Аэробное дыхание. Особенности аэробного дыхания. Цикл Кребса.
Добавил пользователь Евгений Кузнецов Обновлено: 21.01.2026
Роль метоболитов в обмене веществ в клетке.— это окислительный процесс, в ходе которого расходуется кислород. При дыхании субстрат без остатка расщепляется до бедных энергией неорганических веществ с высоким выходом энергии. Важнейшими субстратами для дыхания служат углеводы. Кроме того, при дыхании могут расходоваться жиры и белки.
Аэробное дыхание включает два основных этапа:
1. бескислородный, в процессе которого происходит постепенное расщепление субстрата с высвобождением атомов водорода и связыванием с коферментами (переносчиками типа НАД и ФАД);
2. кислородный, в ходе которого происходит дальнейшее отщепление атомов водорода от производных дыхательного субстрата и постепенное окисление атомов водорода в результате переноса их электронов на кислород.
На первом этапе вначале высокомолекулярные органические вещества (полисахариды, липиды, белки, нуклеиновые кислоты и др.) под действием ферментов расщепляются на более простые соединения (глюкозу, высшие карбоновые кислоты, глицерол, аминокислоты, нуклеотиды и т. п ). Этот процесс происходит в цитоплазме клеток и сопровождается выделением небольшого количества энергии, которая рассеивается в виде тепла. Далее происходит ферментативное расщепление простых органических соединений.
Примером такого процесса является гликолиз — многоступенчатое бескислородное расщепление глюкозы. В реакциях гликолиза шестиуглеродная молекула глюкозы (С6) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С3). При этом образуются две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД+ (никотинамидадениндинуклеотид), который переходит в свою восстановленную форму НАД ∙ Н + Н+. НАД — кофермент, близкий по своей структуре к НАДФ. Оба они представляют собой производные никотиновой кислоты — одного из витаминов группы В. Молекулы обоих коферментов электроположительны (у них отсутствует один электрон) и могут играть роль переносчика как электронов, так и атомов водорода. Когда акцептируется пара атомов водорода, один из атомов диссоциирует на протон и электрон:
а второй присоединяется к НАД или НАДФ целиком:
НАД + + Н + [Н + + е — ] → НАД ∙ Н + Н + .
Свободный протон позднее используется для обратного окисления кофермента.
Суммарно реакция гликолиза имеет вид
Продукт гликолиза — пировиноградная кислота (С3Н4O3) - заключает в себе значительную часть энергии, и дальнейшее ее высвобождение осуществляется в митохондриях. Здесь происходит полное окисление пировиноградной кислоты до СO2 и Н2O. Этот процесс можно разделить на три основные стадии:
1) окислительное декарбоксилирование пировинофадной кислоты,
2) цикл трикарбоновых кислот (цикл Кребса);
3) заключительная стадия окисления — электронтранспортная цепь.
На первой стадии пировиноградная кислота взаимодействует с веществом, которое называют коферментом А (сокращенно его обозначают КоА), в результате чего образуется адетилкофермент А с высокоэнергетической связью. При этом от молекулы пировиноградной кислоты отщепляется молекула СO2 (первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н + .
Вторая стадия — цикл Кребса (названный так в честь открывшего его английского ученого Ганса Кребса).
В цикл Кребса вступает ацетил-КоА, образованный на предыдущей стадии. Ацетил-КоА взаимодействует со щавелево-уксусной кислотой (четырехутлеродное соединение), в результате образуется шестиуглеродная лимонная кислота. Для этой реакции требуется энергия; ее поставляет высокоэнергетическая связь ацетил-КоА. Далее превращение идет через образование ряда органических кислот, в результате чего ацетильные группы, поступающие в цикл при гидролизе ацетил-КоА, дегидрируются с высвобождением четырех пар атомов водорода и декарбоксилируются с образованием двух молекул СO2. При декарбоксилировании для окисления атомов углерода до СO2 используется кислород, отщепляемый от молекул воды. В конце цикла щавелево-уксусная кислота регенерируется в прежнем виде. Теперь она способна вступить в реакцию с новой молекулой ацетил-КоА, и цикл повторяется. В процессе цикла используются три молекулы воды, выделяются две молекулы СO2 и четыре пары атомов водорода, которые восстанавливают соответствующие коферменты (ФАД — флавина-дениндинуклеотид и НАД).
Суммарно реакция цикла может быть выражена следующим уравнением:
ацетил-КоА + ЗН2O + ЗНАД + + ФАД + АДФ + H3PO4 →
КоА + 2СO2 + ЗНАД ∙ Н + Н + + ФАД ∙ Н2 + АТФ.
Таким образом, в результате распада одной молекулы пировиноградной кислоты в аэробной фазе (декарбоксилирование ПВК и цикла Кребса) выделяется ЗСO2, 4НАД ∙ Н + Н+, ФАД ∙ Н2.
Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
С6Н12O6 + 6Н2O + 10НАД + 2ФАД → 6СO2 + 4АТФ + 10НАД ∙ Н + Н+ + 2ФАД ∙ Н2.
Третья стадия — электронтранспортная цепь.
Пары водородных атомов, отщепляемые от промежуточных продуктов в реакциях дегидрирования при гликолизе и в цикле Кребса, в конце концов окисляются молекулярным кислородом до Н2O с одновременным фосфорилированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ Н2 и ФАД ∙ Н2, передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2Н+ + 2е—. Именно в таком виде они и передаются по цепи переносчиков. Путь переноса водорода и электронов от одной молекулы переносчика к другой представляет собой окислительно-восстановительный процесс. При этом молекула, отдающая электрон или атом водорода, окисляется, а молекула, воспринимающая электрон или атом водорода, восстанавливается. Движущей силой транспорта атомов водорода в дыхательной цели является разность потенциалов.
С помощью переносчиков ионы водорода Н+ переносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство.
При переносе пары электронов от НАД на кислород они пересекают мембрану три раза, и этот процесс сопровождается выделением на внешнюю сторону мембраны шести протонов. На заключительном этапе электроны переносятся на внутреннюю сторону мембраны и акцептируются кислородом.
В результате такого переноса ионов Н+ на внешнюю сторону мембраны митохондрий в перимитохондриальном пространстве создается повышенная концентрация их, т. е. возникает электрохимический градиент протонов (ΔμН+).
Протонный градиент представляет собой как бы резервуар свободной энергии. Эта энергия используется при обратном потоке протонов через мембрану для синтеза АТФ. В ряде случаев может наблюдаться непосредственное использование энергии протонного градиента (ΔμН+). Она может обеспечивать осмотическую работу и транспорт веществ через мембрану против градиента их концентрации, использоваться на механическую работу и др. Таким образом, клетка располагает двумя формами энергии — АТФ и ΔμH+. Первая форма — химическая. АТФ растворяется в воде и легко используется в водной фазе. Вторая (ΔμH+) — электрохимическая — неразрывно связана с мембранами. Эти две формы энергии могут переходить друг в друга. При образовании АТФ используется энергия ΔμH+, при распаде АТФ энергия может аккумулироваться в виде ΔμH+.
Когда протонный градиент достигает определенной величины, ионы водорода из Н+-резервуара движутся по специальным каналам в мембране, и их запас энергии используется для синтеза АТФ. В матриксе они соединяются с заряженными частичками О2-, и образуется вода: 2Н+ + О2- → Н2O.
Процесс образования АТФ в результате переноса ионов Н+ через мембрану митохондрии получил название окислительного фосфорилирования. Он осуществляется при участии фермента АТФ-синтетазы. Молекулы АТФ-синтетазы располагаются в виде сферических гранул на внутренней стороне внутренней мембраны митохондрий.
В результате расщепления двух молекул пировиноградной кислоты и переноса ионов водорода через мембрану по специальным каналам синтезируется в целом 36 молекул АТФ (2 молекулы в цикле Кребса и 34 молекулы в результате переноса ионов Н+ через мембрану).
Следует обратить внимание на то, что ферментные системы ориентированы в митохондриях противоположно тому, как это имеет место в хлоропластах: в хлоропластах Н+-резервуар находится с внутренней стороны внутренней мембраны, а в митохондриях — с ее наружной стороны; при фотосинтезе электроны движутся в основном от воды к переносчикам атомов водорода, при дыхании же переносчики водорода, передающие электроны в электронтранспортную цепь, находятся с внутренней стороны мембраны, а электроны в конечном счете включаются в образующиеся молекулы воды.
Кислородный этап, таким образом, дает энергии в 18 раз больше, чем ее запасается в результате гликолиза. Суммарное уравнение аэробного дыхания можно выразить следующим образом:
Совершенно очевидно, что аэробное дыхание прекратится в отсутствие кислорода, поскольку именно кислород служит конечным акцептором водорода. Если клетки не получают достаточного количества кислорода, все переносчики водорода вскоре полностью насытятся и не смогут передавать его дальше. В результате основной источник энергии для образования АТФ окажется блокированным.
22. Аэробное дыхание. Химизм и использование энергии микроорганизмами
Аэробное дыхание - основной процесс катаболизма(энергетического метаболизма) многих прокариот. При аэробном дыхании донором водорода или других электронов являются обычно органические (реже неорганические) вещества, а конечным акцептором электронов- молекулярный кислород.
Аэробное дыхание характерно тем, что основное количество энергии при этом процессе образуется в электротранспортной цепи, то ест в результате мембранного фосфорилирования.
Зависимость микробов от аэробного дыхания
Аэробное дыхание присуще широкому кругу микроорганизмов. Однако различают микроорганизмы - строгие аэробыи факультативные анаэробы. Последние, способны расти как в присутствии, так и в отсутствии кислорода. Факультативные анаэробы способны синтезировать АТФ (аденозинтрифосфорную кислоту) при брожении, но при наличии молекулярного кислорода способ получения АТФ меняется и начинается осуществляться процесс дыхания.
К факультативным анаэробам относят микробы, у которых анаэробное дыхание происходит при использовании в качестве акцепторов электронов нитратов.
Строгими анаэробами считаются микробы, осуществляющее анаэробное дыхание, при котором акцепторами электронов служат карбонаты и сульфаты.
Фазы аэробного дыхания
Процесс аэробного дыхания подразделяется на две фазы:
Серия реакций, благодаря которым органический субстрат окисляется до CO2 , а освобождающиеся атомы водорода перемещаются к акцепторам. Эта фаза состоит из цикла реакций гликолиза, приводящих к образованию пирувата (пировиноградной кислоты) и цикла реакций Кребса(цикла трикарбоновых кислот- ЦТК).
Окисление освобождающихся атомов водорода кислородом с образованием аденозинтрифосфорной кислоты(АТФ).
Обе фазы приводят к окислению субстрата до углекислого газа (CO2) и воды (H2O), и образованию биологически полезной энергии в виде различных соединений: аденозинтрифосфата (АТФ); цитозинтрифосфата (ЦТФ); уридинтрифосфата (УТФ); гуанозинтрифосфата (ГТФ); креатинфосфата; ацетилфосфата.
Цикл Кребса
Цикл Кребса также называется циклом трикарбоновых кислот, так как они образуются в нем в качестве промежуточных продуктов. Представляет собой ферментативный кольцевой конвейер, «работающий» в матриксе митохондрий.
Результатом цикла Кребса является синтез небольшого количества АТФ и образование НАД · H2, который далее направляется на следующий этап клеточного дыхания- дыхательную цепь (окислительное фосфорилирование), расположенную на внутренней мембране митохондрий.
Образовавшаяся в результате гликолизапировиноградная кислота (пируват) поступает в митохондрии, где она в конечном итоге полностью окисляется, превращаясь в углекислый газ и воду. Сначала это происходит в цикле Кребса, затем при окислительном фосфорилировании.
До цикла Кребса пируват декарбоксилируется и дегидрируется. В результате декарбоксилирования отщепляется молекула CO2, Они соединяются с НАД.
В результате из пировиноградной кислоты образуется уксусная, которая присоединяется к коферменту А. Получается ацетилкофермент А (ацетил-КоА) - CH3CO~S-КоА, содержащий высокоэнергетическую связь.
Превращение пирувата в ацетил-КоА обеспечивает большой ферментативный комплекс, состоящий из десятков полипептидов, связанным с переносчиками электронов.
Цикл Кребса начинается с гидролиза ацетил-КоА, при котором отщепляется ацетильная группа, содержащая два атома углерода. Далее ацетильная группа включается в цикл трикарбоновых кислот.
Ацетильная группа присоединяется к щавелевоуксусной кислоте, имеющей четыре атома углерода. В результате образуется лимонная кислота, включающая шесть атомов углерода. Энергию для этой реакции поставляет макроэргическая связь ацетил-КоА.
Далее следует цепь реакций, в которых связанная в цикле Кребса ацетильная группа дегидрируются с высвобождением четырех пар атомов водорода и декарбоксилируются с образованием двух молекул CO2. При этом для окисления используется кислород, отщепляемый от двух молекул воды, а не молекулярный. Процесс называется окислительным декарбоксилированием. В конце цикла щавелевоуксусная кислота регенерируется.
Вернемся на этап лимонной кислоты. Ее окисление проходит за ряд ферментативных реакций, при которых образуются изолимонная, щавелевоянтарная и другие кислоты. В результате этих реакций, на разных стадиях цикла, восстанавливаются три молекулы НАД и одна ФАД, образуется ГТФ (гуанозинтрифосфат), содержащий макроэргическую фосфатную связь, энергия которой впоследствии используется для фосфорилирования АДФ. В результате образуется молекула АТФ.
Лимонная кислота теряет два атома углерода с образованием двух молекул CO2.
В результате ферментативных реакций лимонная кислота превращается в щавелевоуксусную, которая снова может соединиться с ацетил-КоА. Цикл повторяется.
В составе лимонной кислоты присоединившийся остаток ацетил-КоА сгорает с образованием углекислого газа, атомов водорода и электронов. Водород и электроны переносятся на НАД и ФАД, которые являются акцепторами для него.
Окисление одной молекулы ацетил-КоА дает одну молекулу АТФ, четыре атома водорода и две молекулы углекислого газа. То есть углекислый газ, выделяемый при аэробном дыхании, образуется на этапе цикла Кребса. При этом молекулярный кислород (O2) здесь не используется, он необходим лишь на этапе окислительного фосфорилирования.
Атомы водорода присоединяются к НАД или ФАД, в таком виде далее попадают в дыхательную цепь.
Одна молекула глюкозы дает две молекулы пирувата и, следовательно, два ацетил-КоА. Таким образом на одну молекулу глюкозы приходится два оборота цикла трикарбоновых кислот. В общей сложности образуются две молекулы АТФ, четыре CO2, восемь атомов H.
Следует отметить, что не только глюкоза и образующийся из нее пируват поступают в цикл Кребса. В результате расщепления ферментом липазой жиров образуются жирные кислоты, окисление которых также приводит к образованию ацетил-КоА, восстановлению НАД, а также ФАД (флавинадениндинуклеотида).
Если клетка испытывает дефицит углеводов и жиров, то окислению могут подвергаться аминокислоты. При этом образуются ацетил-КоА и органические кислоты, которые далее участвуют в цикле Кребса.
Таким образом неважно, каким был первичный источник энергии. В любом случае образуется ацетил-КоА, представляющий собой универсальное для клетки соединение.
Тема 15 «Аэробное дыхание»
4. Синтез молекул АТФ в дыхательной цепи бактерий и дрожжей.
1. Аэробное дыхание, цикл Кребса.
Аэробное дыхание - это основной процесс энергетического метаболизма многих прокариот, при котором донором водорода или электронов являются органические (реже неорганические) вещества, а конечным акцептором - молекулярный кислород. Основное количество энергии при аэробном дыхании образуется в электронтранспортной цепи, т. е. в результате мембранного фосфорилирования.
Рассмотрим схему аэробного дыхания (рис. 1).
Рис. 1. Схема аэробного дыхания
Пировиноградная кислота, образующаяся в одном из трех путей катаболизма глюкозы, окисляется с участием коэнзима А до ацетил-КоА. В данном процессе работают ферменты пируватдегидрогеназы:
Ацетил-КоА является исходным субстратом цикла Кребса (ЦТК).
В цикл Кребса включается одна молекула ацетил-КоА, которая в реакции с оксалоацетатом, катализируемой цитратсинтетазой, приводит к образованию лимонной кислоты и свободного коэнзима А. Лимонная кислота с помощью фермента аконитазы превращается в цис-акотиновую и изолимонную кислоты. Изолимонная кислота через щавелевоянтарную кислоту превращается в α-кетоглутаровую кислоту, которая подвергается дальнейшему декарбоксилированию.
В конечном итоге, окисление ацетил-КоА в ЦТК приводит к образованию (рис. 2):
- двух молекул СО2;
- одной молекулы АТФ;
- восьми атомов водорода, из которых шесть атомов связаны в молекулах пиридиннуклеотидов и два атома - в молекулах флавопротеинов.
Таким образом, ЦТК можно рассматривать как выработанный клеткой механизм, имеющий двоякое назначение:
1) Основная функция его заключается в том, что это - совершенный клеточный «котел», в котором осуществляется полное окисление вовлекаемого в него органического субстрата и отщепление водорода.
2) Другая функция цикла - обеспечивает биосинтетические процессы клетки различными предшественниками, такими как оксалоацетат, сукцинат, α-кетоглутарат и др. Отсутствие этих кислот привело бы к нехватке оксалоацетата, который служит акцептором для ацетил-СоА и, тем самым, к
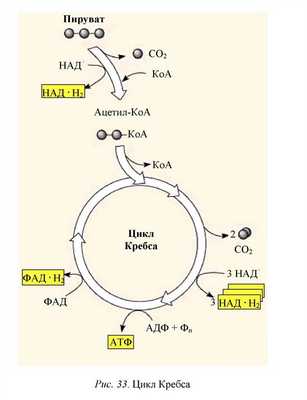
Рис. 2 Цикл Кребса
нарушению цикла. Обычно ЦТК является дальнейшей «надстройкой» над анаэробными энергетическими механизмами клетки.
У некоторых бактерий ЦТК «разорван». Наиболее часто отсутствует этап превращения α-кетоглутаровой кислоты в янтарную. В таком виде ЦТК не может функционировать в системе энергодающих реакций клетки. Основная функция «разорванного» ЦТК - биосинтетическая.
Образовавшиеся на разных этапах окисления органических веществ восстановительные эквиваленты НАД · Н2 и ФАД · Н2 поступают в дыхательную цепь, которая у бактерий находится в цитоплазматической мембране, а у эукариот - в мембране митохондрий. В дыхательной цепи НАД · Н2 и ФАД · Н2 вновь окисляются до НАД и ФАД, а отщепившийся от них водород передается не менее чем через пять переносчиков на заключительный участок цепи, где соединяется с молекулярным кислородом, образуя воду (рис. 1).
Аэробная фаза дыхания. Цикл Кребса, энергетический выход
В присутствии достаточного количества кислорода (аэробная фаза дыхания ) пируват полностью окисляется до СО2 и Н2О в дыхательном цикле, получившем название цикла Кребса или цикла ди- и трикарбоновых кислот. Английский биохимик Г. А. Кребс в 1937 г. предложил схему последовательности окисления ди- и трикарбоновых кислот до СО2. Этот цикл и был назван его именем. Цикл Кребса был открыт на животных объектах. Существование его у растений впервые доказал английский исследователь А. Чибнелл (1939).
Все участки этого процесса локализованы в матриксе или во внутренней мембране митохондрий. Весь процесс можно разделить на три стадии:
Первая стадия − окислительное декарбоксилирование пирувата. Непосредственно в цикле окисляется не сам пируват, а его производное - ацетил-СоА, образующегося в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование пирувата осуществляется при участии пируватдегидрогеназного мульти-ферментного комплекса, включающего три фермента и пять коферментов − тиаминпирофосфат (ТПФ) (фосфорилированное производное витамина В1), липоевая кислота, коэнзим A, FAD и NAD + .
Общая формула данного процесса:
В результате этого процесса образуется активный ацетат − ацетил-КоА, восстановленный НАД и выделяется СО2 (первая молекула).
Вторая стадия − цикл Кребса. Дальнейшее окисление ацетил-СоА осуществляется в ходе циклического процесса, в котором происходит постепенное преобразование ряда органических кислот. Ацетил-КоА конденсируется с енольной формой щавелевоуксусной кислоты (ЩУК) с образованием лимонной кислоты.
В этой реакции под действием фермента цитратсинтазы образуется лимонная кислота. Следующий этап цикла включает две реакции и катализируется ферментом аконитазой, или аконитатгидратазой. В первой реакции в результате дегидратации лимонной кислоты образуется цис-аконитовая. Во второй реакции аконитат гидратируется и синтезируется изолимонная кислота. Изолимонная кислота под действием НАД- или НАДФ-зависимой изоцитратдегидрогеназы окисляется в нестойкое соединение — щавелевоянтарную кислоту, которая тут же декарбоксилируется с образованием α -кетоглутаровой кислоты (α -оксоглутаровой кислоты).
α-Кетоглутарат подвергается реакции окислительного декарбоксилирования. При этом выделяется СО2, образуются NADH и сукцинил-СоА − высокоэнергетический тиоэфир. При участии сукцинил-СоА-синтетазы из сукцинил-СоА, ADP и Н3РО4 образуются янтарная кислота (сукцинат), АТР, регенерирует молекула СоА. АТР образуется в результате субстратного фосфорилирования.
Третий этап. Янтарная кислота окисляется до фумаровой. Реакция катализируется сукцинатдегидрогеназой, коферментом которой является FAD. Фумаровая кислота под действием фумаразы или фумаратгидратазы, присоединяя Н2О, превращается в яблочную кислоту (малат).
Четвертый этап. Яблочная кислота с помощью NAD-зависимой малатдегидрогеназы окисляется в щавелевоуксусную. ЩУК, которая самопроизвольно переходит в енольную форму, реагирует с очередной молекулой ацетил-СоА и цикл повторяется снова.
Большинство реакций цикла обратимы, однако ход цикла в целом практически необратим.
На протяжении одного оборота цикла при окислении пирувата происходит выделение трех молекул СО2, включение трех молекул Н2О и удаление пяти пар атомов водорода. Роль Н2О в цикле Кребса подтверждает правильность уравнения Палладина, который постулировал, что дыхание идет с участием Н2О, кислород которой включается в окисляемый субстрат.
Большинство ферментов цикла Кребса локализовано в матриксе митохондрий, аконитаза и сукцинатдегидрогеназа - во внутренней мембране митохондрии.
Энергетический выход цикла Кребса, его связь с азотным обменом
Цикл Кребса играет чрезвычайно важную роль в обмене веществ растительного организма. Он служит конечным этапом окисления не только углеводов, но также белков, жиров и других соединений. В ходе реакций цикла освобождается основное количество энергии, содержащейся в окисляемом субстрате, причем большая часть этой энергии утилизируется при образовании высокоэнергетических конечных фосфатных связей АТР.
В ходе окисления пирувата имеют место 5 дегидрирований, при этом получаются 3НАДН, НАДФH и ФАДH2. Окисление каждой молекулы НАДH (НАДФH) дает по 3 молекулы АТФ, а окисление ФАДH2 − 2ATФ. Таким образом при полном окислении пирувата образуются 14 молекул АТР. Кроме того, 1 молекула АТР синтезируется в цикле Кребса в ходе субстратного фосфорилирования. Следовательно, при окислении одной молекулы пирувата может образоваться 15 молекул АТР. А поскольку в процессе гликолиза из молекулы глюкозы возникают две молекулы пирувата, их окисление даст 30 молекул АТР.
Итого в ходе окислительного распада молекулы глюкозы в общей сложности образуются 38 молекул АТФ (8 АТФ связаны с гликолизом). Энергия третьей сложноэфирной фосфатной связи АТР равняется 41,87 кДж/моль (10 ккал/моль), то энергетический выход гликолитического пути аэробного дыхания составляет 1591 кДж/моль (380 ккал/моль), причем основное количество этой энергии − 1256 кДж/моль (300 ккал/моль) − поставляют реакции цикла Кребса. Эффективность использования энергии через гликолиз и ЦТК оказывается весьма высокой − 55,4%.
Не менее важную роль цикл Кребса играет в азотном обмене через участие ряда органических кислот в синтезе и распаде белковых веществ. Из кетокислот в ходе реакций переаминирования и восстановительного аминирования образуются аминокислоты. Из пировиноградной кислоты возникает аланин, из щавелевоуксусной и α -кетоглутаровой − соответственно аспарагиновая и глутаминовая кислоты. Для синтеза липидов, полиизопренов, углеводов и ряда других соединений используется ацетил-СоА. Особенно важно, что через реакции цикла Кребса устанавливается тесная связь между обменом трех важнейших групп соединений — белков, жиров и углеводов
Глиоксилатный цикл можно рассматривать как модификацию цикла Кребса. Этот цикл в 1957 г. был впервые описан у бактерий и плесневых грибов Г. Л. Корнбергом и Г. А. Кребсом. Затем оказалось, что он активно функционирует в прорастающих семенах масличных растений и в других растительных объектах, где запасные жиры превращаются в сахара (глюконеогенез).Глиоксилатный цикл локализован в специализированных микротелах - глиоксисомах. В клетках животных этот цикл отсутствует.
В глиоксилатном цикле из ЩУК и ацетил-СоА синтезируется лимонная кислота, образуются цис-аконитовая и изолимонная (изоцитрат), как и в цикле Кребса. Затем изолимонная кислота под действием изоцитратлиазы распадается на глиоксиловую и янтарную кислоты. Глиоксилат с участием малатсинтазы взаимодействует со второй молекулой ацетил-Co А, в результате чего синтезируется яблочная кислота, которая окисляется до ЩУК.
В отличие от цикла Кребса в глиоксилатном цикле в каждом обороте участвует не одна, а две молекулы ацетил-СоА и этот активированный ацетил используется не для окисления, а для синтеза янтарной кислоты. Янтарная кислота выходит из глиоксисом, превращается в ЩУК и участвует, в глюконеогенезе (обращенном гликолизе) и других процессах биосинтеза. Глиоксилатный цикл, позволяет утилизировать запасные жиры, при распаде которых образуются молекулы ацетил-СоА. Кроме того, на каждые две молекулы ацетил-СоА в глиоксилатном цикле восстанавливается одна молекула НАДH, энергия которой может быть использована на синтез АТФ в митохондриях или на другие процессы.
Аэробное дыхание
Аэробное дыхание Аэробное дыхание - основной процесс катаболизма (энергетического метаболизма). - основной процесс катаболизма (энергетического метаболизма) многих прокариот. При аэробном дыхании Аэробное дыхание - основной процесс катаболизма (энергетического метаболизма). донором водорода или других электронов являются обычно органические (реже неорганические) вещества, а конечным акцептором электронов - молекулярный кислород.
Аэробное дыхание Аэробное дыхание - основной процесс катаболизма (энергетического метаболизма). характерно тем, что основное количество энергии при этом процессе образуется в электротранспортной цепи, то есть в результате мембранного фосфорилирования.
Все организмы можно разделить на аэробные (способные жить при высоких концентрациях кислорода) и анаэробные (живут в бескислородных условиях, для них кислород является ядом). Для облигатных анаэробов кислород смертелен даже в очень низких концентрациях. Факультативные анаэробы способны жить как в присутствии, так и в отсутствие кислорода, переключаясь между двумя разными типами метаболизма.
Молекулярный кислород — очень активный окислитель, способный повреждать практически все биомолекулы — ДНК, РНК, белки, липиды мембран и т.д. Поэтому клетки аэробов Аэробы - организмы, в том числе микроорганизмы,в том числе бактерии, для жизнеде. и факультативных анаэробов Анаэробы - это микроорганизмы, в том числе бактерии, энергетические процесс. имеют специальные системы защиты от токсического действия кислорода (особые ферменты, а также вещества-антиоксиданты, “принимающие на себя удар” кислородного окисления). Клетки облигатных анаэробов Анаэробы - это микроорганизмы, в том числе бактерии, энергетические процесс. таких систем лишены.
Почему же, несмотря на токсическое действие кислорода, аэробный метаболизм выгоден и дает эволюционные преимущества?
В процессах анаэробного метаболизма полное окисление органики до СО2 и Н2О невозможно, так как для этого нет достаточно хорошего окислителя. Поэтому в результате анаэробных процессов катаболизма — брожений Брожение - одна из основных форм катаболизма, представляющая собой окислитель. — всегда получаются какие-то недоокисленные органические продукты (молочная кислота, масляная кислота, этиловый спирт и т.д.). Наличие же в окружающей среде кислорода позволяет разорвать все С-С-связи в органическом субстрате, окислить его до СО2 и Н2О и получить максимальное количество энергии.
В присутствии кислорода НАД∙Н не отдаёт атомы водорода на продукты гликолиза, как это происходит при брожении Брожение - одна из основных форм катаболизма, представляющая собой окислитель. . Вместо этого имеет место дальнейшее окисление пировиноградной кислоты до СО2 и Н2О (полное окисление органики). При этом выделяется гораздо больше энергии, чем в процессе анаэробного (бескислородного) окисления. Так происходит потому, что молекулярный кислород является хорошим окислителем, поэтому в реакциях кислородного окисления выделяется много энергии. Это известно вам из бытового опыта — реакцией кислородного окисления органики является и реакция горения. При этом выделяется энергия в виде тепла и света.
Зависимость микробов от аэробного дыхания
Аэробное дыхание Аэробное дыхание - основной процесс катаболизма (энергетического метаболизма). присуще широкому кругу микроорганизмов. Однако различают микроорганизмы - строгие аэробы и факультативные анаэробы Анаэробы - это микроорганизмы, в том числе бактерии, энергетические процесс. . Последние, способны расти как в присутствии, так и в отсутствии кислорода. Факультативные анаэробы способны синтезировать АТФ (аденозинтрифосфорную кислоту) при брожении Брожение - одна из основных форм катаболизма, представляющая собой окислитель. , но при наличии молекулярного кислорода способ получения АТФ меняется и начинается осуществляться процесс дыхания.
Строгими анаэробами считаются микробы, осуществляющее анаэробное дыхание Анаэробное дыхание - это такое дыхание микробов, при котором для окисления орган. , при котором акцепторами электронов служат карбонаты и сульфаты.
Фазы аэробного дыхания
Процесс аэробного дыхания Аэробное дыхание - основной процесс катаболизма (энергетического метаболизма). подразделяется на две фазы:
- Серия реакций, благодаря которым органический субстрат окисляется до CO2 , а освобождающиеся атомы водорода перемещаются к акцепторам. Эта фаза состоит из цикла реакций гликолиза, приводящих к образованию пирувата (пировиноградной кислоты) и цикла реакций Кребса (цикла трикарбоновых кислот - ЦТК).
- Окисление освобождающихся атомов водорода кислородом с образованием аденозинтрифосфорной кислоты(АТФ).
Обе фазы приводят к окислению субстрата до углекислого газа (CO2) и воды (H2O), и образованию биологически полезной энергии в виде различных соединений: аденозинтрифосфата (АТФ); цитозинтрифосфата (ЦТФ); уридинтрифосфата (УТФ); гуанозинтрифосфата (ГТФ); креатинфосфата; ацетилфосфата.
Читайте также:
- Половые хромосомы. Особенности Y-хромосом
- Разрывы миокарда при инфаркте миокарда ( оим, ОИМ ). Разрыв стенки желудочков сердца. Разрыв межжелудочковой перегородки. Разрыв сосочковой мышцы.
- Причины переношенной беременности. Последствия и осложнения перенашивания
- Эрозия зубов: причины, симптомы и лечение
- КТ изменения лимфоузлов