
Высшая нервная деятельность рыбы
ИЗУЧЕНИЕ ПОВЕДЕНИЯ И АДАПТАЦИИ РЫБ К ВНЕШНИМ УСЛОВИЯМ
Изучение поведения рыб - одна из важнейших задач ихтиологии и безмерное поле проведения интереснейших и увлекательнейших экспериментов и исследований. В частности, сохранение запасов ценных проходных и полупроходных рыб в связи с гидростроительством невозможно без успешного изучения поведения этих рыб на нерестилищах, в зоне плотин и рыбопропускных сооружений. Не менее важно предотвращение засасывания рыб в водозаборные сооружения. В этих целях уже используются или прошли испытания такие устройства, как пузырьковая завеса, электрорыбозаградители, механические решетки и т. д., но пока применяемые устройства не достаточно эффективны и экономичны.
Для успешного развития промысла и совершенствования орудий лова крайне важны сведения о поведении рыб в зоне облова, зависимости от гидрометеобстановки и гидрологических факторов, о суточных и периодических вертикальных и горизонтальных миграциях. При этом рациональная организация промысла не возможна без изучения распределения и поведения разновозрастных групп. Сроки и мощность миграций, подходы рыб на места нереста, нагула, зимовки во многом определяются изменениями условий внешней среды и физиологического состояния особей.
Значение органов чувств в восприятии абиотических и биотических сигналов
Изучение поведения рыб проводится на основании регулярных натурных наблюдений, экспериментов в лабораторных условиях и анализа данных о взаимодействии с внешней средой высшей нервной деятельности изучаемых объектов. В процессе взаимодействия с окружающей средой у рыб проявляются три способа ориентации:
- пеленгация - воспроизведение сигнала, идущего от внешнего мира;
- локация - посылка сигналов и восприятие их отражений;
- сигнализация - посылка сигнала одними особями и восприятие их другими.
Восприятие абиотических и биотических сигналов, влияющих на поведение рыб, происходит посредством органов чувств, среди которых выделяют прежде всего зрение, слух, боковую линию, обоняние. Особое значение имеет рефлекторная деятельность рыб.
Зрение рыб
По сравнению с воздушной средой вода, как среда обитания рыб, менее благоприятна для зрительного восприятия. Освещенность водных слоев проникающими в воду солнечными лучами находится в прямой зависимости от количества растворенных и взвешенных частиц, которые обуславливают мутность воды, определяют границы действия органов зрения рыб. В морской воде освещенность достигает глубины 200-300 м, а в пресных водоемах лишь 3-10 м. Чем глубже в воду проникает свет, тем глубже проникают и растения. Прозрачность воды чрезвычайно различна. Она больше вдали от берегов и уменьшается во внутренний морях. Чем больше в воде живых организмов, тем менее вода прозрачна. Очень прозрачные воды морей, особенно красивого насыщенного синего цвета, - это воды, скудные жизнью. Самые прозрачные моря - Саргассово и Средиземное.
Рыбы обладают цветным зрением. Для особей, обитающих в освещенной зоне, оно имеет очень большое значение и обусловливает их поведение. Питание планктонофагов, в том числе молоди рыб, осуществляется благодаря хорошо развитым органам зрения. Присущая рыбам острота зрения позволяет, в зависимости от освещенности и прозрачности воды, различать предметы на расстоянии до нескольких десятков метров. Все вышеперечисленное имеет большое значение для пищевых и оборонительных реакций рыбы. Доказано, что образование и распад стай также связаны с освещенностью водной среды.
Движение рыб против течения контролируется органами зрения, реже органами обоняния. На этом основаны попытки направить рыб в рыбоходах вслед за макетами. Сосвещенностью связаны ритмы и активность питания.
Явление вертикальной зональности и преобладающей окраски животных и растений обусловлено неравномерностью проникновения лучей разной длины волны в толщу воды. Животные очень часто бывают окрашены в цвет той части спектра, которая проникает на данную глубину, в результате чего приобретает защитную окраску, кажутся незаметными. В верхних горизонтах животные большей частью окрашены в буровато-зеленоватые цвета, а глубже - в красные. На больших глубинах, лишенных света, животные большей частью окрашены в черный цвет или совсем лишены окраски (депигментированы).
Слух.
Акустические свойства воды значительно сильнее, чем воздушной среды. Звуковые колебания идут быстрее и проникают дальше. Установлено, что роль звуковой сигнализации увеличивается с наступлением сумерек, по мере уменьшения зрительного восприятия. Центр звукового восприятия - внутреннее ухо рыб. Восприятие ультразвуковых колебаний рыбам не свойственно, но зато они реагируют на низкочастотные звуки. Реакция на ультразвук обнаруживается только при действии мощного источника на небольшом расстоянии и скорее может быть отнесена к болевому ощущению кожи.
При имеющей место реакции на звуковые сигналы, рыбы направленно (рефлекторно) реагируют, прежде всего, на пищевые раздражители или сигнал опасности. В черте города рыбы довольно быстро привыкают к шумам, даже к постоянным очень громким звукам. Возможно поэтому с помощью звуковых сигналов не удалось организовать направленное движение лососей в реки или отпугнуть от сточных вод. Даже вблизи аэродромов рыбы не меняют поведения и продолжают клевать на удочку. Отмечено, что прерывистый звук действует на рыб сильнее, чем постоянный.
Боковая линия
В первую очередь следует отметить функциональную связь боковой линии с органами слуха. Установлено, что нижняя часть звуковых колебаний (частоты 1-25 Гц) воспринимаются боковой линией. Значение боковой линии изучено далеко не полностью. Основной функцией боковой линии является восприятие гидродинамических полей и струй воды. Гидродинамические поля от больших источников, вызывающих у рыб оборонительную реакцию, обычно воспринимаются на значительном расстоянии. Однако в районах образования быстрых течений в реках ниже плотины многие рыбы быстро привыкают к изменившимся условиям.
Гидродинамические поля, вызываемые от движения мелких тел, обычно вызывают у рыб пищевую реакцию. Рыбы с помощью боковой линии точно ориентируются для прицельного броска на сравнительно небольшое расстояние в несколько десятков сантиметров.
С помощью боковой линии сумеречные, ночные и зарослевые хищники ориентируются, достигая добычу. Молоди рыб и планктонофагам боковая линия служит для обнаружения хищника и общей ориентации в среде.
Обоняние рыб
Следует учесть свойство воды как хорошего растворителя. Установлено, что рыбы реагируют на ничтожно малые количества растворенных в воде веществ. С помощью запахов рыбаки привлекают рыбу. В то же время другие вещества, например настойка кожи хищных рыб и морских млекопитающих, действуют отпугивающее.
Восприятие растворенных в воде веществ, по-видимому, связано с органами вкуса. Проходные рыбы находят путь из моря в реки с помощью органов обоняния. Несомненно, что рыбы способны к запоминанию. Этим объясняется хоминг (от англ. home - ≪дом≫) - способность рыб заходить именно в те реки, протоки или гирла, из которых они вышли мальками после развития из икры.
Высшая нервная деятельность и поведение рыб
Способность рыб приобретать условные рефлексы в сочетании с безусловными рефлексами дает возможность управлять их поведением. Условные рефлексы вырабатываются у рыб медленнее, чем у высших позвоночных, и быстро угасают, если не находят подкрепления теми же факторами, которые способствовали их образованию, но способны самопроизвольно возникать через определенное время.
Особую роль в создании и угасании рефлексов играет температура воды. Имеются данные (Юдкин, 1970), что у осетровых условные рефлексы осенью вырабатываются значительно хуже, чем летом. У золотой рыбки понижение температуры воды ниже +13 °С и повышение свыше +30 °С вызывало исчезновение всех ранее приобретенных рефлексов. Все это становится вполне понятно, если учесть, что жизнедеятельность рыб, животных с пониженной температурой крови, зависит от температуры воды.
Условные рефлексы могут возникать у рыб в виде подражания. Необученные рыбы подражают другим, у которых условные рефлексы образовались после соответствующего обучения или приобретения жизненного опыта. Весьма показательно в этом отношении изменение поведения рыб в зоне обловов активных и даже стационарных орудий лова. Часто достаточно одной особи, обнаружившей лазейку для выхода из орудия лова, чтобы его покинула большая часть стаи (например, хамса в ставных и закидных неводах).
Пиленгас способен преодолевать сетные порядки, переваливаясь через верхнюю подбору, выпрыгивать и даже ползти, извиваясь по наклонному полотну при выборке закидных неводов.
Летчики-наблюдатели, длительное время занимавшиеся наводкой промысловых судов на косяки рыбы, отмечали постепенное изменение поведения хамсы: изменение направления движения и выход из кошельковых неводов, ≪приседание≫, рассеивание и т.д.
Не идентичны поведение и быстрота реакций рыб при разном физиологическом состоянии. Жирная рыба быстрее образует скопления, которые усидчивее, чем образованные физиологически ослабленными особями. Часто рыбы реагируют не только на резкие изменения условий, но и на складывающиеся тенденции изменения факторов среды. При слабом же росте температуры воды скопления могут просто распасться, несмотря на то, что температура будет оставаться в пределах оптимальной для ведения промысла.
Большое значение имеет формирование рыб в стаи. Оборонительное значение стаи у рыб так же велико, как и у птиц. Также, охватывая большее водное пространство, стая быстрее находит места откорма, чем отдельные особи.
Наблюдения показали у некоторых видов рыб наличие вертикальных миграций. Так, на ньюфаундлендской банке морской окунь с заходом солнца в течение 60-90 мин поднимается с глубин 500-600 м на глубины 300-400 м. Ночью окунь держится в 200 м от поверхности, а к утру опускается и днем находится у дна. Подобным образом ведут себя треска и пикша. В Черном море вертикальные миграции наиболее свойственны хамсе и ставриде, опускающимся в нижние горизонты в дневные часы и поднимающимся к поверхности в ночные. Такое поведение их связано с движениемпланктона. Для многих рыб нахождение на разной глубине и на разном удалении от берега характерно в различные периоды жизненного цикла.
Все перечисленное имеет непосредственное отношение к поведению рыб. Это необходимо учитывать исследователю для более эффективного влияния на поведение рыб в зонах облова, где необходимо выявлять ведущие факторы для каждого конкретного случая. В настоящее время знание особенностей поведения приобретает особое значение для успешного развития промысла. И связано это, прежде всего, с увеличением интенсивности рыболовства, падением запасов и ростом экономической себестоимости выполнения работ.
Изучение особенностей поведения в зависимости от факторов среды и физиологического состояния рыб позволяет исследователям и промысловикам тактически регулировать лов с повышением его эффективности. Знание биологии промыслового объекта позволяет организовать лов в периоды максимальных концентраций, на глубинах наибольшего распределения и при температурах воды, когда скопления наиболее устойчивы. Одним из инструментов таких исследований является многофакторный Коррелятивный анализ наиболее значимых связей океанологических и биологических критериев для построения математических моделей, описывающих явления и процессы жизненного цикла рыб. Достаточно давно и хорошо на ряде бассейнов зарекомендовали себя прогнозы сроков осенних миграций, образования и распада зимовальных скоплений и начала промысла массовых промысловых рыб. Это способствует сокращению непроизводительных простоев судов и повышению уровня интенсивности лова.
В качестве примеров таких моделей можно привести рассчитанные в АзНИИРХ уравнения регрессии для прогнозирования сроков осенней миграции азовской хамсы через Керченский пролив в Черное море.
У = 27,68- 0,18 Х2- 0,009 (Н).
Начало массовой миграции:
где У и У1 - даты предполагаемого начала осенней миграции и массового хода (отсчет от 1 сентября); X1 и Хз - даты окончательного перехода температуры воды через +16 и +14 °С (соответственно) в южной части Азовского моря (отсчет от 1 сентября); Х2 — количество рыб (в %) в популяции с коэффициентом упитанности 0,9 и более по состоянию на 1 сентября, Н —продолжительность нагула (градус/дни) после нереста на 1 сентября.
Ошибка прогнозов сроков начала миграций по представленным моделям не превышает 2-3 дней.
Лекция
Дата добавления: 2016-11-29 ; просмотров: 2127 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Нервная система рыб делится на периферическую и центральную. Центральная нервная система состоит из головного и спинного мозга, а периферическая – из нервных волокон и нервных клеток.
Головной мозг рыб.
Головной мозг рыб состоит из трех основных частей: передний мозг, средний мозг и задний мозг. Передний мозг состоит из конечного мозга (теленцефалона) и промежуточного мозга – диенцефалона. На переднем конце теленцефалона находятся луковицы, отвечающие за обоняние. Они принимают сигналы от обонятельных рецепторов.
Схему обонятельной цепи у рыб можно описать следующим образом: в обонятельных долях мозга есть нейроны, которые являются частью обонятельного нерва или пары нервов. Нейроны присоединяются к обонятельным участкам теленцефалона, которые также называются обонятельными долями. Обонятельные луковицы особенно выделяются у рыб, которые используют органы чувств, например, акулы, которые выживают за счет нюха.
Диенцефалон состоит из трех частей: эпиталамуса, таламуса и гипоталамуса и выполняет функции регулятора внутренней среды организма рыбы. Эпиталамус содержит шишковидный орган, который, в свою очередь состоит из нейронов и фоторецепторов. Шишковидный орган расположен на конце эпифизы и у многих видов рыб он может быть чувствительным к свету благодаря прозрачности костей черепа. Благодаря этому шишковидный орган может выполнять функцию регулятора циклов активности и их смены.

В среднем мозге рыб находятся зрительные доли и тегментум или покрышка – и то, и другое используются для обработки оптических сигналов. Зрительный нерв рыб очень разветвлен и имеет много волокон, отходящих от зрительных долей. Как и в случае с обонятельными долями, увеличенные зрительные доли можно встретить у рыб, жизнедеятельность которых зависит от зрения.
Тегментум у рыб контролирует внутренние мышцы глаза и тем самым обеспечивают его фокусирование на предмете. Также тегментум может выступать в роли регулятора функций активного контроля – именно здесь расположен локомоторный регион среднего мозга, отвечающий за ритмичные плавательные движения.
Задний мозг рыб состоит из мозжечка, вытянутого мозга и моста. Мозжечок – это непарный орган, выполняющий функцию поддержания равновесия и контроля положения тела рыбы в среде. Продолговатый мозг и мост вместе составляют ствол головного мозга, к которому тянется большое количество черепно-мозговых нервов, несущих сенсорную информацию. Большинство всех нервов связываются с головным мозгом и входят в него через ствол и задний мозг.
Спинной мозг.
Спинной мозг находится внутри нервных дуг позвонков рыбного позвоночника. В позвоночнике имеется сегментация. В каждом сегменте нейроны соединяются со спинным мозгом с помощью дорсальных корешков, а проворные нейроны выходят их через вентральные корешки. Внутри центральной нервной системы также находятся интернейроны, которые обеспечивают сообщение между проворными и сенсорными нейронами.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней — archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.40): передний (I), средний (II) и задний (III). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2) мозг — telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon, a задний — в задний (4) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Александр Борисович Коган
Основы физиологии высшей нервной деятельности
(издание второе, переработанное и дополненное)
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
Все это потребовало коренной переработки учебника, замены устаревшего материала новым, пополнения книги накопившимися за это время фактами и идеями. Особое внимание уделено главам частной физиологии высшей нервной деятельности у разных видов животных, где биолог соответствующей специальности (ихтиолог, энтомолог, герпетолог, орнитолог и др.) может получить интересующие его сведения. Основательно переработаны главы физиологии высшей нервной деятельности человека. Они содержат результаты многочисленных, продолжающих накапливаться психофизиологических и нейропсихологических исследований и могут представлять интерес не только для физиологов биологического профиля, но также для психологов, врачей и педагогов.
Конечно, и после такой переработки учебник не лишен некоторых недостатков и погрешностей. Поэтому автор будет признателен всем, кто поможет избежать их в дальнейшем своими замечаниями и советами.
Автор выражает благодарность сотрудникам Института нейрокибернетики и кафедры физиологии человека и животных Ростовского государственного университета им. М.А. Суслова: Т.В Алейниковой, М.X. Закиян, О.Ф. Маркштедер, Т.А. Амелиной, Е.В. Яковлевой, И.Ф. Калмыкову, оказавшим большую помощь в подготовке данного учебника к печати.
ПРЕДИСЛОВИЕ К ПЕРВОМУ ИЗДАНИЮ
Курс физиологии высшей нервной деятельности читается в университетах, педагогических, медицинских и сельскохозяйственных высших учебных заведениях. Однако учебника по этой дисциплине студенты не имеют. Вытекающие отсюда чрезвычайные трудности преподавания побудили к попытке составить такой учебник для биологов.
К настоящему времени накопился громадный материал по частной физиологии высшей нервной деятельности животных и человека. Собрано очень много, правда пока разрозненных, фактов изучения условных рефлексов у животных разных классов, отрядов и видов. В связи с этим встал вопрос и об использовании фактов, накопленных экспериментальной психологией, освободив их от антропоморфических толкований. Наконец, имеется уже немалое число случаев разнообразнейшего прикладного использования условных рефлексов в практике медицины, педагогики, сельского хозяйства. Все эти сведения, несомненно нужные учащемуся, введены в учебник и заняли в нем свое место.
Учебник написан для биологов. Поэтому он построен в сравнительном и, когда это было возможным, в эволюционном плане. Частная физиология высшей нервной деятельности выделена в большой самостоятельный раздел, потребовавший особенно много труда и внимания. В этом разделе мы хотели собрать для биологов разных специальностей конкретный материал описания и, по возможности, физиологического объяснения поведения изучаемых ими животных. Здесь, к примеру, энтомолог должен найти собранные и систематизированные сведения о высшей нервной деятельности насекомых, ихтиолог — о высшей нервной деятельности рыб, орнитолог — птиц и т.д.
Просмотр корректур перед выходом учебника в свет показал, что за время, прошедшее после сдачи рукописи в печать, в области физиологии высшей нервной деятельности появилось много новых фактов и обобщений. К сожалению, по техническим причинам только малую их часть удалось включить дополнительно в текст.
Полностью отдавая себе отчет в слабых местах этого учебника по новой дисциплине, автор с благодарностью примет все замечания и советы, которые ему будут даны.
В процессе тысячевековой эволюции мира животных развивалась их способность гибко адаптировать свое поведение к условиям окружающей среды. Эта способность достигла высочайшего совершенства в функциях нервной системы, особенно с развитием исключительно ценных для выживания свойств: фиксации следов пережитых событий. Такая память, в широком смысле этого слова, позволяла каждой особи действовать, исходя из собственного жизненного опыта, показывала связь событий в окружающем мире, а при формировании мозга человека явилась основой механизмов его мыслительной деятельности.
Над инертными, врожденными, общими для всех особей вида формами поведения, реализуемыми безусловными рефлексами, возникают и развиваются гибкие, приобретаемые путем жизненного опыта индивидуальные формы поведения, которые реализуются в простейших проявлениях в виде условных рефлексов. Такая высшая нервная деятельность развивается и совершенствуется, обеспечивая все более гибкое и тонкое приспособление поведения к меняющимся условиям окружающей среды.
Учение о высшей нервной деятельности, ознаменовавшее материалистический научный подход к пониманию природы разумного поведения и явлений психики, является великим достижением отечественной научной мысли. Его основы заложены трудами И.М. Сеченова и И.П. Павлова, оно творчески развивается в исследованиях советских ученых.
Аквариумистика — аквариум новичкам, аквариум любителям, аквариум профессионалам
Нервная система рыб гораздо примитивней нервной системы высших позвоночных и состоит из центральной и связанной с ней периферической и вегетативной (симпатической) нервной системой.
ЦНС рыбы включает в себя головной и спинной мозг.
Периферическая нервная система — это нервы, отходящие от головного и спинного мозга к органам.
Вегетативная нервная система — это ганглии и нервы, иннервирующие мышцы внутренних органов и кровеносных сосудов сердца.
Центральная нервная система тянется вдоль всего туловища: часть ее, находящаяся над позвоночником и защищенная верхними дугами позвонков, формирует спинной мозг, а широкая передняя часть, окруженная хрящевым или костным черепом — головной мозг.
Мозг рыбы условно делят на передний, промежуточный, средний, продолговатый и мозжечок. Серое вещество переднего мозга в виде полосатых тел находится в основном в основании и обонятельных долях.
В переднем мозгу происходит обработка информации, поступающей от органов обоняния. А также передний мозг регулирует движение и поведение рыбы. Например, передний мозг стимулирует и непосредственно участвует в регуляции таких важных для рыбы процессов, как икрометание, охрана икры, образование стаи, агрессия.
Промежуточный мозг отвечает за зрение рыбы: от него отходят зрительные нервы. К нижней стороне промежуточного мозга прилегает гипофиз, или питуитарная железа; в верхней части промежуточного мозга находится эпифиз, или пинеальная железа. Гипофиз и эпифиз являются железами внутренней секреции.
Кроме этого, промежуточный мозг участвует в координации движения, и работе других органов чувств.
Средний мозг имеет вид двух полушарий, а также самый большой объем. Доли (полушария) среднего мозга — первичные зрительные центры, обрабатывающие возбуждение, сигналы органов зрения, регуляции окраски, вкуса и равновесия; здесь же происходит и связь с мозжечком, продолговатым и спинным мозгом.
Мозжечок часто имеет форму маленького бугорка примыкающего сверху к продолговатому мозгу. Очень большой мозжечок у сомов, а у мормируса он самый большой среди всех позвоночных.
Мозжечок отвечает за координацию движений, поддержание равновесия, мышечную деятельность. Он связан с рецепторами боковой линии, синхронизирует деятельность других отделов мозга.
Продолговатый мозг состоит из белого вещества и плавно переходит в спинной мозг. Продолговатый мозг регулирует деятельность спинного мозга и вегетативной нервной системы. Он очень важен для дыхательной, скелетно-мышечной, кровеносной и других систем рыбы. Если уничтожить эту часть мозга, например, перерезав рыбу в области за головой, то она быстро умирает. Кроме этого, продолговатый мозг отвечает за связь со спинным.
От мозга отходят 10 пар черепно-мозговых нервов.
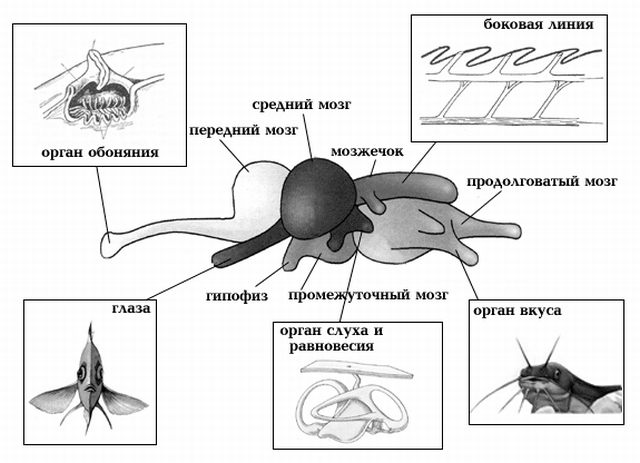
Как и большинство других органов и систем, нервная система развита по-разному у различных видов рыб. Это относится и к ЦНС (различная степень развития долей головного мозга) и к периферийной нервной системе.
Хрящевые рыбы (акулы и скаты) имеют более развитый передний мозг и обонятельные доли. Малоподвижные и донные рыбы имеют небольшой мозжечок и хорошо развитый передний и продолговатый отделы мозга, поскольку обоняние играет в их жизни значительную роль. У быстроплавающих рыб сильно развит средний мозг (зрительные доли) и мозжечок (координация движения). Слабые зрительные доли мозга у глубоководных рыб.
Спинной мозг — продолжение продолговатого мозга.
Особенностью спинного мозга рыбы является его способность к быстрой регенерации и восстановлению деятельности при повреждении. Серое вещество в спинном мозге рыбы находится внутри, а белое — снаружи.
Спинной мозг — это проводник и улавливатель рефлекторных сигналов. От спинного мозга отходят спинномозговые нервы, иннервирующие поверхность тела, туловищные мышцы, а через ганглии и внутренние органы. В спинном мозгу костистых рыб находится урогипофиз, клетки которого вырабатывают гормон, участвующий в водном обмене.
Вегетативная нервная система рыб — это ганглии, находящиеся вдоль позвоночника. Клетки ганглиев связаны со спинномозговыми нервами и внутренними органами.
Соединительные ветви ганглиев объединяют вегетативную нервную систему с центральной. Эти две системы независимы и взаимозаменяемы.
Одно из всем известных проявлений работы нервной системы рыбы — рефлекс. Например, если рыб кормить все время в одном и том же месте в пруду или в аквариуме, то они будут скапливаться именно в этом месте. Кроме того, условные рефлексы у рыб могут выработаться на свет, форму, запах, звук, вкус, температуру воды.
Рыбы вполне поддаются дрессировке и выработке у них поведенческих реакций.
Читайте также: