму припиняється.
2). Внутрішній опір волокна.
3). Діаметр волокна. Чим більший діаметр волокна, тим менший опір воно чинить, а швидкість проведення збудження стає більшою.
Щоб заблокувати проведення нервового імпульсу (наприклад, для знеболення), необхідно або сильно підвищити величину порогу деполяризації нервового волокна або значно знизити амплітуду його ПД. Місцеві знеболюючі (новокаїн, кокаїн, дикаїн) викликають обидві ці зміни одночасно.

Утворення мієлінового нервового волокна
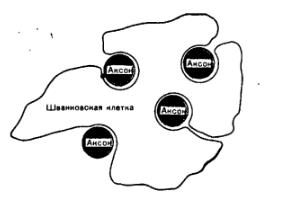
Ізоляція безмієлінових нервових волокон шванівською клітиною

А. Передача збудження по безмієліновому нервовому волокні.
Б. Передача збудження по мієліновому нервовому волокнні.
ІІ. Фізіологія скелетних м’язів
Фізіологічні властивості скелетних м’язів
М’язи бувають скелетні, гладенькі та м’яз міокарду.
Скелетні м’язи називають посмугованими. Вони є активною частиною опорно-рухового апарату і виконують наступні функції:
1) рух організму в просторі;
2) зміна положення частин тіла одних відносно інших;
3) підтримання пози і протидія гравітації;
4) вироблення тепла.
Скелетні м’язи хребтових мають наступні властивості: 1)збудливість; 2) провідність вздовж м’язових волокон; 3) скоротливість.
У природних умовах ці властивості викликаються нервовими імпульсами з нервових рухових (моторних) центрів, а у експерименті – за допомогою електростимулятора.
Збудливість м’язової тканини є меншою, ніж нервової. Тому прикладання електродів до м’язу безпосередньо його не збуджує, а діє через нервове закінчення. Щоб в експерименті отримати ефект чистого прямого подразнення, необхідно блокувати нервові закінчення отрутою кураре або електрод ввести безпосередньо у м’язове волокно.
Реєстрація електричної активності за допомогою внутрішньом’язового введення електродів і підсилювача називається електроміографія, а зареєстрована крива – електроміограма. Цей метод широко використовується у спортсменів, для оцінки стану опорно-рухового апарату, для діагностики ряду захворювань.
У залежності від умов м’язового скорочення воно буває 3-х типів:
1) ізотонічне: м’язове скорочення, при якому волокна вкорочуються, але напруга залишається постійною (скорочення без навантаження);
2) ізометричне: м’язове скорочення, при якому м’яз не може скоротитися (обидва кінці його закріпленні нерухомо), але напруга зростає (при спробі підняти важкий предмет);
3) змішане, яке зустрічається найчастіше.
Нервово-м’язова передача та збудливість нервових волокон
Передача імпульсу з нервового волокна на м’язове відбувається через нервово-м’язовий синапс, який складається з нервового закінчення, частини м’язового волокна і щілини між ними.
На кінці нервового волокна знаходиться потовщення, яке називається синапти-чною бляшкою, у якому розташовані пухирці (везікули) з біологічно активною речо-виною – медіатором ацетилхоліном і яке закінчується пресинаптичною мембраною. За пресинаптичною мембраною є синаптична щілина, а за нею – постсинаптична мембрана кінцевої пластинки м’язового волокна. На постсинаптичній мембрані знаходяться холінорецептори, що реагують на ацетилхолін (АХ), а також нікотин. Тому їх ще називають Н-холінорецептори. При з’єднанні рецепторів з ацетилхоліном в постсинаптичній мембрані відкриваються Na + , К + , Са 2+ канали.
Механізм передачі інформації з нервового на м’язове волокно
Внаслідок поширення збудження по нервовому волокну у пресинаптичній мем-брані відкриваються кальцієві канали, кальцій заходить у бляшку, а АХ виходить з пухирців у синаптичну щілину (СЩ). 1 ПД розкриває 300 пухирців. Далі ацетилхо- лін проходить через щілину, з’єднується з Н-холінорецепторами, внаслідок чого від криваються натрієві та калієві канали (натрій переходить у м’язове волокно, а калій – у щілину). Таким чином, кінцева пластинка деполяризується і розвивається ПД.
АХ виділяється у СЩ у великій кількості, коли рухові центри головного чи спинного мозку посилають збудження з метою здійснення рухової активності (скорочення) та у малій кількості для створення тонусу м’язів у спокої. Далі АХ руйнується ферментом ацетилхолінестеразою.
МПС м’язових волокон рівний приблизно (-90 мВ), на відміну від МПС нерво-вих волокон (-70 мВ). Один аксон іннервує багато волокон м’язів: у м’язах пальців рук – 10-25 м’язових волокон, у м’язах тулуба і кінцівок – декілька сотень, у камбалоподібному м’язі – 2000.
Будова м’язових волокон
М’яз складається з м’язових волокон, останні - з міофібрил, які є скоротливим апаратом м’язових волокон. У посмугованих м’язових волокнах міофібрили розділені на подібні за будовою ділянки – саркомери. У саркомерах впорядковано розміщені протофібрили, які являють собою подовжені полімеризовані молекули білків міозину та актину. Міозинові нитки удвічі товщі актинових. Тонкі довгі актинові нитки прикріплені до мембран саркомерів та входять кінцями між товсті міозинові нитки. Актинові та міозинові волокна з’єднані поперечними рухомими містками.
Якщо розглядати м’язові волокна під світовим мікроскопом, то можна побачити чергування темних і світлих смуг, через що скелетні м’язи назвали посмугованими. Темні широкі смуги були названі анізотропними, тобто А-дисками, так як вони утворені ділянками, де світлові промені проходять через актинові та міозинові нитки (відбувається подвійне променезаломлення). Світлі широкі смуги – ізотропні (І-диски), утворені одними актиновими нитками. Посередині А-диску -світла тонка смужка Н (ділянка одних міозинових волокон), а посередині І-диску – темна смужка Z, яка є тонкою мембраною, через пори якої проходять міофібрили і через яку фібрили не зміщуються всередині м’язового волокна.



При скороченні м’язового волокна нитки не вкорочуються, а ковзають одна по одній: актинові нитки входять у проміжки між міозиновими (І-диск зменшується, А-диск зберігає свій розмір, смужка Н теж зменшується).
Причиною ковзання є хімічна взаємодія між актином та міозином у присутності Са 2+ і АТФ. У стані спокою основна частина Са 2+ знаходиться у ендоплазматичній сітці м’язового волокна (замкнута система трубочок і цистерн, що оточують міофібрилу). Під час збудження потенціал дії поширюється по Т-трубках, стимулює вихід кальцію з сітки у цитоплазму м’язового волокна та проникнення його до міофібрил і створює умови для ковзання актинових і міозинових волокон.
Коли кальцієві помпи відкачують кальцій, настає розслаблення.
У відповідь на 1 подразнення виникає 1 скорочення м’язового волокна. Воно здійснюється у 3 стадії (фази):
1) латентна (прихована) – коротка;
Якщо в експерименті на м’яз діє два поодиноких подразнення, то вони сумуються (додаються) і амплітуда скорочення зростає. Якщо діють ритмічні подразнення з такою частотою, що їх ефекти додаються, настає сильне і тривале скорочення м’язу, що називається тетанічним скороченням або тетанусом (спазм м’язів).
При відносно малій частоті подразнень спостерігається зубчастий тетанус, при більшій частоті – гладкий суцільний тетанус.
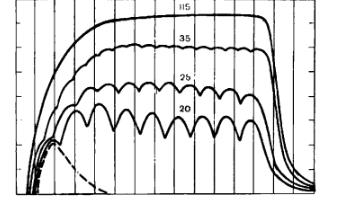
При частоті подразнення 20-25 Гц виникає зубчастий тетанус.
При частоті подразнення 35-45 Гц виникає гладкий (суцільний) тетанус.
Після припинення тетанічного подразнення волокна повністю розслабляються, а їх вихідна довжина відновлюється лише через деякий час (післятетанічна або залишкова контрактура).
Скелетні м’язи характеризуються такими властивостями, як розтяжність і еластичність. Здатність м’яза під впливом навантаження розтягуватись називається розтяжністю. Розтяжність м’язу з паралельними волокнами більша, ніж м’язу з піристою структурою. Якщо до м’язу підвісити тягар, він розтягнеться. Червоні м’язи розтягуються більшою мірою, ніж білі. Під еластичністю розуміють властивість розтягнутого м’язу повертатись до початкового стану після усунення впливу сили, яка зумовила деформацію. Після зняття тягару він набуде початкової довжини, але не зразу (залишкова деформація). Еластичність виражена тим сильніше, чим більша сила, яка його розтягує, і чим триваліша її дія.
Характерна ознака у нервовому волокні – здатність поширюватися. Передача нервового імпульсу полягає саме у проведенні ПД. Показником цього процесу вважають швидкість проведення. Швидкість проведення імпульсу нейроном значною мірою залежить від особливості його будови:
а) діаметра волокна;
б) наявності або відсутності мієлінової оболонки.
Характерна ознака проведення ПД будь-якими збудливими мембранами – бездекрементність: тобто величина ПД завжди однакова, навіть якщо він поширюється на відстань понад 1 м.
Проведення збудження в мієлінізованих нервах. Нервові відростки більшості соматичних нервів мієлінізовані. Лише дуже незначні їх ділянки, так звані перехоплення вузла (перехоплення Ранв’є), укриті звичайною клітинною мембраною. Такі нервові волокна характеризуються тим, що на мембрані лише в перехопленнях розміщені іонні канали. Крім того, ця оболонка підвищує опір мембрани. Тому при зрушенні мембранного потенціалу струм проходить через мембрану перехоплювальної ділянки, тобто стрибками (сальсаторно) від одного перехоплення до іншого. Унаслідок цього різко збільшується швидкість проведення нервового імпульсу.
Рефрактерність. Механізм ПД може поширюватися мембраною в обидва боки від місця свого первинного виникнення. Але у фізіологічних умовах ПД, що виникає, наприклад, у тілі нейрона, поширюється на периферію відростками лише в одному напрямку. Зворотного поширення ПД не відбувається через те, що впродовж деякого часу та ділянка мембрани, де розвинувся ПД, стає незбудливою. Цей стан незбудливості мембрани називають рефрактерністю. Незбудливість мембрани зумовлена тим, що після попереднього збудження натрієві канали на деякий час стають ін активованими. Рефрактерність – це минаючий період стану мембрани. Причому, якщо спочатку мембрана виявляється нечутливою до будь-якої сили подразника, що позначається абсолютним рефрактерним періодом, то потім настає відносний рефрактерний період, під час якогонад граничної сили подразник уже може спричинити ПД.
У людини розрізняють скелетні м'язи, серцевий м'яз (міокард)
та гладкі м'язи. Всі вони мають властивість збуджуватись, проводити це збудження та скорочуватись.
Скелетні м'язи підтримують тіло людини у певній позі, протидіють силам гравітації і беруть участь у реалізації різних локомоторних актів (ходьба, біг, стрибки, плавання тощо).
Морфо-функціональна структура. Скелетні м'язи складаються
з окремих волокон, які містять два види білка — актин та міозин. Міозинові нитки вдвічі товщі, ніж актинові. У стані спокою м’яза тонкі довгі актинові нитки входять у проміжки між товщими та коротшими міозиновими нитками. Ізотропні диски складаються лише з актинових протофібрил, а анізотропні - з актинових та міозинових.
Світла центральна зона (Н) не має актинових ниток. Пластинка Z лежить посередині диска I, скріплюючи між собою ці нитки.
Ділянки міофібрил між пластинками називаються саркомерами.
Пучок ниток міозину в середині саркомера має вигляд темної зони
(А-диск). По обидва боки від нього розташовані світлі смуги
(диски I). Завдяки такому чергуванню світлих та темних смуг волокна скелетного та серцевого м'язів мають поперечносмугастий
вигляд. Міозинові та актинові нитки сполучені між собою містками, що мають головку та шийку. Ці містки розташовані в кілька
рядів уздовж міозинових протофібрил.
На підставі фізіологічних, біохімічних та структурних особливостей виділяють три типи м'язових волокон - швидкі (білі), повільні (червоні) та проміжні. Їх характеристика наведена в табл. 2. Співвідношення цих типів волокон у різних м'язах неоднакове. В одних м'язах переважають швидкі, в інших - повільні волокна. Це обумовлює різний характер скорочення: одні м'язи скорочуються дуже швидко, але короткий час, інші - повільно, але довго і без ознак утоми.
Таблиця 2. Характеристика швидких та повільних м’язових волокон
| Характерні ознаки | Швидкі | Повільні |
| Колір | Білий | Червоний |
| Розмір | Більші | Менші |
| Вміст глікогену | Багато | Мало |
| Кількість мітохондрій | Мало | Багато |
| Кількість міоглобіну | Мало | Багато |
| Кровопостачання | Невелике | Добре |
| Характер обміну | Анаеробний | Аеробний |
| Збудливість | Висока | Нижча |
| Скорочення | Швидке | Повільне |
| Стомлюються | Раніше | Пізніше |
Енергетичні процеси істотно відрізняються. Для скорочення
волокон потрібна АТФ, але її вистачає для скорочення лише протягом 1-2 с. За рахунок фосфокреатину, який бере участь у ресинтезі АТФ, період скорочення триває довше – 7-8 с, а за рахунок анаеробного гліколізу-до 1 хв.
Для тривалішого скорочення м'язів потрібен кисень, оскільки
відбувається окислення вуглеводів, ліпідів та білків і за рахунок
окисного фосфорилювання ресинтезуеться потрібна кількість АТФ.
У різних людей співвідношення між повільними та швидкими волокнами в одних і тих же м'язах може бути неоднаковим. Тому одні люди здатні долати короткі, спринтерські дистанції, а інші - довгі, стаєрські.
Рухова одиниця.Аксони мотонейронів спинного чи довгастого мозку іннервують різну кількість м'язових волокон. Поодиноке нервове волокно мотонейрона і м'язові волокна, що ним іннервуються, називаються нейромоторною одиницею або руховою одиницею (РО). Що точніші рухи виконує м'яз, то менше співвідношення між нейроном і м'язовими волокнами. Наприклад, мотонейрони, що іннервують м'язи гортані чи ока, закінчуються біля 2—3 волокон (співвідношення 1:2 або 1:3), а мотонейрони, що іннервують литковий м'яз, іннервують кілька сотень м'язових волокон. З функціональної точки зору розрізняють швидкі і повільні РО.
Типи скорочень залежно від режиму навантаження.
Розрізняють: ізометричн1, ізотонічні, ексцентричні та змішані скорочення.
l Ізометричні скорочення відбуваються при незмінній довжині м'яза (наприклад, при спробі підняти дуже важкий предмет). У експерименті цей вид скорочень можна одержати, закріпивши м'яз з обох боків та подразнюючи його.
l Ізотонічне скорочення — це скорочення без навантаження, скорочення м’язів з укороченням за збереження постійної напруги.
l Ексцентричне, коли м’яз подовжується.
В організмі людини м'язи рідко
скорочуються в ізотонічному або ізометричному режимі. Зазвичай більшість природних скорочень м’язів змішані, тобто анізотонічного типу, коли м’яз стає коротшим уразі підвищення напруги.
Типи скорочень залежно від частоти подразнення. У відповідь
на одиночне порогове подразнення виникає одиночне скорочення
м'яза, що складається з трьох періодів: латентного, скорочення і розслаблення. Латентний період лежить між моментом подразнення і початком скорочення. У цей час відбуваються в м'язі біохімічні та біофізичні процеси, одним із проявів яких є ПД. Таким чином, ПД з'являється раніше, ніж скорочується м'яз. Період скорочення триває від його початку до
максимуму, а період розслаблення — від максимуму до
початкового рівня. Тривалість одиночного скорочення неоднакова
у різних м'язів людини. Наприклад, у м'яза, який рухає очне яблуко, вона становить 0,01 с, у гомілкового — 0,05 с, у камбаловидного — 0,1 с. У різних тварин тривалість скорочення м'язів неоднакова. Крім того, вона залежить від температури, ступеня стомлення м'яза тощо.
Одиночні скорочення можуть додаватись, і тоді розвиваються тетанічні скорочення — тривалі скорочення м'яза.
Розрізняють зубчастий та суцільний тетанус. 3убчастий
тетанус буває тоді, коли наступне подразнення діє в період
розслаблення, а суцільний — коли наступне подразнення надходить у період скорочення. Амплітуда скорочень під час тетануса більша, ніж при одиночних скороченнях.
У природних умовах спостерігаються асинхронні скорочення
м'язових волокон. Це означає, що імпульси від різних мотонейронів досягають своїх м'язових волокон неодночасно, спочатку скорочується одна їх група, потім друга, потім третя і т.д. А перша група у цей час вже встигає розслабитись. Таким чином, хоча м'язові волокна скорочуються неодночасно, м'яз весь час перебуває у стані скорочення. Сила скорочень залежить від числа волокон, що одночасно скорочуються.
Зв'язок між збудженням та скороченням м'яза. ПД м'язового
волокна поширюються в обидва боки від нервово-м'язового синапсу. Важливу роль відіграють поперечні трубки мембрани волокна
(Т-трубки), що становлять собою заглиблення мембрани. Мембрана цих трубок теж має потенціалзалежні натрієві канали. Поряд з Т-трубками у волокні розташовані цистерни ендоплазматичної сітки, в яких депонується Са 2 +.
Під час збудження ПД поширюється в глиб Т-трубок, зумовлюючи вих1д Са 2 + із цистерн. Концентрація Са 2 + у волокні значно підвищується, і він дифундує до міофібрил. Са 2+ — це основна ланка зв'язку між збудженням та скороченням м'яза.
Механізм скорочення. ПД м'язового волокна — початковий
етап, потрібний для скорочення м'яза. Скорочення пояснюється теорією ковзання. Спостерігається взаємне переміщення (ковзання) товстих міозинових та тонких актинових протофібрил при незмінній їх довжині. Актинові протофібрили втягуються у проміжки між міозиновими. Причиною цього є рух головок поперечних актоміозинових містків. Кожен місток то приєднується, то від'єднуєтьея від сусідньої протофібрили.
У стані спокою місток не може приєднатись до актину, бо між
ними містяться білки тропонін та тропоміозин, які блокують місце приєднання. При підвищенн1 концентрації іонів Са 2 +
(у присутності АТФ) тропінін змінює свою конфігурацію і відсовує молекулу тропоміозину, створюючи умови для з'єднання головки містка з актином. Це супроводжується зміною положення головки і переміщенням нитки актину з наступним розриванням містка. Амплітуда кожного такого переміщення становить близько 20 нм, а частота — 5—50 за 1 с.
Розслаблення волокна вібувається внаслідок діяльності Са 2+ -
насоса, який накачує ці іони назад у цистерни саркоплазматичної
сітки. Концентрація Са 2+ у міоплазмі знижується, і тропоміозин знову блокує активні ділянки актину. Важливе значення для розслаблення м'яза має АТФ: зменшення її концентрації призводить до утворення постійного зв'язку між поперечними містками і актиновими протофібрилами, тобто настає контрактура м'яза. Таке явище спостерігається і при трупному закляканні.
На скорочення чи напруження скелетних м'язів впливає ряд
факторів: кількість та вид волокон, що беруть участь у скороченні,
частота ПД волокон, функціональний стан м'яза (рівень збудливості, вміст АТФ, вуглеводів, ліпідів, іонів Са 2 +, напруга кисню,
кількість діючих поперечних містків тощо), ступінь втоми м'яза.
На функціональний стан м'яза істотно впливає його кровопостачання.
Еластичність і розтяжність скелетного м'яза. Під еластичністю
розуміють властивість деформованого тіла повертатись до початкового стану після усунення впливу сили, яка зумовила деформацію. Якщо до м'яза підвісити тягар, він розтягнеться. Після зняття цього тягаря він набуде початкової довжини. Здатність м'яза під впливом навантаження розтягуватись називається розтяжністю, величина якої залежить від маси тягара, часу його дії, в'язкості і пластичності м'яза. Розтяжність м'яза з паралельними волокнами більша, ніж м'яза із перистою структурою. Червоні м'язи розтягуються більше ніж білі.
При скороченні м'яза хімічна енергія перетворюється на теплову і механічну. При скороченні м'яза виділяється тепло. Розрізняють дві фази тепло продукції — початкову (під час скорочення) і затримане теплотворення. Початкова фаза залежить від хімічних процесів, які
переводять м'яз із стану спокою в активний стан. Відбувається
вона в анаеробних умовах. Друга фаза пов'язана з процесами, які
забезпечують ресинтез АТФ (переважно гліколіз та окисне фосфорилювання). Тривала активність м'язів (протягом кількох годин)
забезпечується за рахунок окислення ліпідів.
Абсолютна сила — це відношення максимальної сили до
площі поперечного перерізу м'яза. Ця величина дає змогу порівняти силу різних м'язів одного організму і силу м'язів різних тварин.
Наприклад, абсолютна сила литкового м'яза людини становить
5,9 кг/см 2 , жувального—10 кг/см 2 , триголового м'яза плеча-
16,8 кг/см 2 . При тренуванні м'язів, яке супроводжується їх робочою гіпертрофією, сила м'яза значно зростає.
Робота м'язів може бути динамічною та статичною. Динамічна робота визначається так: величину піднятого вантажу множать на показник скорочення м'яза. Вимірюється ця робота в кілограмометрах, джоулях та калоріях. Максимальною робота буває при середніх навантаженнях. При надто великому або замалому вантажі величина роботи зменшується. Робота, при якій м'язи майже не скорочуються (ізометричні скорочення), називається статичною. Прикладом її може бути робота при утриманні
(але не переміщенні) вантажу в певному положенні. Статична робота вимірюється величиною вантажу, помноженою на час його
утримання. При статичній роботі втома розвивається швидше, ніж при динамічній.
МЕХАНІЗМИ ПРОВЕДЕННЯ ЗБУДЖЕННЯ (ПД) ПО НЕРВОВОМУ І М′ЯЗОВОМУ ВОЛОКНУ
Механізм поширення ПД вздовж мембрани
Основу поширення ПД по нервових і м’язових волокнах складають локальні електричні струми, які виникають між деполяризованою ділянкою волокна і неполяризованими (спокійними) ділянками мембрани.
Механізм поширення ПД по нервовому волокну. Стрілками показані локальні струми.
Локальний струм, що виникає, має вихідний напрямок у ″ спокійній″ ділянці волокна, це означає, що він спричиняє деполяризацію мембрани. Остання виявляє себе розвитком КЕТП і локальної відповіді. Якщо сумарна деполяризація ( КЕТП + локальна відповідь) досягає критичного рівня, то виникає ПД. А потім все спочатку: локальний струм з’являється між деполяризованою ділянкою волокна, де щойно виник ПД, і наступною - поблизу розміщеною “спокійною” ділянкою. Таким чином, імпульс (ПД) поширюється вздовж волокна.
Деполяризація “спокійної” мембрани допорогової величини виникає на відстані 1-3 мм від місця появи ПД.
Фактори, що визначають швидкість проведення ПД
Фактори, які мають значення для проведення збудження по нервових та м’язових волокнах, можна поділити на дві групи: фізичні та фізіологічні.
І Фактори фізичні:
а) опір зовнішнього середовища, яке оточує волокно.
Якщо ділянку нервового волокна помістити у середовище, в якому немає іонів, тобто середовище, яке має великий опір (наприклад, розчин сахарози), то проведення збудження через цю ділянку повністю припиняється. Воно одразу ж відновлюється, якщо дві розділених неелектролітом ділянки волокна з’єднати металевим провідником;
б) опір внутрішнього середовища волокна. Він визначається опором аксоплазми на одиницю довжини (поздовжній опір). Цей опір залежить від площі поперечного перетину, а отже, від діаметра волокна. Що більший діаметр волокна, то менший поздовжній опір і вища швидкість проведення ПД. Протилежна картина спостерігається при зменшенні діаметра волокна;
в) пасивні електричні параметри мембрани волокна – ємність та опір.
У мієлінізованих нервових волокнах мієлін є чудовим ізолятором, тому опір мембрани дуже високий і ПД не може поширюватися через покриті мієліном ділянки волокна.
ІІ Фактори фізіологічні:
а) амплітуда ПД (Епд), тобто сила імпульсів збудження. Чим вище Епд, тим на більшу відстань від ділянки деполяризації можуть поширюватися локальні струми.
Епд визначається силою вхідного натрієвого струму, який, у свою чергу, залежить від концентрації Na + у позаклітинному середовищі, кількості одночасно відкритих натрієвих каналів, швидкості наростання інактивації натрієвих каналів і активації калієвих;
б) збудливість волокна. Інтегральним показником цієї властивості, як уже зазначалося, є поріг деполяризації (ΔV). Усі фактори, які збільшують ΔV, зменшують швидкість проведення імпульсів аж до повної блокади.
Для характеристики фізіологічних факторів, що визначають швидкість поширення ПД по нервових волокнах, було введено показник фактор надійності (ФН).

Чим вищий ФН, тим більша швидкість проведення імпульсів і навпаки. У нормі для нервових волокон ФН = 5-6. Якщо ФН становить менше 1, то проведення ПД через таку ділянку повністю припиняється (блокада проведення). Із наведеної формули видно, що ФН зменшується, якщо Епд зменшується або ΔV збільшується.
Закони проведення імпульсів по нервових і м’язових волокнах
Проведення імпульсів по волокнах підпорядковується таким законам.
1 Анатомічна і фізіологічна неперервність волокна. Є обов’язковою умовою проведення збудження. Порушення анатомічної неперервності виникає при перетині нервів або їх травмі. Під порушенням фізіологічної неперервності розуміють зміни функціональних характеристик нервового волокна, які визначають проведення імпульсів – зменшення фактора надійності (наприклад пересихання нерва).
2 Двобічне проведення. ПД поширюється в обидва боки від місця виникнення збудження, а також по розгалуженнях нервових волокон. Якщо подразнювати електричним струмом ділянку нерва, то імпульси поширюються як в аферентному, так і в еферентному напрямках.
3 Ізольоване проведення. У периферичному нерві імпульси поширюються по кожному волокну ізольовано, тобто не переходять з одного волокна на паралельне, поряд розміщене. Внаслідок цього імпульси здійснюють вплив тільки на ті клітини, з якими контактують закінчення даного нервового волокна. Ця обставина має важливе значення у зв’язку з тим, що кожний периферичний нерв містить велику кількість нервових волокон – рухових, чутливих, вегетативних, - які іннервують різні за структурою і функціями клітини та тканини.
4 Бездекрементне проведення, тобто проведення без затухання, при якому ПД не змінює своїх характеристик у процесі поширення по волокну.
5 Проведення імпульсів не спричиняє втомлення волокон – відносна невтомлюваність нерва.
Енергетичне забезпечення проведення імпульсів по нервах
Під час ПД іони Na + входять у клітину, а К + виходить з неї. Це повинно спричиняти збільшення внутрішньоклітинної концентрації Na + і зменшення К + , тобто зменшення градієнта концентрації названих іонів. Але під час поодиноких ПД переміщення іонів через мембрану настільки малі, що зміни їх концентрації у клітині несуттєві – їх навіть не можна зареєструвати.
Інша справа, коли мова йде про проведення імпульсів по нервовому волокну, по якому проходить від 100 тис. до 50 млн імпульсів за 1 годину. У цей час градієнти концентрацій Na + і К + можуть суттєво зменшуватися. Для того щоб цього не відбувалося, необхідно відновлювати названі градієнти, для чого служать Na + - К + - насоси, робота яких потребує енергії. У цілому ж валові енерговитрати нервового волокна на роботу Na + - К + - насосів порівняно невеликі. Так, 1 г нерва у жаби виділяє при максимальному подразненні тільки на 20-100% більше тепла, ніж у стані спокою. Це набагато менше, якщо порівнювати зі збудженням м’язів.
Даною обставиною, власне, і пояснюється відносна невтомлюваність нерва.
Особливості проведення збудження по мієлінізованих нервових волокнах
Як відомо, нерв у своєму складі містить мієлінізовані (товсті) та немієлінізовані (тонкі) нервові волокна. У нервах, середніх за величиною, перших приблизно у 2 рази менше, ніж других. Мієлінізовані волокна мають мієлінову оболонку, утворену шванівськими клітинами. Вона являє собою багато разів закручену навколо аксона плазматичну мембрану шванівських клітин. Її головним компонентом є сфінгомієлін, який має властивості ізолятора: він зменшує іонний струм через мембрану у 5 тис. разів і зменшує ємність мембрани у 50 разів.
Між двома поблизу розміщеними клітинами Шванна є так звані перехвати Ранв’є – ділянки нерва, невкриті мієліновою оболонкою, довжина яких становить 2-3 мкм. Це єдина ділянка мієлінізованого волокна, яка здатна проводити через себе іони. У перехватах Ранв’є щільність натрієвих каналів на одиницю площі у 100 разів вища, ніж у мембрані немієлінізованих нервових волокон.
У зв’язку з великим електричним опором мієліну ПД не проводиться через мієлінізовану ділянку, а може проходити тільки через перехвати Ранв’є, в яких немає мієліну. ПД нібито перестрибує від одного перехвату Ранв’є на інший (рис.7.2). Таке проведення імпульсів має назву сальтаторного.
Сальтаторне проведення імпульсів по мієлінізованих нервових волокнах.
Сальтаторне проведення – це проведення імпульсів у мієлінізованих нервових волокнах від одного перехвату Ранв’є до іншого без затримки проведення на покритих мієлінових ділянках волокна. Завдяки сальтаторному проведенню збільшується швидкість поширення імпульсів від 5 до 50 раз. Оскільки деполяризація виникає тільки на ділянках перехвату Ранв’є, то тільки тут мають місце переміщення іонів. Воно у сотні раз менші, ніж в немієлінізованих волокнах. Тому у волокнах, покритих мієліном, значно менші енерговитрати на роботу Na + - К + - насосів, а отже, значно менша втомлюваність порівняно з немієлінізованими волокнами.
Швидкість проведення імпульсів по мієлінізованих волокнах залежить від факторів, про які мова йшла вище. Крім того, на цей параметр впливає довжина мієлінізованих ділянок, тобто відстані між поруч розміщеними перехватами Ранв’є. Що більші ці відстані, то вища швидкість проведення імпульсів.
Сумарний ПД нервового стовбура
Якщо відводити ПД від окремого нервового волокна, то амплітуда й інші характеристики ПД не залежать від сили подразнення – закон „все або нічого”. Якщо ж електричні потенціали відводити від нерва, який складається з великої кількості нервових волокон, то у цьому випадку виявляє себе закон силових відносин: при збільшенні сили подразника (електричного струму) амплітуда потенціалів, що реєструються, також збільшується, але до деякої максимальної величини, а потім залишається постійною незалежно від подальшого збільшення сили подразнення.
Це пояснюється тим, що електрична відповідь цілого нерва є сумою відповідей окремих його волокон, з’єднаних паралельно. Пороги подразнення окремих волокон відрізняються один від одного. При малій силі стимулу збудження виникає у найбільш збудливих нервових волокнах. Збільшення стимулу веде до збільшення кількості збуджених волокон. Тому сумарна відповідь на подразнення збільшується доти, доки усі волокна не будуть залучені до реакції.
Якщо електроди, за допомогою яких подразнюють і реєструють сумарний потенціал від нерва, розмістити на достатньо великій відстані один від одного (10-20 см), то можна виявити декілька „піків” сумарного потенціалу.
Реєстрація сумарного потенціалу дії нервового стовбура.
Поява хвиль А, В, С пов’язана з тим, що швидкість проведення імпульсів в окремих нервових волокнах різна (див.нижче).
У клініці широко використовують реєстрацію сумарних ПД великих стовбурів і скелетних м’язів в умовах їхньої природної активності в організмі. Ці методи отримали назву електронейрографії і електроміографії.
Класифікація нервових волокон
Найбільшого поширення дістали дві класифікації :
Класифікація волокон за Ерлангером і Гасером

Класифікація сенсорних нервових волокон за Ллойдом і Хантом

Основні причини і механізми порушень проведення імпульсів по нервових волокнах
І Порушення анатомічної неперервності: перетин нерва, його травма.
ІІ Порушення фізіологічної неперервності: зміна фізіологічних характеристик нервових волокон, що визначають проведення імпульсів (зменшення фактора надійності).
В основі порушень фізіологічної неперервності можуть лежати такі механізми:
1 Зменшення амплітуди ПД. Це може відбуватися,
- зменшенні позаклітинної концентрації іонів Na + ;
- дії блокаторів натрієвих каналів (тетродотоксину);
- дії місцевих анестетиків (речовини, які уповільнюють відкриття активаційних воріт натрієвих каналів і прискорюють інактивацію натрієвих і відкриття калієвих.
2 Зменшення збудливості нервових волокон.
Причиною цього можуть бути :
- гіперкаліємія;
- локальне збільшення позаклітинної концентрації К + у тканинах при ушкодженні клітини;
- гіпокаліємія;
- гіперкальціємія;
- ушкодження мембрани;
- дія місцевих анестетиків (зміщують критичний рівень деполяризації за рахунок збільшення кількості інактивованих натрієвих каналів.
ІІІ Порушення енергетичного забезпечення нервових волокон.
Ці порушення є найбільш поширеною причиною розладів провідності нервових волокон. Гіпоксія, голодування, гіповітамінози, токсини, отрути, порушуючи енергетичний обмін, призводять до розладів роботи Na+-K+-насосів. Це спричиняє стійку деполяризацію мембрани і, як наслідок, зменшення збудливості нервових волокон.
IV Демієлінізація нервових волокон.
Руйнування мієліну і порушення його утворення є причинами зменшення швидкості проведення імпульсів. Найбільш поширеними причинами цих змін є:
- порушення діяльності і загибель клітин Шванна (спадкові і набуті дефекти);
- порушення біохімічних процесів, що забезпечують синтез хімічних компонентів мієліну (наприклад, при діабетичних нейропатіях);
- руйнування мієліну внаслідок аутоімунних механізмів (наприклад, при аутоалергічних хворобах).
Читайте также: