
Вестибулярный ганглий расположен в
Вестибулярный анализатор (статокинетический) — нейродинамическая система, осуществляющая восприятие и анализ информации о положении и движении тела в пространстве. Вестибулярный анализатор состоит из рецепторов, проводящих путей (афферентных и эфферентных), промежуточных центров и коркового отдела. Поступающие в ц. н. с. импульсы от вестибуло- и проприорецепторов обусловливают возникновение вестибуло-моторных (тонических), вестибуло-сенсорных и вестибуло-вегетативных рефлексов.

Рецепторы Вестибулярного анализатора расположены в перепончатом лабиринте внутреннего уха (см.). Лабиринт представлен тремя полукружными каналами и двумя мешочками — utriculus и sacculus. Каждый канал выходит из utriculus и, описав две трети круга, вновь в него впадает. На месте впадения имеется расширение — ампула, а внутри последней — гребешок (crista ampullris), поверхность к-рого образована слоем волосковых (сенсорноэпителиальных) клеток; одни из них имеют форму кувшина и локализуются на верхушках крист, другие — цилиндрической формы, расположены в склонах; они более организованны и реагируют, видимо, на более локальные раздражения. Те и другие клетки несут на свободном конце сенсорные волокна, каждое из которых представляет собой пучок, состоящий из 70— 80 неподвижных стереоцилий и одной длинной подвижной киноцилии на периферии. Гребешок (рис., А, нижний) покрыт студенистой массой (cupula), примыкающей вплотную к стенке ампулы, в силу чего под влиянием сдвигов эндолимфы происходит ее смещение. Рецепторы мешочков (macula) также состоят из волосковых клеток двух типов и слоя опорных клеток. Над сенсорным эпителием лежит желатиноподобная мембрана, в к-рую вкраплены известковые образования — отолиты (рис., А, верхний). Между отолитами и macula имеется узкое субмембранное пространство, к-рое дает возможность отолитовой мембране скользить по macula и деформировать волоски сенсорных клеток. Macula utriculi имеет форму вогнутого овального пятна размером 3x1 —5 X 2 мм и располагается в основном по дну, по передней и медиальной стенке utriculus. Macula sacculi лежит на медиальной его стенке, имеет овальную форму (0,48 мм 2 ) и образует с macula utriculi угол ок. 90° (см. Отолитовый аппарат).
Естественными раздражителями В. а. являются инерционные сдвиги эндолимфы, возникающие в полукружных каналах при воздействии поворотных и кориолисовых ускорений (см. Ускорение), и перемещение отолитов (преимущественно, видимо, скольжение) относительно волосковых клеток при прямолинейных ускорениях и измененной силе тяжести. Расположение полукружных каналов соответственно плоскостям трехмерного пространства позволяет рецепторам реагировать на инерционные силы, возникающие практически при любых перемещениях тела (активных или пассивных), изменениях величины и вектора гравитации.
От рецепторов лабиринтов начинаются первичные нервные волокна. Первый нейрон проводящей системы находится в вестибулярном ганглии, лежащем в глубине внутреннего слухового прохода и представляющем скопление клеток с двумя отростками. Длинные отростки нейрона составляют вестибулярный нерв (см. Преддверно-улитковый нерв), несущий импульсы ко вторым нейронам — комплексу ядер в мосту, проецирующихся на латеральные углы ромбовидной ямки (area vestibularis).
Вестибулярные волокна оканчиваются в основном на нейронах в центральной части верхнего ядра и латерального ядра, в задне-латеральной части медиального ядра и в задне-медиальной части нижнего преддверного ядра. Часть первичных волокон, не прерываясь в ядрах, направляется в мозжечок. В ядрах берут начало вторичные вестибулярные волокна и направляются в составе медиального продольного пучка к моторным нейронам глазодвигательной мускулатуры и через вестибуло-спинальный тракт до нижних сегментов спинного мозга.
В ядрах ретикулярной формации, по-видимому, проецируются верхние, латеральные и нижние вестибулярные ядра. Обнаружено, что в составе вестибулярного нерва проходят миелиновые волокна, связанные с мозговым стволом и нек-рыми ядрами мозжечка; их основной источник — промежуточное ядро среднего мозга и ядро заднего продольного пучка. От подкорковых образований начинаются волокна нейронов третьего порядка, заканчивающиеся в коре головного мозга (рис., Б).
Локализация коркового отдела В. а. изучена в основном на кошках путем регистрации вызванных потенциалов. Изучение В. а. человека показывает, что корковый отдел В. а. включает поле 21 по К. Бродманну, верхнюю теменную дольку по О. Ферстеру (1936), кору верхней височной извилины по У. Пенфилду (1957) и постцентральную извилину по Корнхуберу (рис., В).
Афферентные пути от вестибулярных рецепторов к коре изучены недостаточно.
Важно, что на нейронах вестибулярных ядер промежуточных центров и в корковой вестибулярной зоне осуществляется конвергенция импульсов, приходящих по различным афферентным волокнам.
Многочисленные афферентные и эфферентные связи Вестибулярного анализатора с различными образованиями ц. н. с. способствуют генерализации эффекта от вестибулярных раздражений. У высокоорганизованных животных и человека пространственный анализ дополняется зрительным, проприоцептивным, тактильным, слуховым анализаторами при главенствующей роли коры головного мозга как интегратора афферентной импульсации. При дефиците сенсорной информации или при чрезмерных вестибулярных раздражениях возможна вестибуло-сенсорная реакция, выражающаяся в потере пространственной ориентации (возникновение головокружения, иллюзорных ощущений).
При перевозбуждении В. а. возникает комплекс вестибуло-вегетативных расстройств, выражающихся повышением тонуса, гл. обр. парасимпатической нервной системы: урежение пульса, снижение кровяного давления, изменения ЭКГ, сужение зрачка, усиление потоотделения, перистальтики жел.-киш. тракта и др. Существенную роль в происхождении вегетативных расстройств играет ретикулярная формация мозгового ствола, с к-рой вестибулярные ядра имеют тесные структурно-функциональные взаимоотношения. Кроме того, необходимо учитывать тесные связи различных ядер В. а. с внутренними органами (В. С. Райцес, А. М. Дутов, 1971). Вестибуло-вегетативные рефлексы могут составлять основу симптомокомплекса болезни передвижения.
Методы оценки функционального состояния В. а. основаны на анализе реакций, вызванных адекватными дозированными раздражениями. Для этого существуют специальные вращающиеся кресла, центрифуги, установки, имитирующие пониженную опору, лифты. Практикуется также использование неадекватных раздражителей (электрический ток, калорический метод — см. Вестибулометрия). Путем повторных воздействий на В. а. достигается привыкание к раздражителям, или тренировка (см. Вестибулярная тренировка), физиологический смысл к-рой сводится к снижению порогов вестибуло-спинальных рефлексов и торможению сенсорных и вегетативных проявлений. Возможно, что патогенетическую основу привыкания составляет иной характер распространения возбуждения в коре головного мозга и изменение корково-нодкорковых взаимоотношений. В. а. функционирует на основе тесного взаимодействия с другими системами мозга и анализаторами: мозжечком, стрио-паллидарной системой, зрительным анализатором и т. д.
Изучение В. а. приобретает особое значение в практике освоения космоса, когда в результате воздействия невесомости и нарушения определенных видов чувствительности создаются условия, временно меняющие физиологические свойства всех анализаторов, определяющих положение тела в пространстве.
Биофизические механизмы ориентационной информации
Рецептор гравитации животных и человека всегда функционирует по одной и той же схеме, к-рая включает в себя три основных элемента. Первый — упоминавшаяся пробная масса; второй — отходящие от вершины рецепторных клеток способные к отклонению чувствительные образования (киноцилии и стереоцилии); третий элемент — рецепторная клетка, оценивающая изменения чувствительных образований и передающая в закодированном виде информацию ц. н. с. Благодаря особому расположению чувствительных образований на вершине рецепторных клеток при движении пробной массы одни клетки возбуждаются, другие тормозятся. Структурная и функциональная организация рецептора гравитации позволяет ц. и. с. определять ориентацию и осуществлять установку тела животных и человека.
Нарушение функции вестибулярного анализатора при лучевом воздействии. Чувствительность и реактивность В. а. в условиях острого облучения организма в различных дозах исследованы при использовании адекватного раздражителя (углового ускорения и ускорения Кориолиса). Установлено, что изменения в функции лабиринта отмечаются уже при дозах 50—100 рад. При этих дозах, как правило, в первые часы по-еле облучения повышается возбудимость анализатора. Облучение в больших дозах (500 и более рад) приводит к значительному первоначальному угнетению функции лабиринта, что выражается в снижении возбудимости и реактивности В. а. При действии раздражителя большей силы развиваются парадоксальные и ультрапарадоксальные реакции. Через 5 и более часов после облучения происходит нормализация деятельности анализатора. В условиях хронического облучения организма животных в течение 3—6 лет при суммарной дозе 50 рад/год функция вестибулярного анализатора существенно не нарушается, ответные реакции адекватны величине примененного раздражителя; аналогичные результаты были получены и в клинике.
В экспериментах с локальным облучением живота, лицевой части головы или только головного мозга было показано, что в возникновении функциональных нарушений со стороны В. а. большую роль играют так наз. отраженные эффекты. Облучение периферических частей тела также приводит к опосредованному действию ионизирующей радиации на В. а., что выражается в угнетении его деятельности.
Используя рекомендуемые средства профилактики лучевой болезни (см. Радиопротекторы), можно ослабить функциональные нарушения со стороны Вестибулярного анализатора.
На основании проведенных клинико-экспериментальных исследований сделан вывод, что при особых условиях комбинированного действия излучения и различных физ. факторов окружающей среды (вибрация, шумы и др.) возможно сопряженное повышение реактивности В. а.
Нарушение функции В. а. при его облучении может наблюдаться при облучении среднего уха в терапевтических целях, у работающих в определенных профессиональных условиях (напр., рентгенологи), а также у космонавтов при длительных космических полетах.
Нарушение функции вестибулярного анализатора при других патологических состояниях — см. Вестибулярный симптомокомплекс, Лабиринтит, Лабиринтопатия, Меньера болезнь, Укачивание.
Библиография: Винников Я. А. Цитологические и молекулярные основы рецепции, Л., 1971, библиогр.; Горгиладзе Г. И. Корковые механизмы вестибулярной функции, Усп. совр, биол., т. 70, в. 1, с. 65, 1970, библиогр.; Григорьев Ю. Г., Фарбер Ю. В. и Волохова Н. А. Вестибулярные реакции (Методы исследования и влияния различных факторов внешней среды), М., 1970, библиогр.; Курашвили А. Е. и Бабияк В. И. Физиологические функции вестибулярной системы, Л., 19 75, библиогр.; Парин В. В. и др. Очерки по космической физиологии, М., 1967, библиогр.; Разумеев А. Н. и Шипов А. А. Нервные механизмы вестибулярных реакций, М., 1969, библиогр.; Рецептор гравитации, эволюция структурной, цитохимической и функциональной организации, под ред. В. Н. Черниговского, Л., 1971, библиогр.; Хилов К. Л. Кора головного мозга в функции вестибулярного анализатора, М.— Л., 1952, библиогр.; он же, Функция органа равновесия и болезнь передвижения, Л., 1969, библиогр.; Циммерман Г. С. Ухо и мозг, М., 19 74; Handbook of sensory physiology, ed. by H. Autrum a. o., v. 1, В. a. o., 1971; Penfield W. Vestibular sensation and the cerebral cortex, Ann. Otol. (St. Louis), v. 66, p. 691, 1957.
Ф. П. Ведяев, М. Д. Емельянов. Я. А. Винников, К. А. Койчев (биофиз.), Ю. Г. Григорьев (рад.)
Вестибулярный ганглий представлен верхней и нижней группой клеток. Верхняя группа клеток иннервирует гребешки переднего и наружного полукружных протоков, пятно эллиптического мешочка и передний верхний участок пятна сферического мешочка. Нижняя группа клеток иннервирует большую часть пятна сферического мешочка и заднего полукружного протока. Из верхней части вестибулярного ганглия начинаются крупные волокна, а из нижней части - мелкие. Верхняя группа клеток формирует верхний преддверный нерв, нижняя группа - нижнюю порцию преддверного нерва.
Вестибулярный ганглий залегает на дне внутреннего слухового прохода. Центральные отростки его клеток образуют преддверный корешок VIII нерва, который во внутреннем слуховом проходе образует анастомоз с улитковым корешком VIII нерва. В области мостомозжечкового угла преддверно-улитковый нерв внедряется в вещество мозга (мост).
В области моста волокна вестибулярного нерва делятся на восходящие, которые оканчиваются на нейронах верхних вестибулярных ядер или в мозжечке, а также нисходящие волокна, завершающие свой путь в нижней группе вестибулярных ядер.
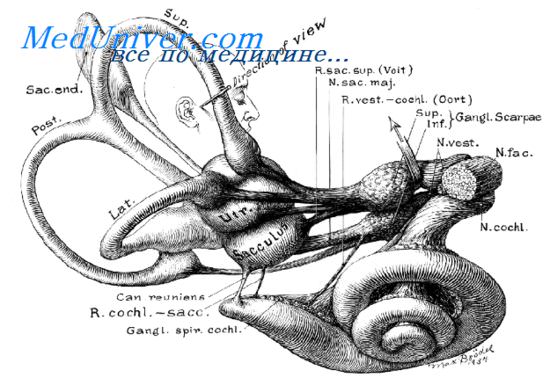
Все ядра расположены в вестибулярном поле на дне IV желудочка на границе между мостом и продолговатым мозгом. Выделяют четыре вестибулярных ядра: верхнее (ядро Бехтерева), медиальное (ядро Швальбе), латеральное (ядро Дейтерса) и нижнее (ядро Роллера). Ядра формируют регион расположения II нейрона.
В медиальном ядре заканчиваются афферентные волокна гребешков полукружных протоков и мозжечка. В наружную часть ядра поступают волокна от пятна эллиптического мешочка и от ретикулярной формации. К нижнему ядру подходят афферентные волокна от мозжечка, пятен обоих мешочков и волокна от гребешков.
Клетки, окружающие вестибулярные ядра, формируют несколько трактов. Эти пути направляются в спинной мозг, кору головного мозга, мозжечок в составе медиального и заднего продольного пучков. Путь от лабиринта к коре головного мозга переключается в ядрах таламуса (область Ш нейрона) и оканчивается в височно-теменной области в зонах Пен-филда и Форстера (регион IV нейрона).
Вестибулярные ядра связаны с ретикулярной формацией, глазодвигательными ядрами, с моторной частью спинного мозга, а также с мозжечком, вегетативной нервной системой и височными долями больших полушарий мозга.
Благодаря обширным связям вестибулярных ядер осуществляются различные рефлексы. Выделяют кинетические рефлексы, которые контролируют координированное сокращение мышц и движение глаз при перемещении. Эта группа рефлексов инициируются активностью пятен и гребешков на линейные и угловые ускорения. А также рефлексы, направленные на поддержание позы и мышечного тонуса, они возникают при активности статокинетических пятен и служат компенсаторным ответом на действие гравитационных сил. Рефлекторная дуга включает: рецептор, первичный нейрон, нейрон II порядка (ядра), нейрон Ш порядка (мотонейроны) и эффектор (отвечающий орган).
Вестибулярный анализатор включает периферической аппарат, нервные проводящие пути, центральные подкорковые и корковые ядерные образования, комплекс ассоциативных связей, осуществляющих взаимодействие вестибулярного анализатора с соматической и вегетативной нервной системой, а также с другими анализаторами.
Периферическая часть вестибулярного анализатора
Периферическая часть вестибулярного анализатора представлена симметрично расположенной системой перепончатых образований, состоящих из двух пар мешочков преддверия и трех пар полукружных протоков, заключенных в соответствующие костные каналы (рис. 1).
Рис. 1. Анатомо-топографическое положение улитки и вестибулярного аппарата (по Waever Е., 1965): 1, 2, 3 — соответственно боковой, задний и передний полукружные каналы; 4, 5 — соответственно эллиптический и сферический мешочки; 6, 7 — верхняя и нижняя части преддверного ганглия; 8 — вестибулярный нерв; 9 — лицевой нерв; 10 — улитковый нерв; 11 — эндолимфатический проток; 12 — эндолимфатический мешок; 13 — общий гладкий конец переднего и заднего полукружных каналов; 14 — улитка; 15 — соединительный проток
Полости мешочков преддверия и полукружных протоков входят в единую замкнутую эндолимфатическую систему ушного лабиринта височной кости, окруженную перилимфой.
В мешочках преддверия и в полукружных протоках расположены специальным образом ориентированные рецепторы, реагирующие на механическое смещение инерционных масс вестибулярного аппарата (эндолимфы полукружных протоков и отолитовых мембран мешочков преддверия).
Периферическая часть вестибулярного анализатора (см. рис. 1) состоит из преддверия (4, 5) и трех костных полукружных каналов (canales semicirculares (1, 2, 3)). Последние ориентированы таким образом, что при вращении головы в любой плоскости всегда возникает наиболее функционально активная пара каналов, которая определяет вектор возникающих ответных сенсорных и двигательных реакций. Гравирецепторная система преддверия (4, 5), представленная отолитовым аппаратом, ориентирована так, что его рецепторные образования реагируют на любые виды прямолинейного движения, а также на вектор силы земного притяжения и изменение его направления.
Отолитовый аппарат состоит из эллиптического (4) и сферического мешочков (5), а также трех полукружных протоков — бокового, или горизонтального, заднего, или сагиттального (2), и переднего (верхнего), или фронтального (3). Каждый из каналов имеет два конца — один булавовидный (ампула полукружного протока), другой гладкий. Гладкие концы переднего и заднего каналов объединяются в один общий проток и вступают в эллиптический мешочек (13). Этот мешочек сообщается с улиткой (14) посредством капиллярного соединительного протока (15), роль которого заключается в пропуске эндолимфы, продуцируемой сосудистой полоской улитки, в вестибулярные эндолимфатические полости.
Вестибулярные рецепторы относятся к механорецепторам, воспринимающим механическую энергию, развиваемую силами инерции и гравитации. Различают два типа рецепторных клеток вестибулярного аппарата (рис. 2). Клетки первого типа (I) имеют форму колбы и являются в эволюционном отношении более молодыми; клетки второго типа (II) — цилиндрической формы и в эволюционном отношении более древние.
Рис. 2. Вестибулярные рецепторные клетки: I — клетка первого типа; II — клетка второго типа; К — киноцилий; С — стереоцилии; Э — эфферентные связи; А — афферентные связи
Рецепторные клетки связаны с нейронами, центральные отростки которого (аксоны) образуют вестибулярную порцию преддверно-улиткового нерва.
Указанный поляризационный принцип строения волоскового аппарата характерен не только для рецепторов полукружных каналов, но и для клеток утрикулюса и саккулюса (эллиптического и сферического мешочков). Утрикулюс расположен в горизонтальной плоскости, саккулюс — в сагиттальной плоскости. Между ними существует сообщение в виде узкого протока.
Концы волосков рецепторных клеток утрикулюса и саккулюса приходят в соприкосновение с так называемой отолитовой, или статокониевой, мембраной (рис. 3, 2), состоящей из мукополисахаридного геля с включенными в него беспорядочно переплетающимися волокнами и расположенными среди них микроскопическими кристаллами кальция (1), размер которых составляет от 0,5 до 30 мкм. Удельный вес этих кристаллов 2,93-2,95, что приблизительно в 3 раза больше плотности эндолимфы, в которой они находятся. Отолитовая мембрана играет роль инерциальной и гравитационной массы для рецепторов отолитового аппарата (8).
Рис. 3. Строение отолитового аппарата: 1 — микроскопические кристаллы углекислого кальция; 2 — отолитовая мембрана; 3 — кино-цилий; 4 — рецептивное поле (пятно); 5 — дендрит вестибулярного узла; 6 — вестибулярный узел; 7 — вестибулярная порция преддверно-улиткового нерва (пучок аксонов вестибулярного узла); 8 — рецепторная клетка; 9 — стереоцилии
Иннервация вестибулярного аппарата
Волокна вестибулярного нерва берут начало в преддверном узле, расположенном в глубине внутреннего слухового прохода (см. рис. 1, 6, 7). Дендриты клеток этих узлов следуют в периферическом направлении через отверстия костного дна внутреннего слухового прохода, подходят к рецепторам ампулярных гребешков полукружных протоков и пятен отолитового аппарата. Аксоны клеток преддверного узла составляют преддверную часть VIII нерва, которая по выходе из узла соединяется с улитковым нервом и вместе с ним образует преддверно-улитковый нерв, следующий в центральном направлении. Во внутреннем слуховом проходе преддверно-улитковый нерв присоединяется к лицевому нерву и вместе с ним покидает пирамиду через внутреннее слуховое отверстие и проникает в полость черепа, далее — в толщу дна ромбовидной ямки продолговатого мозга, где вступает в вестибулярные ядра — верхнее, нижнее, медиальное и латеральное. Вестибулярные ядра посредством проводящих путей связаны с мозжечком, ядрами глазодвигательных нервов, ретикулярной формацией, двигательными нейронами спинного мозга, зрительным бугром, височными долями головного мозга и др.
Кровоснабжение и лимфоотток из органов преддверия осуществляются той же сосудистой системой, что и улитка.
Клинико-анатомическое значение центральных образований вестибулярной системы заключается в том, что при диагностике некоторых отоневрологических синдромов, связанных с поражением ствола головного мозга, возникающие соматомоторные нарушения следует оценивать с учетом приведенных сведений о связях вестибулярных ядер с соответствующими двигательными ядрами ствола головного и спинного мозга.
Оториноларингология. В.И. Бабияк, М.И. Говорун, Я.А. Накатис, А.Н. Пащинин
Общая характеристика – чувствительный, сознательный, 3-х нейронный, перекрещенный.
I нейрон – биполярные клетки преддверного ганглия. Их дендриты заканчиваются на волосковых сенсорных (нейросенсорных) клетках ампулярных гребешков и пятен. Аксоны формируют преддверную часть преддверно-улиткового нерва, в области мостомозжечкового угла они входят в мост, где переключаются на тела II нейронов.
II нейроны – клетки медиального (Швальбе), верхнего (Бехтерева), латерального (Дейтерса) и нижнего (Роллера) вестибулярных ядер. АксоныII нейронов переходят на противоположную сторону, присоединяются к проводникам медиальной петли, в составе которой достигают тела III нейронов.
III нейроны –клетки латерального ядра зрительного бугра. Их аксоны в составе таламокортикального пути через заднюю ножку внутренней капсулы приходят в кору верхней височной извилины.
Часть аксонов II нейронов от ядер Дейтерса и Роллера идут в нисходящем направлении по своей стороне (в составе нижней ножки мозжечка) в качестве экстрапирамидного преддверно-спинномозгового пути.
Часть проводников вестибулярного пути направляется в кору мозжечка, обеспечивая его участие в поддержании равновесия.

Слуховой проводящий путь

Вестибулярный проводящий путь
Тестовые задания
1. К наружному уху относится
а. барабанная полость
б. наружный слуховой проход
в. слуховая труба
г. мышца стремечка
2. К наружному уху относятся
а. окно преддверья
б. вторичная барабанная перепонка
в. барабанная перепонка
3. К наружному уху относятся
а. ушная раковина
К элементам барабанной перепонки не относится
а. напряженная часть
б. расслабленная часть
в. промежуточная часть
К слоям барабанной перепонки не относится
Соотношение хрящевой и костной частей наружного слухового прохода
6. Барабанная перепонка относится
а. к среднему уху
б. к наружному уху
в. к внутреннему уху
К образованиям среднего уха не относится
а. мышца стремячка
б. слуховая труба
в. барабанная перепонка
г. надбарабанный карман
8. Образования среднего уха
а. мышца, напрягающая барабанную перепонку
б. костный лабиринт
г. перепончатый лабиринт
9. Барабанная полость сообщается
а. с полостью носа.
б. с пещерой и ячейками сосцевидного отростка
в. с полостью ротоглотки
г. с полостью гортаноглотки
Барабанная полость посредством слуховой трубы сообщается
а. с наружным слуховым проходом
б. с внутренним слуховым проходом
в. с полостью носа
г. с полостью носоглотки
11. Содержимое барабанной полости
г. цереброспинальная жидкость
Передней стенкой барабанной полости является
а. яремная стенка
б. лабиринтная стенка
в. сонная стенка
г. сосцевидная стенка
13. Задней стенкой барабанной полости является
а. перепончатая стенка
б. сосцевидная стенка
в. яремная стенка
г. сонная стенка
14. Нижней стенкой барабанной полости является
а. сосцевидная стенка
б. покрышечная стенка
в. лабиринтная стенка
г. яремная стенка
15. Верхней стенкой барабанной полости является
а. сосцевидная стенка
б. перепончатая стенка
в. покрышечная стенка
г. сонная стенка
16. Медиальной стенкой барабанной полости является
а. лабиринтная стенка
б. покрышечная стенка
в. сонная стенка
г. сосцевидная стенка
17. Латеральной стенкой барабанной полости является
а. сосцевидная стенка
б. перепончатая стенка
в. покрышечная стенка
г. лабиринтная стенка
18. К слуховым косточкам не относится
19. В окне преддверия (овальном окне) располагается
б. барабанная перепонка
г. вторичная барабанная перепонка
20. В окне улитки (круглом окне) располагается
б. барабанная перепонка
г. вторичная барабанная перепонка
21. Мышцы среднего уха
б. мышца, напрягающая барабанную перепонку
22. Мышцы среднего уха
а. мышца стремечка
23. Образования костного лабиринта
б. слуховая труба
г. пещера сосцевидного отростка
24. Образования костного лабиринта
б. маточково-мешочковый проток
в. слуховая труба
г. соединительный проток
25. Образования костного лабиринта
а. костная часть наружного слухового протока
в. полукружные протоки
г. полукружные каналы
26. Образования улитки
а. спиральная пластинка
б. эллиптический карман
в. сферический карман
г. полукружные каналы
27. Образования улитки
в. сферический карман
г. спиральный канал
28. Образования полукружных каналов
а. окно преддверия ( овальное)
б. простая ножка
в. эллиптический карман
29. Образования полукружных каналов
а. пещера сосцевидного отростка
б. надбарабанный карман
в. эллиптический карман
г. ампулярная ножка
30. Полукружные каналы представлены
а. передним полукружным каналом
б. внутренним полукружным каналом
в. нижним полукружным каналом
31. Полукружные каналы представлены
а. нижним полукружным каналом
б. внутренним полукружным каналом
в. латеральным полукружным каналом
32. Части перепончатого лабиринта
а. сферический карман
б. эллиптический карман
г. улитковый канал
33. Части перепончатого лабиринта
а. простые ножки полукружных каналов
б. ампулярные ножки полукружных каналов
в. улитковый канал
34. Части перепончатого лабиринта
а. мышца, напрягающая барабанную перепонку
б. мышца стремечка
в. вторичная барабанная перепонка
35. Части перепончатого лабиринта
а. соединительный проток ( ductus reuniens)
б. мышца стремечка
в. вторичная барабанная перепонка
г. полукружные каналы
36. Соединительный проток (ductus reuniens) соединяет
а. полукружные протоки с маточкой
б. маточку с мешочком
в. мешочек с улитковым протоком
г. маточку с улитковым протоком
37. Улитковый проток разделяет полость улитки на
а. лестницу преддверия
б. полукружные протоки
в. эллиптический карман
г. сферический карман
38. Улитковый проток разделяет полость улитки на
а. барабанную лестницу
б. полукружные протоки
39. Полость костного лабиринта заполнена
г. цереброспинальной жидкостью
40. Полость перепончатого лабиринта заполнена
г. цереброспинальной жидкостью
41. Элементы улиткового протока
а. ампулярные гребешки
б. пятно маточки
в. спиральный (кортиев) орган
г. пятно мешочка
42. Элементы улиткового протока
а. спиральная мембрана (основная пластинка)
б. пятно маточки
в. ампулярные гребешки
г. пятно мешочка
43. Спиральный (кортиев) орган располагается
а. в полукружных каналах
44. Слуховой путь является
г. 5-ти нейронным
45. Слуховой путь является
а. дважды перекрещенный
б. частично перекрещенным
46. I нейроны слухового пути представлены
а. униполярными клетками
б. псевдоуниполярным клетками
в. биполярным клетками
г. мультиполярным клетками
47. I нейроны слухового пути располагаются в
а. преддверном ганглии
б. спиральном ганглии
в. в чувствительном ганглии V пары
г. в чувствительном ганглии VII пары
48. Спиральный ганглий располагается
а. в наружном слуховом проходе
б. во внутреннем слуховом проходе
в. в кортиевом органе
г. в спиральном канале улитки
49. II нейроны слухового пути располагаются в
а. в продолговатом мозге
в. в среднем мозге
г. в промежуточном мозге
50. II нейронами слухового пути является
а. собственные ядра моста
б. вентральное улитковое ядро
в. верхнее слюноотделительное ядро
г. ядра крыши верхних холмиков среднего мозга
51. II нейронами слухового пути является
б. ядро Дейтерса
в. дорзальное улитковое ядро
г. ядро Бехтерева
52. Перекрест слухового пути представлен
а. пирамидным перекрестом
б. слуховыми (мозговыми) полосками
в. перекрестом петель
г. вентральным перекрестом Фореля
53. Перекрест слухового пути представлен
а. посегментным перекрестом
б. надядерным перекрестом
в. перекрестом петель
г. трапециевидным телом
54. Аксоны II нейронов слухового пути после перекреста образуют
а. медиальную петлю
б. латеральную петлю
в. мостомозжечковый путь
г. трапециевидное тело
55. III нейроны сознательной порции слухового пути залегают в
а. верхних холмиках среднего мозга
б. нижних холмиках среднего мозга
в. латеральном коленчатом теле
г. медиальном коленчатом теле
56. III нейроны бессознательной порции слухового пути залегают в
а. верхних холмиках среднего мозга
б. нижних холмиках среднего мозга
в. латеральном коленчатом теле
г. медиальном коленчатом теле
57. Корковый конец слухового анализатора расположен в
а. надкраевой извилине
б. покрышечной части
в. верхней височной извилине (извилине Гешля)
г. угловой извилине
58. Эфферентным звеном бессознательной порции слухового пути является
а. покрышечно-спинномозговой путь
б. красноядерно-спинномозговой путь
в. пирамидные пути
г. преддверно-спинномозговой путь
59. Вестибулярный путь является
г. 5-ти нейронным
60. Вестибулярный путь является
а. дважды перекрещенный
б. частично перекрещенным
61. Рецепторы вестибулярного пути залегают
а. в полукружных протоках
б. в ампулах полукружных протоков
в. в улитковом канале
г. в улитковом протоке
62. Рецепторы вестибулярного пути залегают
а. во внутреннем слуховом проходе
б. в улитковом протоке
в. в наружном слуховом проходе
г. в маточке и мешочке
63. Вестибулярный путь является
в. дважды перекрещенным
г. частично перекрещенным
64. I нейроны вестибулярного пути представлены
а. униполярными клетками
б. псевдоуниполярным клетками
в. биполярным клетками
г. мультиполярным клетками
65. I нейроны вестибулярного пути располагаются в
а. преддверном ганглии
б. спиральном ганглии
в. в чувствительном ганглии V пары
г. в чувствительном ганглии VII пары
66. Вестибулярный ганглий располагается
а. в наружном слуховом проходе
б. во внутреннем слуховом проходе
в. в кортиевом органе
г. в спиральном канале улитки
67. Аксоны I нейронов вестибулярного пути проходят в составе
а. V пары черепных нервов
б. VI пары черепных нервов
в. VII пары черепных нервов
г. VIII пары черепных нервов
68. II нейроны вестибулярного пути располагаются в
а. в продолговатом мозге
в. в среднем мозге
г. в промежуточном мозге
69. II нейронами вестибулярного пути является
а. ядро Якубовича
г. ядра крыши верхних холмиков среднего мозга
70. II нейронами вестибулярного пути является
а. дорзальное улитковое ядро
б. вентральное улитковое ядро
в. ядро Дейтерса
г. латеральное коленчатое тело
71. II нейронами вестибулярного пути является
а. собственные ядра моста
б. ядро одиночного пути
г. ядро Бехтерева
72. Перекрест вестибулярного пути происходит на уровне
а. продолговатого мозга
в. среднего мозга
г. промежуточного мозга
73. Аксоны II нейронов вестибулярного пути после перекреста входят в состав
а. медиальной петли
б. латеральной петли
в. мостомозжечкового пути
г. корково-мостового пути
74. III нейроны сознательной порции вестибулярного пути залегают в
а. латеральном ядре зрительного бугра
б. в подушке зрительного бугра
в. латеральном коленчатом теле
г. медиальном коленчатом теле
75. Корковый конец вестибулярного анализатора расположен в
а. надкраевой извилине
б. покрышечной части
в. верхней височной извилине (извилине Гешля)
г. угловой извилине
76. I нейронами бессознательной порции вестибулярного пути являются
б. ядро Дейтерса
в. ядро Бехтерева
г. ядро Якубовича
77. Эфферентным звеном бессознательной порции вестибулярного пути является
Читайте также: