
Регистрация пд нервного волокна
При раздражении нерва пороговым и сверхпороговым значениями электрического тока отводящие электроды, размещенные на его поверхности, регистрируют суммарный ПД. Этот ПД является производным изменения разности потенциалов между отводящими электродами, обусловленной динамикой ионных токов, протекающих от невозбужденных к возбужденным участкам нервных волокон.
О с н а щ е н и е: двухлучевой осциллограф, усилитель биопотенциалов, влажная камера для нерва с двумя отводящими, заземляющим и двумя раздражающими электродами, электронный стимулятор, набор инструментов, раствор Рингера для холоднокровных животных. Объект исследования - седалищный нерв лягушки.
С о д е р ж а н и е р а б о ты. Выделите у лягушки седалищный нерв от позвоночника до коленного сустава. Поместите его на раздражающие, заземляющий и отводящие электроды влажной камеры (рис. 5.1 из руководства к практич. занятиям ). Закройте влажную камеру крышкой, края которой предварительно обработайте вазелином. Соедините отводящие электроды с усилителем биопотенциалов и осциллографом, а раздражающие электроды со стимулятором. Установите выходные параметры стимулятора: режим раздражения - одиночная серия, число импульсов в серии - 3-4, длительность раздражающих импульсов - 0,1-0,2 мс, частота - 100-150 Гц. Подберите сначала пороговый, а затем максимальный токи.
При нанесении на нерв импульсов максимального тока на экране осциллографа можно увидеть высокоамплитудные двухфазные ПД. Наличие двух фаз в составе ПД нервного ствола объясняется тем, что возбуждение последовательно проходит под 1-м и 2-м отводящими электродами. В момент появления волны возбуждения в зоне 1-го электрода фиксируется разность потенциалов между возбужденным (1) и невозбужденным (2) участками нерва - формируется первая фаза ПД. В дальнейшем ситуация изменяется. Волна возбуждения уходит из зоны l-го электрода, перемещаясь в зону 2-го электрода. В этом случае вновь фиксируется разность потенциалов, но теперь уже между невозбужденным (1) и возбужденным (2) участками нерва - в дополнение к первой в составе ПД формируется вторая фаза (рис. 5.2 из руководства к практич. занятиям ).
На данном этапе опыта можно доказать закон двустороннего проведения возбуждения по нерву. для этого снимите нерв с электродов и уложите его на электроды вторично, но так, чтобы конец, ранее находившийся в зоне отводящих электродов, теперь оказался бы в зоне раздражающих электродов. Включите стимулятор; вновь наблюдаются ПД.
Преобразуйте регистрируемые двухфазные ПД в монофазные. Для этого повредите нерв под отводящим электродом 2, используя пинцет или зажим (рис. 5.3 из руководства к практич. занятиям ). В этом случае возбуждение перестает распространяться в зону электрода 2 - потенциал электрода 2 становится неизменным. Последнее приводит к тому, что ПД, регистрируемый в зоне активного электрода 1 относительно индифферентного электрода 2, превращается из двухфазного в монофазный (рис. 5.4 из руководства к практич. занятиям ).
Зарегистрируйте монофазные ПД при нанесении на нерв пороговых (а), субмаксимальных (б) и максимальных (в) электрических импульсов тока (см. рис. 5.4 из руководства к практич. занятиям ). Отметьте, что в диапазоне пороговых, субмаксимальных и максимальных воздействий амплитуда ПД нерва изменяется в соответствии с законом "силы". Это связано с тем, что целый нерв состоит из множества волокон, каждое из которых обладает различной возбудимостью. Следовательно, при пороговых и сверхпороговых воздействиях различия амплитуды ПД объясняются неодинаковым числом возбужденных волокон в составе целого нерва.
Смочите небольшой ватный тампон 2 % раствором новокаина. Наложите его на участок нерва, расположенный в зоне заземляющего электрода. Спустя 5-7 мин после наложения отметьте характерные изменения амплитудно-временных характеристик регистрируемых ПД (рис. 5.5 из руководства к практич. занятиям ). Спустя 7-10 мин ПД исчезают. Угнетение генерации ПД под влиянием новокаина объясняется тем, что он, вытесняя Са из участков его связывания на сенсоре напряжения активационного воротного механизма Nа-каналов, делает потенциалзависимые Nа-каналы нечувствительными к изменениям мембранного потенциала. В результате нервные волокна теряют и возбудимость, и проводимость.
Оформление протокола. 1. Зарисуйте схему отведения двухфазных и монофазных ПД, зарисуйте двухфазные и монофазные ПД.
2. Используя калибровочные сигналы осциллографа, определите длительность ПД, амплитуду пороговых, субмаксимальных и максимальных ПД.
3. Объясните биофизический механизм возникновения двухфазных и монофазных ПД при расположении отводящих электродов на поверхности нерва.
4. Зарисуйте изменения ПД нерва после наложения на него ватного тампона, смоченного новокаином.
5. Сделайте вывод о возможности использования новокаина в практической медицине.
В зависимости от расположения электродов на объекте различают два метода регистрации ПД — биполярный (двухполюсный) и униполярный (однополюсный). Внутриклеточная регистрация ПД осуществляется только униполярным методом.
Биполярным методом регистрации электрических потенциалов называют такой, при котором оба отводных электрода расположены на участках объекта исследования, электрические потенциалы которых изменяются во времени. В случае пользования униполярным методом регистрации один отводный электрод (активный) расположен на участке объекта исследования, электрический потенциал которого изменяется во времени, а второй (пассивный) — на участке этого же объекта, электрический потенциал которого является постоянным (например, нулевым). Рассмотрим суть методов биполярной и униполярной регистрации ПД на примере его отведения от изолированного нерва.
В случае биполярной регистрации ПД нерва оба отводных электрода располагают на его поверхности таким образом, чтобы волна ПД (волна возбуждения), которая распространяется вдоль нервных волокон, могла пройти сначала под одним, а затем под вторым электродом. В результате этого кривая ПД, который отводится биполярным методом, будет формироваться как алгебраическая сумма двух, будто разных ПД, а, следовательно, должна иметь двухфазную форму.
Конструкцией катодного осциллографа предусмотрено, что отводные электроды можно условно считать разными полюсами (один, что соединенный на входе усилителя с землей, является негативным, а второй — позитивным полюсом). Потому обе фазы двухфазного ПД всегда регистрируются разнонаправленными: при прохождении электроотрицательной волны возбуждения под негативным отводным электродом луч осциллографа отклоняется вверх от изолинии, то есть в позитивную часть его шкалы. При прохождении же той же волны под позитивным отводным электродом луч осциллографа отклоняется вниз от изолинии.
В случае униполярной регистрации ПД нерва отводные электроды размещают на его поверхности таким образом, что ПД проходит только под одним электродом („активным”) и не может достичь того участка нерва, на котором помещен второй электрод („пассивный”). Кривая ПД, зарегистрированного униполярным методом, всегда имеет однофазную форму.
Если нужно униполярное отведение ПД от изолированного нерва, то можно превратить один из отводных электродов в пассивный, повредив участок нерва, расположенный между отводными электродами, тем самым, лишив ее возможности проводить возбуждение.
Цель работы: обнаружить форму кривой ПД, зарегистрированного биполярным методом.
Для работы необходимо: установка для регистрации ПД (катодный осциллограф), усилитель, электростимулятор, раздражающие и отводные электроды, влажная камера, вазелиновое масло, изолированный седалищный нерв лягушки, концентрированный (9,5%) раствор аммиака.
Ход работы
1. Подготовительный этап работы.
Изолированный седалищный нерв лягушки помещают во влажную камеру, и присоединяют к нему раздражающие и отводные электроды (на расстоянии 3-5 см). Заливают нерв вазелиновым маслом. Переводят стимулятор на работу в режиме „ритмическая серия”, а осциллограф — „ожидание на развертывание” с запуском от синхронизирующего импульса стимулятора. Включают приборы и усилитель в сеть.
Подавая на нерв ритмичную серию электрических импульсов (с частотой 10—30 имп/с), увеличивают их силу к моменту появления на экране осциллографа изображения ПД нерва достаточно большой амплитуды. Подбирают на осциллографе желаемую скорость развертывания, после чего прекращают стимуляцию нерва.
2.Основний этап работы.
Возобновить стимуляцию нерва, не изменяя ранее подобранных значений характеристик электрических импульсов.
Отметить, какую форму имеет на экране осциллографа кривая ПД, которая отводится от нерва. Умертвить с помощью 9,5% раствора аммиака участок нерва, расположенный между отводными электродами. Отметить, как изменилась на экране осциллографа форма кривой ПД.
Рекомендации относительно оформления результатов работы: в протоколе исследования изобразить кривые ПД, которые наблюдались на экране осциллографа при разных способах расположения на нерве отводных электродов.
В выводах указать: при каком способе расположения отводных электродов на нерве кривая ПД имеет двухфазную форму, а при каком — однофазную форму; какой метод регистрации ПД называют биполярным, а какой — униполярным.
Литература
Основная
1. Физиология человека (учебник для медвузов). Покровский В.М., Коротько Г.Ф., ред. М.: Медицина, 2003, 2005.
2. Физиология. Под ред. В.Г.Шевчука. Пособие по физиологии.- Винница: Новая книга. 2005. – 576 с. (русский перевод практических работ кафедрой).
3. Методические указания для самостоятельной работы студентов. Утверждены на методическом заседании кафедры. 2006
Дополнительная
- Textbook of medical physiology / Arthur C. Guyton, John E. Hall. 2000, 2006.
Физиология человека: в 3-х томах. Перевод с английского. Под ред. Р.Шмидта и Г.Тевса. – М: Мир, 1996
Потенциал действия нервной клетки
- Потенциал действия нервной клетки
- Ионные механизмы потенциала действия
- Проведение возбуждения по нервным волокнам
- Законы проведения возбуждения
- Типы нервных волокон и их функции
- Все страницы
Проведение ПД по мембране можно сравнить с поджиганием пороховой дорожки: вспыхнувший порох немедленно воспламеняет впереди лежащие частицы, и пламя движется вперёд до конца дорожки.
Продолжительность потенциала действия нервной клетки измеряется единицами миллисекунд (мс).
Потенциалы действия, зарегистрированные двумя электродами, один из которых находится внутри клетки, а другой — в окружающем растворе, представлены на рис. 5-3 и 5-7.
Рис. 5–3. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке –80 мВ — исходный уровень МП.
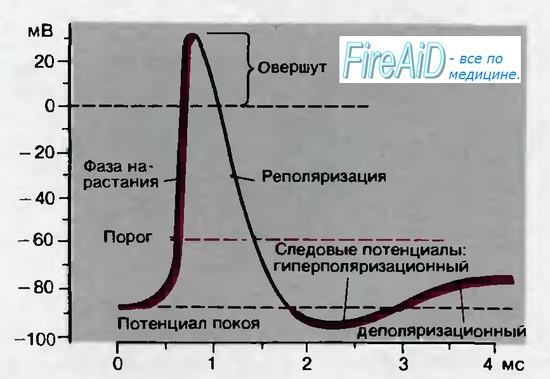
Между моментом нанесения раздражения и первым проявлением ПД имеется задержка — латентный период. Латентный период соответствует времени, когда ПД движется по мембране нервной клетки от места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны — локальный ответ. При достижении критического уровня деполяризации, который в среднем составляет —55 мВ, начинается фаза деполяризации. В эту фазу уровень МП падает до нуля и даже приобретает положительное значение (овершут), а затем возвращается к исходному уровню (фазареполяризации). Фазы деполяризации, овершута и реполяризации образуют спайк (пик) ПД. Длительность спайка составляет 1—2 мс. После спайка наблюдается замедление скорости спада потенциала — (раза следовой деполяризации. После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации. Эти следовые потенциалы могут длиться десятки и сотни миллисекунд.
В основе изменений мембранного потенциала (МП), происходящих в течение потенциала действия (ПД), лежат ионные механизмы. На рис. 5—7 представлены суммарные ионные токи, протекающие через мембрану нервной клетки в ходе потенциала действия.
Рис. 5-7. Потенциал действия и ионные токи нервной клетки [5|. По оси ординат отложены значения МП (мВ), по оси абсцисс — время (мс)
Локальный потенциал. Раздражение клетки приводит к открытию части Na+-каналов и появлению локального (нераспространяющегося) потенциала.
Фаза деполяризации. При достижении критического уровня деполяризации мембраны (соответствует порогу активации потен-циалозависимых Na+-каналов) начинается лавинообразный процесс открытия большого количества Na+-каналов. В фазу деполяризации происходит массивный вход в клетку ионов Na+ по концентрационному и электрохимическому градиентам.
Овершут. Деполяризация мембраны приводит к реверсии МП (МП становится положительным). В фазу овершута Na+-ток начинает стремительно спадать, что связано с инактивацией потенциало-зависимых Na+-каналов (время открытого состояния –– доли миллисекунды) и исчезновением электрохимического градиента Na+.
Фаза реполяризации. Помимо инактивации Na+-каналов, развитию реполяризации способствует открытие потенциалозависимых K+-каналов. Этот процесс происходит медленнее, чем открытие Na+-каналов, но K+-каналы остаются открытыми более продолжительное время. Выход К+ наружу способствует полному завершению фазы реполяризации.
Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов. Восстановление исходного уровня МП приводит Na+- и K+-каналы в состояние покоя.
В ходе развития ПД происходят изменения возбудимости мембраны нервной клетки.
Абсолютно рефрактерный период. Во время фазы деполяризации и большей части фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течение которого даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинства Na+-каналов.
Относительно рефрактерный период. В конце фазы реполяризации, а также во время следовой гиперполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители. Это связано со значительным реполяризующим действием выходящих калиевых токов.
Наличие рефрактерности ограничивает частоту генерации ПД. Физиологическое значение рефрактерности заключается в создании условий для своевременного и полного осуществления восстановительных процессов в нервной клетке. Феномен рефрактерности лежит в основе понятия о функциональной подвижности, или лабильности (Н.Е. Введенский [1] ).
Лабильность — максимально возможная частота генерации ПД для данного типа возбудимой клетки. Лабильность большинства нейронов составляет приблизительно 400 ПД/с, а у интернейронов спинного мозга доходит до 1000 ПД/с.
В нормальных условиях рефрактерность предохраняет нервные клетки от излишне частой генерации ПД. Мутации, гипоксия, механические травмы и другие патологические воздействия приводят к значительным изменениям возбудимости нейронов. Такие нейроны или группы нейронов являются потенциальными источниками возникновения пароксизмальных состояний ЦНС, эпилептических припадков и других неврологических расстройств.
Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских [2] клеток в периферических нервах. Нервные волокна подразделяют на 2 типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5–8) и существенно зависит от диаметра нервных волокон.
Рис. 5–8. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых.
Безмиелиновые нервные волокна (рис. 5–9А). В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется, и наружная поверхность мембраны приобретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны.
Рис. 5–9. Проведение возбуждения в нервных волокнах [7]. А — безмиелиновое волокно (электротоническое проведение), Б — миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье [3] — проводник.
Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении.
Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5–2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения ПД (см. рис. 5–8).
Миелиновое нервное волокно (рис. 5–9Б) состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм, это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату (скачкообразное проведение).
Плотность потенциалозависимых Na+?каналов аксолеммы в перехватах Ранвье — до 2000 на 1 мкм2 (в перикарионе — 50–70, в начальном сегменте аксона — 2000, в межузловых сегментах Na+?каналы практически отсутствуют). В силу высокой плотности Na+?каналов перехваты Ранвье характеризуются высокой возбудимостью, а локальные токи достаточно велики для возбуждения соседнего перехвата.
Локальные токи текут от перехвата к перехвату (через внеклеточную жидкость кнаружи от миелина и через аксоплазму внутри аксона) с минимальными потерями.
Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется.
В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс).
Нарушение миелинизации нервных волокон приводит к нарушениям проводимости (демиелинизирующие заболевания). При разрушении миелиновой оболочки происходит резкое снижение скорости и надёжности проведения возбуждения по нервам. Наиболее распространённым среди демиелинизирующих заболеваний является множественный склероз, проявляющийся различными параличами и потерей чувствительности.
Бездекрементное проведение возбуждения. Амплитуда ПД в различных участках нерва одинакова, то есть проведение возбуждения по нервному волокну осуществляется без затухания (бездекрементно). Таким образом, кодирование информации осуществляется не за счёт изменения амплитуды ПД, а путём изменения их частоты и распределения во времени.
Изолированное проведение возбуждения. Нервные стволы обычно образованы большим количеством нервных волокон, однако ПД, идущие по каждому из них, не передаются на соседние. Эта особенность нервных волокон обусловлена:
- наличием оболочек, окружающих отдельные нервные волокна и их пучки (в результате образуется барьер, предупреждающий переход возбуждения с волокна на волокно);
- сопротивлением межклеточной жидкости (жидкость, находящаяся между волокнами, имеет гораздо меньшее сопротивление току, чем мембрана аксонов; поэтому ток шунтируется по межволоконным пространствам и не доходит до соседних волокон).
Физиологическая и анатомическая целостность. Необходимым условием проведения возбуждения является не только его анатомическая целостность, но и нормальное функционирование мембраны нервного волокна (физиологическая целостность). В клинике широко применяют различные ЛС, нарушающие физиологическую целостность нервных волокон. Так, эффекты местных анестетиков (новокаин, лидокаин, и др.) основаны на блокаде потенциалозависимых Na+?каналов. Нарушение физиологической целостности чувствительных нервных волокон вызывает анестезию (потерю чувствительности).
При регистрации электрической активности нервного ствола Джозеф Эрлангер и Герберт Гассер в 1937 г. обнаружили составной характер тока действия нервного ствола. На основании полученных данных (диаметр, скорость проведения, функция) разработана классификация (табл. 5—1), в соответствии с которой нервные волокна подразделены на группы А, В и С с дальнейшими градациями ( ? , ? , и т.д.).
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология : Учебник. - М.: ГЭОТАР-Медиа, 2009. 688 с. - Глава 5. Физиология нейронов. - Потенциал действия. С. 73-78.
[2] Шванн Теодор (Schwann T.), немецкий гистолог и физиолог (1810–1882); вместе с М. Шульце создал клеточную теорию (1839 г.); в 1836 г. открыл пепсин, в 1838 г. опубликовал первую работу по строению миелиновой оболочки.
[3] Ранвье Луи (Ranvier L.), французский патолог (1835–1922). Занимался изучением нервной ткани с применением азотнокислого серебра и хлорного золота. Его именем названы безмиелиновые участки (узловые перехваты) миелинового нервного волокна.
Электрические потенциалы, возникающие в тканях и отдельных клетках человека, животных и растений, важнейшие компоненты процессов возбуждения (См. Возбуждение) и торможения (См. Торможение). Исследование Б. п. имеет большое значение для понимания физико-химических и физиологических процессов в живых системах и применяется в клинике с диагностической целью (Электрокардиография, Электроэнцефалография, Электромиография и др.).
Различают следующие основные виды Б. п. нервных и мышечных клеток: потенциал покоя, потенциал действия, возбуждающие и тормозные постсинаптические потенциалы, генераторные потенциалы.
Потенциал покоя (ПП, мембранный потенциал покоя). У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60—90мв, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной (рис. 1). ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К + (Ю. Бернштейн, 1902, 1912; А. Ходжкин и Б. Катц, 1947). Концентрация К + в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na + примерно в 100 раз ниже, чем для К + , диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К + . В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl - , диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na + и К + падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na + значительно превышает скорость увеличения проницаемости мембраны для К + .
Постсинаптические потенциалы (ПСП) возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10—15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД (рис. 5, а, б). ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора (рис. 5, в). В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
Генераторные потенциалы возникают в мембране чувствительных нервных окончаний — рецепторов. Они внешне сходны с ВПСП — их амплитуда порядка нескольких мв и зависит от силы приложенного к рецептору раздражения (рис. 6). Когда генераторный потенциал достигает порогового (критического) значения, в соседнем участке мембраны нервного волокна возникает распространяющийся ПД. Ионный механизм генераторных потенциалов ещё недостаточно изучен.
Наряду с перечисленными относительно быстро развивающимися Б. п., в нервных клетках, волокнах гладких мышц и некоторых растительных клетках регистрируются также очень медленные колебания мембранного потенциала неизвестной природы, причём на гребне волны деполяризации мембраны часто возникают разряды импульсов.
Все Б. п. могут быть зарегистрированы и точно измерены только с помощью внутриклеточных микроэлектродов, позволяющих отводить разности потенциалов между внутренней и наружной сторонами мембраны клетки. При отведении колебаний Б. п. от целых нервов, мышц или мозга с помощью поверхностных электродов регистрируется лишь суммарно потенциал множества синхронно или, чаще, асинхронно работающих клеток. Так, электромиограмма представляет собой результат сложения (интерференции) ПД множества скелетных мышечных волокон; электрокардиограмма — результирующая колебаний электрических потенциалов мышечных волокон различных отделов сердца; электроэнцефалограмма — результат суммации главным образом ВПСП и ТПСП множества клеток различных слоев коры больших полушарий. Регистрация таких интерференционных электрограмм, хотя и не позволяет анализировать колебания Б. п. отдельных клеток, имеет важное значение для суждения о состоянии исследуемого органа в целом. В клинической практике электромиограмму, электрокардиограмму и электроэнцефалограмму регистрируют с помощью электродов, расположенных на коже соответствующих частей тела. Оценка данных, полученных этими методами, основана на сопоставлении изменений характера соответствующей кривой с результатами клинических, физиологических и патологоанатомических исследований.
Лит.; Физиология человека, М., 1966; Гальвани Л. и Вольта А., Избр. работы о животном электричестве, М. — Л., 1937; Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Экклс Дж., Физиология нервных клеток, пер. с англ., М., 1959; его же, Физиология синапсов, М., 1966; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969.
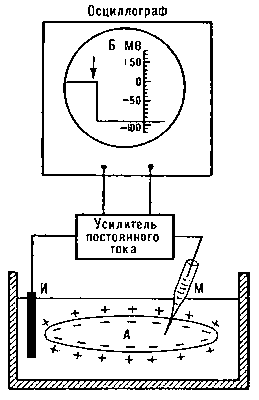
Рис. 1. Схема измерений мембранного потенциала покоя с помощью внутриклеточного стеклянного микроэлектрода (М). Второй электрод (И) помещен в омывающую клетку жидкость.
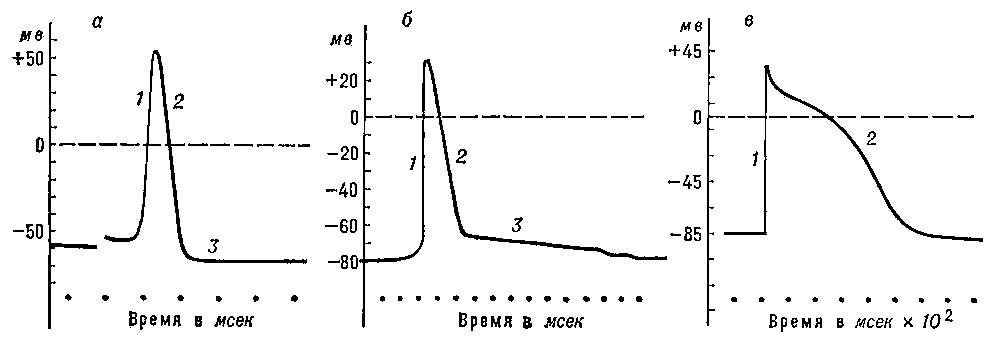
Рис. 2. Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов: а — гигантского аксона кальмара; б — скелетного мышечного волокна; в — волокна мышцы сердца собаки; 1 — восходящая фаза ПД; 2 — нисходящая фаза; 3 — следовая гиперполяризация (а) и следовая деполяризация (б).
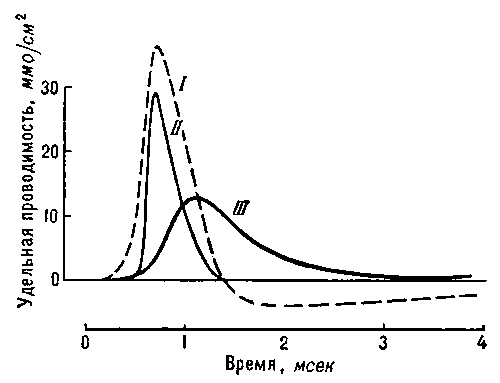
Рис. 3. Изменения натриевой и калиевой проводимости мембраны нервного волокна во время генерации потенциала действия (I). Изменения проводимости пропорциональны изменениям проницаемости для Na + (II) и К + (III).
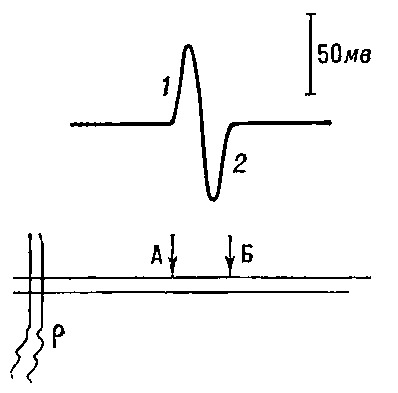
Рис. 4. Регистрация распространения потенциала действия вдоль нервного волокна. А, Б — внеклеточные электроды; р — раздражающие электроды. Вверху — отклонение луча осциллографа под влиянием волны возбуждения; 1 — волна возбуждения под электродом А; 2 — под электродом Б.
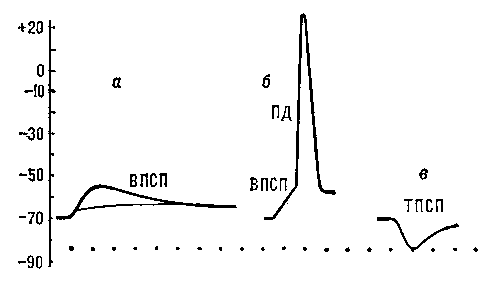
Рис. 5. Постсинаптические потенциалы: а — подпороговые ВПСП, возникающие в нервной клетке в ответ на раздражения соответствующих нервных волокон; б — ВПСП, достигший порогового значения, достаточного для возникновения ПД; в — ТПСП, вызванный раздражением тормозных нервных волокон.
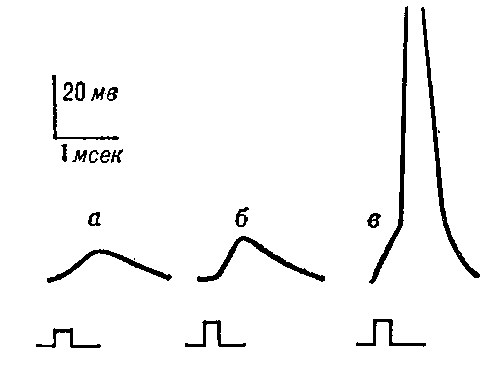
Рис. 6. Генераторные потенциалы: увеличение амплитуды при усилении раздражения рецептора (а — в). При достижении порогового значения (в) генераторный потенциал вызвал в чувствительном нервном волокне потенциал действия.
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
На рисунке показаны изменения, возникающие на мембране во время потенциала действия, с переносом положительных зарядов внутрь волокна вначале и возвращением положительных зарядов наружу в конце. В нижней части рисунка графически представлены последовательные изменения мембранного потенциала в течение нескольких 1/10000 сек, иллюстрирующие взрывное начало потенциала действия и почти столь же быстрое восстановление.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Временной ход потенциала действия в нейроне; показаны последовательные фазы потенциала действия, описанные в тексте.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na + /K + -насосу и каналам К + /Na + -утечки.
Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна.
Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз.
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
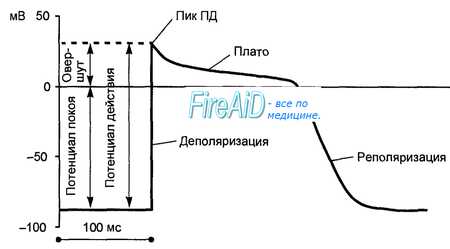
Потенциал действия клетки рабочего миокарда.
Быстрое развитие деполяризации и продолжительная реполяризация. Замедленная реполяризация (плато) переходит в быструю реполяризацию.
- Вернуться в оглавление раздела "Физиология человека."
Читайте также: