
Утилизация глюкозы из цнс

В настоящее время признано, что диагноз "шизофрения" определяют лишь синдром разных расстройств и что изучение данного синдрома в целом препятствует выявлению основных звеньев патогенеза внутри синдрома "шизофрении".
Исследования экспрессии генов в центральной нервной системе человека (ЦНС) позволяют предположить, что нарушение регуляции метаболических путей играет важную роль в этиопатогенезе шизофрении. Тезис о том, что изменения в метаболических путях нарушаются у больных шизофренией, получил дальнейшее подтверждение в исследованиях протеома и метаболома в тканях головного мозга при этом психическом расстройстве.
Важно отметить, что исследования метаболизма ЦНС в целом указывают на роль изменений метаболизма глюкозы в патогенезе шизофрении. Это важно, потому что ЦНС не имеет обширных энергетических резервов в форме гликогена и, следовательно, в значительной степени зависит от получения своих энергетических потребностей из доступной глюкозы. Кроме того, недавно было высказано предположение, что нейронная активность может сильно зависеть от глюкозы. Если высокие уровни глюкозы находятся во внеклеточной среде, это может свидетельствовать о неспособности стриатных клеток поглощать глюкозу, что согласуется с результатами исследований нейровизуализации при шизофрении.
Интересно отметить, что лечение крыс хлорпромазином подавляет гликолиз в ЦНС, но не на периферии. Кроме того, было показано, что лечение галоперидолом снижает уровень глюкозы и пирувата в общем мозге, но не вызывает изменений в уровнях ацетил-КоА и повышает уровни лактата в мозге. Наконец, сообщалось, что галоперидол не изменял скорости метаболизма полосатого тела, измеренные с использованием [18F] фтордезоксиглюкозы у галоперидол-чувствительных пациентов с шизофренией. Также отмечено, что лечение типичными антипсихотическими препаратами само по себе не изменяет метаболизм глюкозы в ЦНС.
Таким образом, любое изменение метаболизма глюкозы может повлиять почти на все клеточные процессы в нейронах и нейроглии, влияя на процессы, включая нейротрансмиссию и скорость гибели клеток. Кроме того, изменения метаболизма глюкозы в ЦНС могут особенно влиять на функцию нейронов, потому что для каждой клетки требуется наибольшего количества энергии и, следовательно, необходим быстрый доступ к глюкозе для метаболизма. Как и в любой клетке, нейроны метаболизируют глюкозу в течение ряда этапов, конечным продуктом которых является генерация клеточной энергии в ряде форм, включая АТФ, причем , последняя также может выступать в роли нейротрансмиттера сама по себе. Таким образом, любые значимые изменения в метаболизме глюкозы в нейронах, в частности, окажут глубокое влияние на функцию мозга, что может привести к таким расстройствам, как шизофрения.
Двумя путями, которые играют главную роль в метаболизме глюкозы, являются путь Эмбдена-Мейерхофа ( Embden-Meyerhof) и цикл трикарбоновых кислот (ТСА). Путь Embden-Meyerhof метаболизирует глюкозу в пируват с чистой продукцией двух молекул АТФ и двух молекул восстановленного никотинамидадениндинуклеотида (NADH), оба из которых представляют собой энергетический обмен для клетки. Затем пируват может быть превращен в ацетил-КоА комплексом пируватдегидрогеназы, причем ацетил-КоА является субстратом для цикла ТСА. Когда одна молекула субстрата, ацетил-КоА, обрабатывается через TCA, происходит высвобождение электронов из NADH и образуется восстановленный флавин-адениндинуклеотид (FADH 2) для облегчения производства 4 молекул АТФ.
В литературе сообщалось, что активность аконитазы, комплекса α-кетоглутаратдегидрогеназы и сукцинат тиокиназы ниже в дорсолатеральной префронтальной коре у больных шизофренией. Эти данные важны, потому что эти ферменты катализируют первые три реакции TCA. Кроме того, исследование экспрессии массива генов показали более низкие уровни мРНК для сукцинатдегидрогеназы комплекса субъединицы А и лактатдегидрогеназы А в гиппокампе больных шизофренией. Сукцинатдегидрогеназа является еще одним ферментом в цикле TCA, тогда как лактатдегидрогеназа может превращать пируват в лактат - продукт, который не вступает в путь Embden-Meyerhof. Эти данные свидетельствуют о том, что изменения метаболизма глюкозы в ЦНС у субъектов с шизофренией могут быть широко распространены в ряде регионов.
Полосатое тело при шизофрении
Исследования , проведенные с помощью нейровизуализации были использованы для оценки метаболизма глюкозы в ЦНС человека, причем , в таких исследованиях сообщалось об изменениях метаболизма глюкозы при шизофрении. Важно отметить, что об изменениях в утилизации глюкозы в стриатуме ( полосатом теле ) у лиц, страдающих шизофренией ,сообщалось в ряде исследований нейровизуализации. Эти данные предполагают, что полосатое тело может быть особенно затронуто изменениями метаболизма глюкозы у пациентов с шизофренией. Несмотря на такие данные нейровизуализации, посмертные исследования, по-видимому, не фокусировались на полосатом теле для того , чтобы обнаружить механизмы, которые могут лежать в основе изменений метаболизма глюкозы у больных шизофренией.
Возникает предположение , что аномальный метаболизм глюкозы в полосатом теле способствует патогенезу шизофрении. Эта гипотеза обусловлена данными о низком уровне PDHB и более высоких уровнях пирувата, а также о тенденции к более высоким уровням ацетил-КоА в стриатуме у больных шизофренией. Эти результаты будут соответствовать снижению активности цикла TCA и / или пути Эмбдена-Мейера у пациентов с шизофренией.
Лактат
Одним из подходов к определению механизмов, посредством которых метаболизм глюкозы изменяется в ЦНС у больных шизофренией, может быть измерение активности, уровней белка или уровней экспрессии ферментов в таких путях, как путь Эмбдена-Мейерхофа или TCA. Однако другой подход заключается в измерении уровней отдельных метаболитов как суррогатных маркеров изменений активности глюкозы с использованием метаболических путей. При шизофрении сообщалось о более низких уровнях лактата в передней части внутренней капсулы, однако , более высокие уровни этого метаболита были обнаружены в мозжечке и коре головного мозга больных шизофренией. Кроме того, по сравнению с контролем, в медиодорсальном таламусе у пациентов с шизофренией отмечены более низкие уровни пирувата. В аэробных условиях, которые, вероятно, преобладают большую часть времени в ЦНС, более низкие уровни пирувата могут свидетельствовать об увеличении обработки этого субстрата комплексом пируватдегидрогеназы или уменьшении продукции пирувата по пути Эмбдена-Мейерхофа. Напротив, превращение пирувата в лактат и лактатный анализ в основном связаны с анаэробным метаболизмом, процессом, который, как известно, начинает доминировать через некоторое время после смерти. Чтобы различать эти процессы, важно отметить, что комплекс пируватдегидрогеназы является стадией, ограничивающей скорость превращения пирувата в ацетил-КоА; следовательно, можно ожидать, что изменения, включающие аэробный метаболизм, будут очевидны из-за изменений в активности или уровнях комплекса пируватдегидрогеназы, которые будут сопровождаться обратными изменениями субстрата фермента (пирувата) и его продукта (ацетил-КоА) без изменения в анаэробном продукте.
Комплекс пируватдегидрогеназы
Комплекс пируватдегидрогеназы состоит из трех ферментов: пируватдегидрогеназы, дигидролипоил трансацетилазы и дигидролипоилдегидрогеназы. Пируватдегидрогеназа представляет собой четырехсторонний комплекс, состоящий из 2 α и 2 β субъединиц, каждая из которых играет роль в регулировании активности пируватдегидрогеназы и комплекса пируватдегидрогеназы. Можно ожидать, что любое изменение уровней любой из субъединиц комплекса пируватдегидрогеназы изменит активность комплекса пируватдегидрогеназы и, следовательно, уровней пирувата и ацетил-КоА. Поэтому сообщение об изменениях уровней пирувата в ЦНС у больных шизофренией будет соответствовать изменениям метаболизма пирувата пируватдегидрогеназным комплексом.
Дефицит мускариновых рецепторов
Некоторые авторы показали, что можно выделить подгруппу больных в пределах шизофрении, основываясь на заметной потере связывания коркового мускаринового рецептора - подмножество больных шизофренией с дефицитом мускариновых рецепторов (MRDS). Показано, что у субъектов с MRDS наблюдается широко распространенная потеря кортикальных мускариновых рецепторов, сходная с той, которая была отмечена у некоторых больных в исследовании нейроизображения мускариновых рецепторов при шизофрении. У больных с MRDS также уменьшилось связывание радиолиганда со стриатными мускариновыми рецепторами и более высокие уровни белка постсинаптической плотности 95, показывающие изменения в молекулярной цитоархитектуре ЦНС у больных с MRDS, простирающимися за пределы коры. Отметим, что уже давно было показано , что мускариновые рецепторы, действуют как модуляторы поглощения глюкозы в ЦНС и что компоненты путей метаболизма глюкозы, такие как PDH, могут модулировать уровни активности мускариновых рецепторов.
Данные ряда исследований подтвердили гипотезу, согласно которой изменения глюкозы, пирувата, PDHB и ацетил-КоА ограничены субъектами с не-MRDS, тогда как изменения в лактате происходят при MRDS. Эти данные позволяют предположить , что изменения в метаболизме глюкозы в полосатом теле могут быть основной причиной возникновения симптомов у больных без дефицита мускариновых рецепторов. Поскольку изменения в маркерах метаболизма глюкозы обнаруживаются только у субъектов с не-MRDS, но не у MRDS, у которых заметен более низкий уровень мускариновых рецепторов в их коре и стриатуме, и может быть значительным, отсюда , можно седлать вывод о наличии двунаправленных связей между активностью и / или уровнями мускариновых рецепторов и уровнями метаболизма глюкозы. В частности , низкие уровни мускариновых рецепторов ЦНС могут быть защитными от изменений метаболизма глюкозы у подгруппы больных шизофренией с MRDS.
Ранее в литературе сообщалось , что субъекты с MRDS имеют широко распространенную потерю кортикальных мускариновых рецепторов, нарушения в передаче сигналов мускаринового рецептора M1, четкую картину метилирования промотора гена рецептора мускаринового M1 и более высокие уровни микроРНК. который действует для того , чтобы уменьшить экспрессию рецептора, и иметь ослабленный ответ на мускариновый M1-рецептор-положительный аллостерический модулятор.
Нам надо понять для чего мы едим так много сладкого, почему страдаем в том числе диабетом.
Сегодня мы продолжаем ту развернутую тему, о которой мы долго говорили и с интересом. Это сосудистая система и способы питания, включая каннибализм.
Все это называется нейроэнергетика. Это то что происходит с обменом в мозге. Нам надо понять для чего мы едим так много сладкого, почему страдаем в том числе диабетом. Мозг питается всем тем комплексом, который содержит белки, жиры и углеводы плюс микроэлементы.
В детстве, когда мы развиваемся в утробе матери и после рождения у нас огромную роль играют китоновые тела. Плюс в мозге есть еще очень важный компонент -это гликоген. Его много в мышцах.
Вся мускулатура работает за счет расщепления этого глюкогена, а затем использование его энергии через синтез атф в митохондриях для того, что бы наши мышцы сокращались.
В мозге его меньше в 10 раз, но тем не менее он используется. Это тот самый страховочный круг, который дает нейрону, в случае каких то катаклизмов, травм или еще чего то, прожить в мозге примерно 100 минут до начала необратимых изменений.
Дело в том, что сахар в мозге играет огромную роль. Его в нейронах на шесть минут запаса. Плюс небольшой запас в окружающей глии.
Что же происходит с глюкозой, когда она попадает в головной мозг? Глюкоза ничто иное как главный источник энергии для нейронов, соответственно наше поведение заточено, что бы эту глюкозу найти.
Поэтому если детей совсем решать сладкого, то проблемы будут с развитием. Развивающийся мозг и его отростки требуют огромного количества АТФ, которая является химическим способом запасания энергии в нашем головном мозге. А образуется она достаточно интенсивно и быстро из этой самой глюкозы или фруктозы .
У нас на кончике языка сосредоточены все рецепторы сладкого. Т.е главное это найти корм для мозга и мускулатуры. Так как в мускулатуре запасается глюкоген.
Первая порция пищи, независимо от того что вы едите, превращается в два метаболита, которые доходят до мозга через эту самую сосудистую систему, на которую вы потратили столько времени.
Какие это два метаболита?
В первую очередь -это сахар, а во вторую алкоголь. Наш мозг и сосуды так устроены, что наш мозг может узнать о том, что вы съели что то стоящее, если из этого стоящего можно извлечь алкоголь и сахар.
Поэтому мы такие блаженные и счастливые, если долго не ели. Поэтому все трезвеники, но любители поесть что то сладенькое-это хронические алкаши.
Наш эндогенный алкоголь образуется у нас по биологическим законам. Для каждой этнической группы есть свой минимальный уровень внутреннего образующегося в организме алкоголя. Это обычно и учитывается, что бы не преследовать людей, которые просто плотно поели.
Если вы не делаете ничего, то в этом состоянии мозг потребляет 10 % из нее довольно много идет глюкозы. Глюкозы может уходить до 30-40 %, если вы не занимаетесь физическим трудом. Еще 20-25 % кислорода, как раз кислород для того, что бы окислять эту глюкозу. Т.е дыхание как таковое, это добыча кислорода из глюкозы. В результате глюкоза расщепляется, там длинная ферматотивная реакция, до углекислого газа и воды. А в качестве побочного продукта, т.е целей этих всех АТФ, где в химических связях запасается та самая энергия, которая нам нужна.
Зачем же нужно такое количество энергии? Оказывается основной расход -это синоптический контакт. На создание мембран тоже нужно, для транспорта тех самых метаболитов, которые передают сигнал от клетки к клетке, тоже нужно много энергии. Поэтому нейрон пашет, для того что бы обеспечить свои элементарные функции и никаких запасов гигантских нет.
Внутри самого мозга, эта глюкоза потребляется с дикой скоростью. Но если образуется избыток, то происходит несчастье. У нас накапливаются высокие сахара и их надо утилизировать. Эта утилизация происходит с помощью инсулина.
У нас существует целая система внутренней рецепции на наличие или отсутствие глюкозы.
Вдруг захотелось сладенького, с чего бы это? Я вам рассказывал, что у нас питьевое поведение регулируется рецепторами находящимися внутри желудочка мозга, которые создают иллюзию пересыхания губ и пересыхания рта, что бы заставить попить. Вот примерно такая же ерунда у нас существует с глюкозой. Т.е это желание возникает внутри головы.
Оказалось, что глюкоза- чувствительные нейроны существуют в самых различных частях мозга. Моторно человек должен чувствовать есть у него запас сахара или нет.
Когда глюкозы не хватает, то мозгом запускается очередное мелкое надувательство, фантомное ощющение и вас тянет на сладенькое и это значит мозг ваш успел поработать.
Церебральный метаболизм глюкозы достаточно забавен, пройдя через гематоцефалический барьер, он конвектируется и через цикл трикарбоновых кислот происходят накопления АТФ.
Но проблема заключается в том, что производится много побочных продуктов, которые говорят о том что вы приняли пищу.
Когда начинается цикл трикарбоновых кислот в результате расщепления глюкозы образуется 18 молекул.Среди побочных продуков есть глютомат. Его добавляют в фаст фуд и даже запускают через вентеляционные трубы, что бы вы думали что едите что то очень вкусное.
Глюкомат является конечной молекулой, когда прошел весь цикл трикорбанатных кислот. Т.е нейроны понимают, что их накормили не какой-то дрянью, а чем то ценным, глюкозой, а побочный продукт глюкомат.
А если вы его в фаст фуде распылите в воздух и граждане надышатся им, то у них начинается райское наслаждение до того как они съели фальшивую котлету. Таким образом возникает наркотическая зависимость от принятия пищи.
А дальше все эти продукты распада надо выбросить и они естественно выносятся и использоваться так что бы СО2 выходил из воздуха. Т.е ваше дыхание это инструмент для запасания энергии внутри мозга в виде химических связей.
Когда вы вдыхаете кислород доносится до мозга, там идет на утилизацию глюкозы и потом дальше выходит. Т.е 25% ваших легких работает исключительно на мозг.
Соответственно мозг обладает способом ее регуляции. Информация о том, что обмен сахаров зависит только от инсулина и поджелудочной железы сходит на нет. И все больше и больше информации о том, что концентрация глюкогена в мозге чувствительна к глюкозе концентрауии инсулина и глюкозы. Получено масса информации о том, что все эти события мозг не может не реугулировать собственный энергетический баланс.
Многие запасные вещества такие как сахар и гликоген, которые сидят в астроцитов могут быть вовлечены. Что происходит? Многие диабетики у них страдают мозг, зрение и почки.
Мозг очень страдает, потому оно не может не влиять на обмен сахара, который является ключевым элементом в энергетики мозга. При осложнение диабета возникает суперкомпенсация при первом падении сахара и спустя некоторое время сознание судорожного состояния депрессии, которые зачастую перерастают в Альтцгемера. Таким образом мозг не только потребитель сахара, он еще и сложнейший инструментом для регуляции сахарного обмена.
А сейчас отвечу на некоторые вопросы.
Вопрос: Можно ли повысить эффективность работы мозга за счет равномерного и дозированного потребления глюкозы из аптеки в течение рабочего дня?
Ответ: Из аптеки ничего потреблять нельзя. Лучше чай с медом, но это если вы его расходуете.
Если у вас и физическая и интеллектуальная нагрузка. А то вы так в течение дня будете хлебать и через месяц у вас будет сахарный диабет.
В Зубцовском университете, Сани Микулович вскрыла нейроны храбрости, в гипокампе.
Естественно, что нейроны возбуждаются, когда происходят хоть какие -то нейрогармональные стрессы. Нашла их конечно у самых храбрых мышей. Это так называемые LM нейроны.
Они делали мышам электроэнцелографию, при этом мышь пугали волосом от кошки. Мышь пугалась ужасно и пряталась. А вот если начинали долбить электронными нейронами, то мышь оказывается успокаивалась.
Представляете, если у вас в норме обычно стимулируется электродами, которые вам воткнули, потом ходишь как чумной, как после позитронной томографии. То конечно такая ерунда как волос кошки мышей переставал раздражать.
Есть еще прекраснейшая работа, пришедшая нам из Торонто. Исследование было сделано, что бы показать, а полезно или вредно детям врать?
Если ваш ребенок много врет, то оказывается это прекрасно. Потому что ложь детская стимулирует когнитивные функции, т.е получается лживость позволяет придумывать второй слой. Пришли к выводу, что если детей в эксперименте научить обманывать и оказалось, что отбор мелких мерзавцев очень эффективен. Поскольку их когнитивные функции развивались.
Это говорит о том, что то что происходило во время эволюции головного мозга в Европе, когда церковь боролась с инокомыслием, отбирались самые хитрые, лживые и подлые. Поэтому Европа такая процветающая.
Так что учите своих детей врать, смотришь они и поумнеют.
Из материалов авторской программы профессора Сергея Вячеславовича Савельева "Вынос мозга" (выпуск №56 от 15.09.2018)
После приёма пищи большое количество глюкозы поступает в печень по воротной вене. Эта глюкоза используется прежде всего для синтеза гликогена (схема реакций приводится на рисунке 2). Содержание гликогена в печени здоровых людей обычно составляет от 2 до 8% массы этого органа.
Гликолиз и пентозофосфатный путь окисления глюкозы в печени служат в первую очередь поставщиками метаболитов-предшественников для биосинтеза аминокислот, жирных кислот, глицерола и нуклеотидов. В меньшей степени окислительные пути превращения глюкозы в печени являются источниками энергии для обеспечения биосинтетических процессов.

Рисунок 1 Главные пути превращения глюкозо-6-фосфата в печени. Цифрами обозначены: 1 - фосфорилирование глюкозы; 2 - гидролиз глюкозо-6-фосфата; 3 - синтез гликогена; 4 - мобилизация гликогена; 5 - пентозофосфатный путь; 6 - гликолиз; 7 - глюконеогенез

Рисунок 2 Схема реакций синтеза гликогена в печени

Рисунок 3 Схема реакций мобилизации гликогена в печени
В некоторых условиях (при голодании низкоуглеводной диете, длительной физической нагрузке) потребность организма в углеводах превышает то количество, которое всасывается из желудочно-кишечного тракта. В таком случае образование глюкозы осуществляется с помощью глюкозо-6-фосфатазы, катализирующей гидролиз глюкозо-6-фосфата в клетках печени. Непосредственным источником глюкозо-6-фосфата служит гликоген. Схема мобилизации гликогена представлена на рисунке 3.
Мобилизация гликогена обеспечивает потребности организма человека в глюкозе на протяжении первых 12 - 24 часов голодания. В более поздние сроки основным источником глюкозы становится глюконеогенез - биосинтез из неуглеводных источников.
Основными субстратами для глюконеогенеза служат лактат, глицерол и аминокислоты (за исключением лейцина). Эти соединения сначала превращаются в пируват или оксалоацетат - ключевые метаболиты глюконеогенеза.
Глюконеогенез - процесс, обратный гликолизу. При этом барьеры, создаваемые необратимыми реакциями гликолиза, преодолеваются при помощи специальных ферментов, катализирующих обходные реакции (см. рисунок 4).
Из других путей обмена углеводов в печени следует отметить превращение в глюкозу других пищевых моносахаридов - фруктозы и галактозы.

Рисунок 4 Гликолиз и глюконеогенез в печени
Ферменты, катализирующие необратимые реакции гликолиза: 1 - глюкокиназа; 2 - фосфофруктокиназа; 3 - пируваткиназа.
Ферменты, катализирующие обходные реакции глюконеогенеза: 4 -пируваткарбоксилаза; 5 - фосфоенолпируваткарбоксикиназа; 6 -фруктозо-1,6-дифосфатаза; 7 - глюкозо-6-фосфатаза.
- 6837
- 5,3
- 9
- 8
Чтобы голова была светлой, а чистый разум воссиял, клеткам мозга пришлось освоить разные профессии, разделив функции уже на этапе утилизации источников энергии
![]()
Евгения Самохина

![]()
Антон Чугунов![]()
Андрей Панов




Спонсор номинации — Future Biotech, проект, объединяющий профессионалов и энтузиастов в области биологии и биотехнологий.
Генеральным спонсором конкурса, согласно нашему краудфандингу, стал предприниматель Константин Синюшин, за что ему огромный человеческий респект!
Общеизвестно, что для нормального функционирования всех органов нашего тела необходима энергия. Бόльшую часть энергии при обычном рационе человек получает путем превращения поступающих в организм углеводов в глюкозу и разложения последней до углекислого газа и воды, что сопровождается запасанием необходимой для нас энергии в виде аденозинтрифосфатов (АТФ) или других макроэнергетических соединений.
Роли предопределены
Нейроны — это высокоспециализированные клетки, способные генерировать и проводить электрические импульсы. Это — клетки-специалисты, так как функция каждого нейрона строго определена. В течение долгого времени происходит так называемый процесс обучения нейрона выполняемой им функции. Средний человеческий мозг содержит около 100 миллиардов обученных нейронов, и в среднем каждый нейрон соединяется с 1000 других нейронов. Это приводит к образованию обширных и сложных нейронных сетей, которые служат основой для обработки и передачи мозгом информации. Ввиду сложных интегративных взаимодействий каждого нейрона замена этих клеток в нейронных сетях с сохранением целостности выполняемой ими функции почти (если не совсем) невозможна.
Астроциты — это специализированные глиальные клетки, чья функция заключается главным образом в обеспечении нейронов энергетическими ресурсами и в борьбе с активными формами кислорода (АФК) и азота [4]. При этом количество астроцитов в мозге в несколько раз превышает количество нейронов, и в результате получается, что каждый нейрон включен в целый ансамбль астроцитарных клеток.
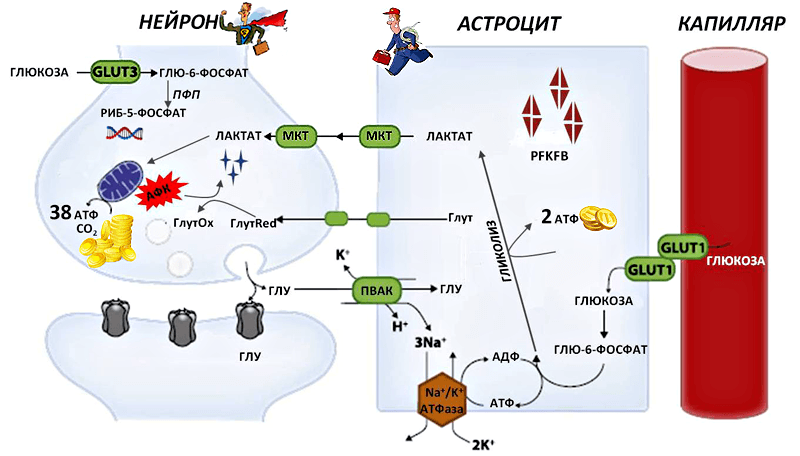
Рисунок 1. Схема метаболических взаимодействий между нейронами и астроцитами. Глутамат (ГЛУ) — нейромедиатор, высвобождающийся из синаптического окончания нейрона. Часть высвобожденного глутамата поглощается астроцитами с помощью переносчиков возбуждающих аминокислот (ПВАК) совместно с тремя ионами Na + . Ионы затем выталкиваются с помощью работы Na + /K + -АТФазы, потребляющей энергию в форме аденозинтрифосфата (АТФ). Это стимулирует поглощение глюкозы астроцитами. С помощью переносчиков (GLUT1) глюкоза из капилляра поступает в астроцит и в процессе гликолиза превращается в лактат. При этом освобождается две молекулы АТФ. Лактат (молочная кислота) посредством специальных переносчиков (МКТ) поступает в нейрон и после нескольких превращений, в том числе в митохондриях, дарит клетке 38 молекул АТФ. Сами нейроны тоже могут поглощать глюкозу — посредством рецепторов GLUT3. Глюкозо-6-фосфат, образовавшийся в нейроне из глюкозы, направляется в пентозофосфатный цикл, который поставляет предшественников для синтеза нуклеотидов ДНК и РНК. Регулирует гликолиз в нейронах и астроцитах фермент PFKFB. Предшественники антиоксидантной (глутатионовой) системы нейрона (Глут) также поступают в него от астроцитов и участвуют в обезвреживании АФК, превращаясь из восстановленной формы (ГлутRed) в окисленную (ГлутOx).
Строгий контроль
Для точной настройки скорости гликолиза (высокой в астроцитах и относительно низкой — в нейронах) во всех клетках мозга работает регуляторный фермент 6-фосфофрукто-2-киназа/фруктозо-2,6-бисфосфатаза (PFKFB) [7]. Высокий уровень ферментативной активности PFKFB в астроцитах способствует высокой скорости в них гликолитических реакций. Однако что произойдет, если нейроны снизят скорость основного ПФП и, подобно астроцитам, наладят процессы гликолиза? Экспериментально показано: за этим последует катастрофа — гибель нейронов. Дело в том, что такое усиление гликолиза в нейронах ведет к сокращению образования фермента антиоксидантной системы — глутатиона (между прочим, единственного пептидного вещества, образующегося непосредственно в нейронах и спасающего их от окислительного стресса), усилению окислительного стресса и наконец к апоптотической гибели клетки. Таким образом, разделение энергетических путей оказывается процессом, строго приспособленным к повышению выхода энергии мозгом и одновременно очень консервативным с точки зрения возможности реализации в разных типах клеток.
Согласно наиболее популярной сейчас точке зрения, именно в изменении энергетического состояния мозга лежит причина (по крайней мере, одна из главных причин) судорожных состояний и гибели клеток в структурах мозга [8]. В результате снижения энергообеспечения клеток мозга из-за травм, ишемии или опухоли под ударом оказываются в первую очередь системы регуляции тормозных процессов в нервной ткани. Как ни странно, именно тормозные процессы требуют от нейронов мобилизации энергетических затрат. Недостаток энергии приводит к неспособности клеток затормозить возбуждение и к постепенному распространению возбуждающей волны во все области мозга. Неконтролируемая постоянная активация клеток вызывает еще большее истощение их энергетических запасов и приводит к окислительному стрессу. В результате падения активности антиоксидантной защиты ниже критического уровня происходят необратимые изменения в клетках. Формируется замкнутая цепь губительных событий: судорожная активность вследствие развившегося дефицита энергии в одних структурах мозга вызывает новые эпизоды приступов. И получается, что, однажды начавшись, судороги постоянно порождают новые судороги.
В исследованиях механизмов развития эпилептической активности было установлено, что судорожные приступы развиваются в первую очередь при наследственных заболеваниях, нарушающих нормальный метаболизм энергии в мозге [9]. Причем резкое снижение главного источника энергии — глюкозы — даже у людей, не страдающих эпилепсией, приводит к тяжелым судорожным припадкам [10]. Аналогичный эффект наблюдается у людей, страдающих эпилепсией, после сна, когда концентрация глюкозы в крови резко падает из-за длительного перерыва в поступлении пищи, то есть примерно восьмичасового голода [11].
d) повышенным всасыванием глюкозы в кишечнике
24. Нормой глюкозы в крови следует считать:
25. Легкоусвояемыми углеводами следует считать:
B) глюкозу
26. Межуточный обмен углеводов нарушается при:
A) недостатке инсулина
B) гипоксиях
c) недостатке желчи
27. Нарушение всасывания глюкозы в кишечнике наблюдается при наследственном дефиците:
b) глюкозо 6-фофатазы
c) кислой α-1,4-глюкозидазы
28. Болезнь Помпе развивается при наследственном дефиците:
b) глюкозо 6-фофатазы
c) кислой α-1,4-глюкозидазы
29. Инсулин стимулирует:
a) аэробный гликолиз
30. Для какого вида комы при сахарном диабете характерна наиболее высокая гипергликемия:
31. Инсулин препятствует:
a) аэробному гликолизу
32. При недостатке инсулина в крови повышается концентрация:
c) кетоновых тел
33. Болезнь Гирке – это болезнь накопления:
34. Почечный порог – это уровень глюкозы:
a) в крови, при котором она попадает в первичную мочу
b) в первичной моче, поступающей во вторичную мочу
c) в крови, превышение которого делает невозможным ее полную реабсорбцию
35. Причинами гипогликемии может быть повышение в крови:
36. Причинами гипогликемии может быть:
37. Причинами сахарного диабета первого типа являются:
a) недостаточное образование проинсулина
b) повышенный катаболизм инсулина
c) дефицит рецепторов к инсулину на органах-мишенях
38. Инсулин обеспечивает поступление глюкозы в:
a) жировую ткань
39. Признаками гипогликемии тяжелой степени являются:
a) нарушение координации движений
c) запах ацетона из рта
d) потеря сознания
40. Для сахарного диабета I типа характерны:
41. Запах ацетона изо рта наблюдается при:
a) гипогликемической коме
b) почечном диабете
c) кетоацидотической коме
d) гиперосмомолярной коме
42. К осложнениям сахарного диабета относятся:
a) гипогликемическая кома
b) гиперосмолярная кома
c) поражение сосудов почек
D) гангрена
43. К осложнению сахарного диабета следует отнести:
A) анемию
B) ретинопатию
44. В моче при сахарном диабете присутствуют:
d) кетоновые тела
45. Углеводное голодание приводит к:
a) увеличению образования кетоновых тел
c) увеличению синтеза белка
d) не имеет последствий
46. При сахарном диабете нарушается:
a) только белковый обмен
b) только жировой обмен
C) все виды обменов
D) только углеводный обмен
E) только водно-солевой обмен
47. Гипогликемическая кома может развиться при:
b) недостатке инсулина
c) гиперфункции коры надпочечников
D) инсуломе
48. Межуточный обмен углеводов нарушается при:
a) гиповитаминозе В1
B) гипоксиях
c) недостатке желчи
49. Для гипогликемической комы характерно:
a) ацетон в моче
b) дыхание типа Куссмауля
c) мягкие глазные яблоки
d) снижение глюкозы в крови
Ответы:
1bd, 2b, 3b, 4d, 5a, 6a, 7bcd,8ad, 9a, 10bcd, 11d, 12ac, 13bc, 14ad, 15ab, 16d, 17ad, 18c, 19ad, 20bd, 21bd, 22d, 23ac, 24b, 25b, 26ab, 27a, 28c, 29ac, 30b, 31bcd, 32acd, 33d, 34c, 35a, 36abd 37a, 38ab, 39bd, 40ac, 41с, 42bcd,43b, 44cd, 45bc, 46ab, 47c, 48ab, 49d.
Читайте также: