Дегенерация и регенерация периферического нерва. Скорость роста нерва
Добавил пользователь Евгений Кузнецов Обновлено: 22.01.2026
1. Гомазков О.А. Нейрогенез, как адаптивная функция мозга. - М.: И-т биомедицинской химии, 2014. - 85 с.
2. Григорян А.С., Кругляков П.В. Клеточная терапия при травме мозга // Клет. трансплантология и тканевая инженерия. - 2008. Т.4, № 1. - С. 35-42.
3. Коржевский Д.Э. Петрова Е.С., Кирик О.В., Безлин Г.В., Сухорурока Е.Г. Нейральные маркеры, используемые при изучении дифференцировки стволовых клеток // Клет. трансплантология и тканевая инженерия. - 2010. - Т.5, № 3. - С. 57-63.
4. Коржевский Д.Э., Кирик О.В., Григорьев И.П., Сухорукова Е.Г. Сырцова М.А. Маркирование дифференцирующихся нервных клеток при изучении развития и патологии головного мозга // Вопросы морфологии ХХI века. - 2015. - Вып. 4. - С. 34-36.
5. Обухов Д.К., Пущина Е.В. Радиальная глия - как источник новых нейронов в постнатальном развитии ЦНС // Межд. журн. экспер. обр. - 2011. - № 6. - С. 10-11.
6. Обухов Д. К., Пущина Е. В., Вараксин А. А. Структура пролиферативных зон в ЦНС взрослых позвоночных животных // Вопросы морфологии ХХI века. - 2015. - Вып. 4. - С. 43-51.
7. Полежаев Л.В., Александрова М.А., Витвицкий В.Н. Трансплантация ткани мозга в биологии и медицине. - М.: Наука, 1993. - 239 с.
8. Пущина Е.В., Вараксин А.А., Обухов Д.К. Газообразные посредники в головном мозге симы // Журн.эвол.физиол. и биох. - 2012. - Т. 48. - С. 85-96.
9. Пущина Е.В., ,Вараксин А.А., Обухов Д.К. Репаративный нейрогенез в мозге и изменения в зрительном нерве взрослой форели после механического повреждения глаза // Онтогенез. - 2016. - Т. 47, № 1. - С. 1-24.
10. Семченко В.В., Еринеев С.И., Степанов С.С. Сергиенко Г.Г. Трансплантация незрелой нервной ткани в экспериментальной и клинической неврологии. - Омск: Омский дом печати, 2000. - 340 с.
11. Семченко В.В.. и др., Регенеративная биология и медицина. Книга I. Генные технологии и клонирование / под. ред, В.П. Пузырева и др. - Омск. 2012. - 296 с.
12. Яковлев А.В., Ситдикова Г.Ф. Физиологическая роль сероводорода в нервной системе // Гены и клетки. - 2014. - Т.9, № 3. - С. 34-44.
13. Ярыгин К.Н., Ярыгин В.Н.. Нейрогенез в центральной нервной системе и перспективы регенеративной неврологии // Журнал неврологии и психиатрии им. С. С. Корсакова. - 2012. - Т. 112, № 1. - С. 4-13.
14. Ярыгин К.Н. и др., Регенеративная биология и медицина Книга II. Клеточные технологии в терапии болезней нервной системы / под ред. В.Н. Ярыгина и др. - Екатеринбург-Омск, 2015. - 360 с.
15. Cameron HA, McKay RD. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus // J Comp Neurol. - 2001. - Vol. 435. - P. 406-417.
16. Grandel H , Brand M. Comparative aspects of adult neural stem cell activity in vertebrates // Dev. Genes Evol. - 2013. - Vol. 223. - P. 131-147.
17. Pinto L., Götz M. Radial glial cell heterogeneity-The source of diverse progeny in the CNS // Progress in Neurobiology. - 2007. - Vol. 83. - P. 2-23.
18. Pushchina E.V., Obukhov D.K., Varaksin A.A., Shukla S. Neurochemical signaling and participation of H2S and NO in fishes adult neurogenesis // Nitric Oxide. - 2014. - Vol. 39 (Suppl). - P. 41-43.
19. Pushchina E.V., Obukhov D.K., Varaksin A.A. Structure, сhemoarchitectonics and postembryonic histogenesis of a central nervous system in a teleost fish // In book:Teleosts: Evolutionary Development, Diversity and Behavioral Ecology / Ed. Carone S. New York: - Nova Science Publishers Inc. - USA, 2014. - Ch. 5. - P. 97-152.
20. Pushchina E. V., Varaksin A. A., Obukhov D. K. Participation of neurochemical signaling in adult neurogenesis and differentiation // In book: Neurochemistry (edt. Th.Heinboocken). 2014. - Intech Corp. USA. Ch.8. - P. 225-255.
21. Pushchina E. V., Varaksin A. A., Obukhov D. K. Cystathionine β-Synthase in the CNS of Masu Salmon Oncorhynchus masou (Salmonidae) and Carp Cyprinus carpio (Cyprinidae) // Neurochemical Journal. - 2011. - Vol. 5. - P. 24-34.
22. Pushchina E.V., Obukhov D.K. Is the brain of cherri salmon a new model for investigation of postembryonic neurogenesis? // Engineering. Supplement. - 2012. - P. 76-79.
24. Zupanc G Towards brain repair: insights from teleost fish // Seminars in Cell & Developmental Biology. - 2009. - Vol. 20. - P. 683-690.
25. Zupanc G.K.H., Sîrbulescu R.F. Teleost Fish as a Model System to Study Successful Regeneration of the Central Nervous System // Current Topics in Microbiology and Immunology. - 2013. - Vol. 367. - P. 193-233.
Введение. Проблема физиологической и репаративной регенерации нервной системы всегда была в центре внимания нейробиологов и неврологов. В конце ХХ века большое количество исследований было посвящено исследованию трансплантации нервной ткани. (Полежаев и др., 1993; Семченко и др., 2000). Однако, несмотря на определенные достижения, полноценного приживления нервной ткани и восстановления функциональных связей при разного видах алло- и ксенотрансплантациях достигнуть не удалось.
Открытие нейрональных стволовых клеток (НСК), их обнаружение во взрослой нервной системе позвоночных животных и человека и развитие клеточных технологий позволило по новому взглянуть на эту проблему (Семченко и др. 2012; К.Н. Ярыгин, В.Н. Ярыгин, 2012; Ярыгин и др., 2015). В данной работе приводится краткий обзор собственных и имеющихся в литературе данных по регенерации нервной ткани в норме и в условиях эксперимента.
Нейрональные стволовые клетки (НСК) относятся к группе тканеспецифичных или региональных стволовых клеток. Они обладают характеристиками самоподдерживающейся популяции клеток, которые при дифференцировке способны давать нейроны, астроциты и олигодендроциты в развивающемся и взрослом мозге. Впервые они были обнаружены в ЦНС человека в 1995 году при анализе посмертных срезов мозга, окрашенных иммунногистохимически на BrdU (бромдезоксиуридин) (Gage et al., 1995). В настоящее время найден целый ряд нейрональных маркеров, которые позволяют более или менее надежно идентифицировать НСК и их потомки. Среди них следует отметить: ядерный антиген нервных клеток - NeuN; маркер нейробластов - даблкортин (DCX); нейрон-специфическую энолазу - NSE; молекулы адгезии нервных клеток - PSA-NCAM; цитоскелетные белки - нестин, β-тубулин III; транскрипционные факторы - Sox-1, Sox-2, Dlx2, Pax 6 и ряд других. Часть из этих маркеров специфична для клеток нервной ткани, другие направлены на выявление свойств, характерных для разных популяций стволовых клеток. (Гомазков, 2014; Коржевский и др., 2010, 2015).
Обнаружение НСК как в развивающемся, так и во взрослом мозге поставило вопрос об их происхождении. В ранний период эмбрионального развития НСК происходят из клеток нейроэпителия, которые путем симметричного и асимметричного деления дают начало нейронам, глиальным клеткам и клеткам-предшественникам, которые включаются в процессы нейрогенеза на более поздних этапах развития, включая постнатальный период. Характер деления клеток - один из механизмов выбора НСК путей развития. В случае симметричного митоза образуются две одинаковые дочерние клетки, которые либо сохраняют пролиферативный потенциал - т.е. остаются стволовыми, либо могут уйти на путь нейрогенеза или глиогенеза - опять же обе. При втором варианте симметричного деления популяция стволовых клеток может потерять способность к самообновлению и истощится. При ассиметричном делении одна клетка остается пролиферативной (стволовой), другая выходит в дифференцировку. При этом пул НСК сохраняется.
Популяция нейрональных клеток - предшественников достаточно гетерогенна. На основную роль предшественника в позднем пренатальном и в постнатальном периодах претендует т.н. «радиальная глия - RG». Она выполняет двоякую роль - ее отростки, пронизывающие всю толщу развивающейся стенки нервной трубки, служат направляющими для миграции молодых нейробластов, а также обладает потенциями НСК, давая начало новым популяциям нейронов и глии. Особо необходимо отметить, что именно из потомков радиальной глии формируются основная масса интернейронов коры головного мозга млекопитающих животных и человека. Иммунологически в клетках радиальной глии помимо маркеров, традиционных для глиальных клеток (ГКФБ, виментин, нестин), выявляются специфические маркеры радиальной глии: фермент ароматаза-В (Aro-B), BLBP (brain lipid binding protein) и GLAST - глутаматный транспортер. Фермент ароматаза-В (Aro-B) связан с синтезом ароматизированных стероидов и синтезируется в клетках радиальной глии мозга молодых и взрослых позвоночных животных. Для них также характерна экспрессия полисиаловой молекулы адгезии нервных клеток (PSA-NCAM), транскриптационного фактора Sox-2 и фактора RG2. Важно подчеркнуть, что в клетках радиальной глии обнаруживаются маркеры и нейрональной линии дифференцировки (ТН- тирозингидроксилаза, ГАМК и NADP - диафораза). (Обухов, Пущина, 2011). На роль нейрональных предшественников также претендуют клетки эпендимы, астроциты, NG2 клетки и танициты (Гомазков, 2014; Pinto, Götz. 2007).
Таким образом, во взрослом мозге позвоночных животных и человека сохраняются группы клеток-предшественников, которые обладают свойствами НСК и способны в течение длительного периода обновлять популяции нейронов и глии.
Нейрогенные ниши. Нейрональные стволовые клетки и их потомки находятся в тесном взаимодействии со многими элементами окружающей их структуры мозга, формируя вмести с ними своеобразную - нейрогенную нишу (stem niche). В ее состав входят НСК и их потомки, клетки эпендимы, астроциты, олигодендроциты, эндотелий капилляров мозга и компоненты межклеточного матрикса (Обухов и др., 2015). Клетки «ниши» способны экспрессировать целый ряд факторов, необходимых для сохранения популяции НСК и регуляции нейро- и глиогенеза. Среди факторов, влияющих на нейрогенез следует отметить группу транскрипционных факторов (Shh, Sox1, Sox2, Tbr1, Wnt, BMP, Notch1, Pax6 и др). Они действуют на разные стадии нейро- и глиогенеза, причем часто прямо противоположно. Например: транскрипционные факторы Notch1 и BMP подавляют нейрональную дифференцировку, направляя развитие клеток предшественников в глиальном направлении, а фактор Trb2 наоборот - стимулирует нейрогенез. Фактор Shh (sonic hedgehog) и транскрипционные факторы из семейства Sox регулируют процесс пролиферации клеток-предшественников. Эти факторы могут влиять и на другие этапы нейро-и глиогенеза, проходящие в пролиферативных зонах (миграцию нейробластов, образование определенных типов клеток, формирование отростков у нейронов, развитие синаптических связей). Разнообразные ростовые факторы (эпидермальный фактор роста - EGF, трансформирующий фактор роста - TGFa, основной фактор роста фибробластов - bFGF, инсулиноподобный ростовой фактор - IGF1, фактор роста эндотелия - VEGF; интерферон гамма - IFN-γ и др.) также влияют на пролиферацию и дифференцировку клеток-предшественников. Особое место среди сигнальных молекул занимают нейромедиаторы и нейромодуляторы. В настоящее время установлено, что нейроны вскоре после образования из клеток-предшественников и задолго до начала миграции и формирования межнейрональных связей начинают секретировать молекулы нейромедиаторов, которые оказывают существенное влияние на развитие клеток в течение эмбриогенеза, а также в ходе постэмбрионального нейрогенеза. В последнее время особое внимание уделяется роли газообразных посредников (NO, H2S, CO) в регуляции процессов нейрогенеза в пре- и постнатальном периодах развития ЦНС. Было показано, что они существенно влияют на процесс миграции нейробластов, рост аксонных и дендритных ветвлений, пролиферацию и апоптоз НСК и их потомков в нейрогенных нишах и за их пределами (Пущина и др., 2012; Яковлев, Ситдикова, 2014; Puschina et al., 2011, 2014).
Особо следует отметить, что сами НСК способны синтезировать и секретировать подобные вещества, действующие в данном случае по типу пара- или аутокринной регуляции.
Организация пролиферативных зон в мозге млекопитающих. Зоны взрослого нейрогенеза у млекопитающих, включая приматов, обнаружены в субвентрикулярной зоне (SVZ) латеральных мозговых желудочков конечного мозга и в субгранулярной зоне (SGZ) зубчатой фасции гиппокампа. Наличие подобных зон в других отделах ЦНС млекопитающих в настоящее время не доказано, а имеющиеся данные носят крайне противоречивый характер. (Ярыгин и др., 2014; Ярыгин, Ярыгин, 2012). Субвентрикулярная зона (SVZ) образована несколькими слоями клеток (от двух до пяти), в составе которых выделяют несколько типов клеток. Скорость увеличения числа новых клеток в зубчатой извилине гиппокампа (SGZ) взрослого мозга определяется как 9000 единиц в течение суток, что составляет примерно 6 % от ощего количества нейронов в зубчатой фасции гиппокампа крысы.или около 250 тысяч в месяц. (Cameron, McKay, 2001). Вновь образованные нервные клетки мигрируют на места своей локализации в данной структуре мозга, формируют систему отростков и синапсов и встраиваются в функциональные нейронные сети. Cледует отметить, что, хотя факт интеграции новых нейронов в существующие нейронные сети доказан, функциональные аспекты этого процесса во многом еще неясны.
В этом плане весьма интересным является обнаружение подобных пролиферативных зон в разных отделах головного мозга у представителей других групп позвоночных животных (рыб, амфибий, птиц). Взрослый нейрогенез у этих животных идет более интенсивно и дольше, чем у млекопитающих. (Обухов и др., 2015; Puschina et al., 2014; Grandel, Brand, 2013).
Взрослый нейрогенез и перспективы репаративной регенерации нервной ткани. Исследование последствий ишемии мозга показали, что она сопровождается усилением нейрогенеза в пролиферативных зонах и миграцией молодых клеток в зону повреждения. (Гомазков, 2014; Solway et al., 1998). Эти данные вызвали целую серию работ, направленных на изучение возможности использования НСК и их потомков для трансплантации в поврежденный мозг, а также поиска модельных объектов для экспериментальных работ (Григорьян, Кругляков, 2008; Семченко и др., 2012). Одной из удачных моделей явились рыбы разных видов. В серии работ с помощью иммуногистохимического маркирования PCNA (пролиферативного ядерного антигена), ядерного маркера нейрональной дифференцировки (HuCD), транскриптационного фактора Pax6 и серии других маркеров, в разных отделах мозга рыб был идентифицирован ряд пролиферативных зон (ПВЗ), свидетельствующих о наличии постоянного постнатального нейрогенеза в ЦНС рыб. (Обухов и др., 2015; Zupanc, 2009; Zupanc, Sîrbulescu, 2013; Puschina, Obukhov, 2012; Pushchina et al., 2014 a, b). Однако, в настоящее время неизвестно как этот процесс связан с нейрогенезом во взрослом мозге, и какие элементы матричных зон мозга рыб участвуют в репаративном нейрогенезе.
Были поставлены эксперименты на молоди нескольких видов рыб, которым наносили механическую травму в разные структуры мозга (зрительный нерв, крыша среднего мозга, полушария конечного мозга). При механическом повреждении разных отделов мозга молоди рыб (сетчатки, среднего мозга и мозжечка) было выявлено усиление пролиферативной активности как в традиционных пролиферативных зонах нейрогенеза (перивентрикулярные области), так зафиксировано появление новых нейрогенных участков. Процесс репарации после нанесения механической травмы глаза начинается с апоптоза поврежденных элементов. Апоптотический ответ наблюдается уже через полчаса после нанесения повреждающего воздействия и продолжается до 21 дня после нанесения травмы. Эти данные подтверждены результатами маркирования TUNEL-позитивных фрагментов ДНК в зоне повреждения (зрительного нерва), а также данными электронно-микроскопического анализа. Ультраструктурные изменения ядра свидетельствуют о различных стадиях процесса апоптоза в поврежденных клетках. Апоптоз, как механизм элиминации поврежденных в результате травмы клеток мозга рыб существенно отличается от такового у млекопитающих. У последних, основным способом элиминации поврежденных клеток в зоне травмы является некроз. Апоптоз же затрагивает незначительный объем клеток в прилегающих к травме областях. Наличие некроза в зоне травмы млекопитающих является одной из причин развития последующего вторичного воспаления в зоне повреждения, что в свою очередь вызывает дальнейшее нарастание некротического ответа в области травмы, в результате которого формируются большие полости, лишённые клеток. Эти полости, как правило, ограничены зоной реактивных астроцитов, создающих как механический, так и биохимический барьеры, затрудняющие рост нервных волокон и миграцию клеток в зону повреждения. В отличие от некроза при апоптозе отсутствуют признаки воспалительной реакции, а сами клетки впоследствии уничтожаются с помощью макрофагов/микроглии. Прижизненный мониторинг клеток в зоне повреждения с помощью мультифотонной конфокальной микроскопии показал, что уже через час после повреждающего воздействия наблюдается физиологический ответ со стороны макрофагов и микроглии, которые мигрируют в область нанесения механической травмы и активно участвуют в элиминации поврежденных клеток с помощью фагоцитоза. Дифференцировка клеток в нейрональном направлении, обнаруженная при помощи маркирования клеток антителами против белка HuC/D, происходила в пролиферативных зонах теленцефалона, зрительного тектума, мозжечка и продолговатого мозга форели уже через 2 дня после травмы (Пущина и др., 2016; Puschina et al., 2014).
Таким образом, показано, что после механической травмы в мозге экспериментальных животных источником новых нейронов являются появляющиеся в пролифератиных областях мозга новые зоны индуцированного нейрогенеза: нейрогенные ниши и участки вторичного нейрогенеза. (Пущина и др., 2016). Полученные данные послужат основой для дальнейших исследований особенностей постнатального нейрогенеза в ЦНС животных и человека в норме и при патологии.
Работа выполнена при финансовой поддержке гранта Президента РФ (МД 4318.2015.04) и Программы фундаментальных исследований ДВО РАН «Дальний Восток» (проект № 15-I-6-116).
7.2. Механизмы, прогноз и признаки восстановления функции нервных проводников
7.2.1. Механизмы восстановления функций при повреждении периферических нервов и сплетений
Поврежденное нервное волокно не способно срастаться. Однако одновременно с процессом денервации начинаются восстановительные процессы, которые могут идти по трем направлениям.
(1) Регенерация нерва: проксимальная культя формирует аксональные выросты (наплывы аксоплазмы, или «колбы роста»), которые начинают продвигаться дистально и врастать в эндоневральные трубки (разумеется, лишь в тех случаях, когда последние сохранили свою целостность). Миелиновая оболочка вновь образующегося волокна формируется из тяжей леммоцитов. Скорость регенерации аксонов составляет приблизительно 1.5-2 мм в день [Buchtal F., Kuchl V., 1979]. Отдельные нервные проводники имеют различную способность к регенерации: среди периферических нервов особенно хорошо восстанавливается функция лучевого и мышечно-кожного нервов, а наихудшими регенеративными способностями обладают локтевой и малоберцовый нервы [Карчикян С.И., 1962; Weber R., 1996J. Для достижения хорошего восстановления растущие аксоны должны подключиться к дистальной культе нерва до того, как в ней наступят выраженные пери- и эндоневральные рубцовые сращения [Lefferet R., 1985]. В случаях формирования по ходу прорастающего волокна соединительнотканного рубца часть аксонов не распространяется в дистальном направлении, а беспорядочно отклоняется в стороны, формируя травматическую неврому.
При полном анатомическом повреждении нервного ствола через 2-3 недели после травмы на центральном конце образуется ампутационная неврома.
Регенерация нервного ствола может происходить гетерогенно: часть двигательных волокон врастает в оболочки чувствительных, а одноименные - в пучки, иннервирующие разноименные участки конечности [Гайдар Б.В.,1997].
(2) В случаях, когда в нервном стволе поражаются не все, а лишь часть нервных волокон, восстановление функции мышц возможно за счет разветвления сохранившихся аксонов и "захвата" ими тех мышечных волокон, которые иннервировались погибшими аксонами; при этом происходит укрупнение двигательных единиц мышцы. За счет этого механизма мышца может сохранять свою работоспособность в случаях утраты до 50% иннервировавших ее аксонов (а для мышц, не развивающих значительных усилий - даже до 90%), однако для полного завершения процесса компенсаторной иннервационной перестройки требуется около года [Weber R.,1996].
(3) В ряде случаев (обычно при травме нервного ствола типа ушиба) восстановление или улучшение функций связано с обратимостью некоторых патоморфологических процессов: с исчезновением реактивных воспалительных явлений, с рассасыванием мелких кровоизлияний и т.д. При нетяжелых травмах проводимость нерва, даже после полной ее потери, восстанавливается в течение первых дней или недель.
7.2.2. Факторы, определяющие прогноз восстановления
К основным факторам, определяющим при периферических невропатиях и плексопатиях скорость и степень спонтанного восстановления нарушенных функций (и, следовательно, объем и направленность лечебных вмешательств), относятся следующие:
- степень повреждения нервного проводника;
- уровень поражения;
- характер повреждающего агента.
7.2.2.1. Степень повреждения нервных проводников (при локальной травме)
Реабилитологи чаще всего определяют степень повреждения нерва по 3 категориям согласно классификации H.Seddon [1943]. Иногда используют также классификацию S.Sunderland [1990], выделяющего 5 степеней повреждения нервов; эта классификация основана на классификации H.Seddon, детализируя ее. Согласно классификации H.Seddon, все локальные повреждения нервных стволов делят, в зависимости от сохранности аксона и соединительнотканных структур, на три группы: (1) нейрапраксия; (2) аксонотмезис; (3) нейротмезис. (1) Нейрапраксия (neurapraxia, англ.) - это повреждение нерва, не приводящее к гибели аксона. Часто наблюдается при компрессии нерва (например, "ночной субботний паралич" вследствие компрессии лучевого нерва), при легкой травме нерва. Клинически характеризуется снижением вибрационной, проприоцептивной, иногда тактильной чувствительности. Болевая чувствительность страдает реже. Часто наблюдаются двигательные нарушения и парестезии. Блок проведения нервного импульса, наблюдающийся вследствие локального повреждения миелиновой оболочки, носит преходящий характер и регрессирует по мере восстановления миелина. Восстановление двигательных и чувствительных функций может продолжаться до 6 месяцев.
(2)Аксонотмезис (axonotmesis, англ.) - повреждение нерва, приводящее к гибели аксона при сохранности эпиневрия, периневрия, эндоневрия и шванновских клеток. Нередко наблюдается при закрытых переломах либо вывихах костей конечностей, а также при сдавлениях нервных стволов. Нарушаются двигательные, чувствительные и судомоторные функции нерва. Восстановление функций происходит за счет регенерации аксона. Скорость и степень восстановления зависит от уровня поражения, возраста (у молодых регенерация происходит быстрее) и общего состояния больного. В случаях, когда прорастание аксона происходит медленно, может произойти рубцевание эндоневральной трубки, в которую прорастает аксон, и восстановление не наступает. По этой же причине неблагоприятный прогноз имеется в случаях, когда дефект нервного ствола имеет значительную длину. При благоприятных условиях происходит постепенная невротизация дистального отдела поврежденного нерва, которая продолжается в течение многих месяцев, иногда год и более. Наблюдается восстановление утраченных функций, но не всегда полное.
О) Нейротмезис (neurotmesis, англ.) - разрыв нерва с пересечением аксона и соединительнотканных оболочек нерва. Из-за того, что повреждаются эндоневральные трубки, невозможным становится прорастание в них аксонов, регенерация аксонов приводит к образованию травматической невромы. Прогноз восстановления неблагоприятный. Данная классификация основана на микроскопических изменениях в нервном стволе. Макроскопически различить степень повреждения практически невозможно. Диагностика основана на динамическом клиническом и электрофизиологическом наблюдении. В связи с этим при закрытых травмах нервных стволов отечественные авторы нередко применяют иную классификацию, основанную на выделении следующих 4 форм поражения нервного ствола [Макаров А.Ю., Амелина О.А., 1998]: сотрясение, ушиб, сдавление, тракция. Сотрясение не сопровождается морфологическими изменениями в нерве, нарушения функции нерва кратковременны (не более 1-2 недель) и полностью обратимы. Ушиб нерва характеризуется возникновением мелких кровоизлияний, участков размозжения нервных волокон и пучков, что приводит к полному либо частичному нарушению проводимости, длительным и стойким выпадением функций. При сдавлении нерва степень нарушения проводимости зависит в первую очередь от длительности сдаачения: при своевременном удалении сдвливающих нерв субстратов (гематома, инородное тело, отломок кости и т.д.) может наблюдаться быстрое и полное восстановление проводимости, тогда как при длительном сдавлении в нервном стволе развиваются дегенеративные изменения. Отсутствие восстановления функции в течение 2-3 месяцев является критерием полного анатомического перерыва нерва. Тракция (например, тракдия ветвей плечевого сплетения при вправлении вывиха плеча) обычно сопровождается частичным нарушением функции, однако восстановление проводимости по нерву происходит достаточно длительно (в течение нескольких месяцев).
7.2.2.2. Уровень поражения
7.2.2.3. Характер повреждающего агента
Периферические невропатии и плексопатии могут иметь самую различную этиологию (таблица 7.2). В мирное время наиболее частой формой поражений периферических нервов являются туннельные невропатии, составляя около 30-40% от всех заболеваний периферической нервной системы. Туннельная невропатия - это локальное поражение нервного ствола, обусловленное его компрессией и ишемией в анатомических каналах (туннелях) или вследствие внешнего механического воздействия [Лейкин И.Б.,1998]. Предрасполагающие к развитию туннельных невропатий факторы включают генетически обусловленную узость естественных вместилищ нерва, приобретенную узость этих вместилищ вследствие отеков и гиперплазии соединительной ткани при различных заболеваниях (например, сахарном диабете, гипотиреозе, коллагенозах), длительное перенапряжение мышечно-связочного аппарата у лиц определенных профессий, последствия трав, мышечно-тонические и нейро-дистрофические нарушения при рефлекторных синдромах остеохондроза позвоночника, ятрогенные травмирующие воздействия (неправильное наложение гипсовой повязки, кровоостанавливающего жгута). Нарушение функции нерва происходит вследствие как демиелинизации, так и поражения аксона (ухудшение нейротрофического контроля в связи с недостаточностью аксонального транспорта).
Туннельные поражения нервов проявляются в первую очередь болью, чувствительными и вегетативными расстройствами. Двигательные нарушения развиваются лишь у одной трети больных и заключаются, как правило, в снижении мышечной силы, гипотрофии мышц, развитии контрактур. Прогноз восстановления функций при рано начатом лечении обычно благоприятный, однако это восстановление может происходить достаточно длительно, до нескольких месяцев. Кроме того, прогноз зависит от основного заболевания, на фоне которого развилась невропатия, от того, сохраняются ли профессиональные перегрузки конечности. В 30-40% случаев туннельные невропатии рецидивируют [Герман А.Г.и соавт., 1989].
На втором месте по частоте находятся травматические невропатии. Среди причин травматических невропатий прогностически наиболее благоприятными являются резаные ранения, при которых своевременное оперативное вмешательство обеспечивает хороший исход. Тракционные и огнестрельные травмы имеют худший прогноз, поскольку при них нередко измененными оказываются центральный отрезок нерва и нейрон спинномозговых центров, что существенно затрудняет регенерацию нерва. Разрушение нервного ствола на большом протяжении может наблюдаться также при электротравме, при химическом повреждении (случайное введение в нерв различных лекарственных веществ). Весьма неблагоприятно сопутствующее повреждению нерва нарушение кровообращения в конечности (кровотечение либо длительное наложение кровоостанавливающего жгута, тромбоз магистральной артерии), которое может приводить к развитию в мышцах, сухожилиях, суставных сумках, коже и подкожной клетчатке атрофирующего склерозирующего процесса с образованием контрактур. Препятствовать восстановлению движений могут также вторичные изменения в суставах и сухожилиях, которые развиваются вследствие растяжения связок и суставных сумок при пассивном свешивании конечностей в случае вялого паралича или пареза.
При невро- и плексопатиях, развившихся на фоне соматических заболеваний, вследствие иммунных, неопластических, инфекционных, токсических поражений и воздействий прогноз зависит от характера течения основного заболевания либо процесса.
7.2.3. Клинические и электрофизиоло-гические признаки восстановления нервных проводников
Дегенерация и регенерация нервных волокон
Нейроны являются несменяемой клеточной популяцией. Им свойственна только внутриклеточная физиологическая регенерация, заключающаяся в непрерывной смене структурных белков цитоплазмы. Отростки нейронов и соответственно периферические нервы обладают способностью к регенерации в случае их повреждения. При этом регенерации нервных волокон предшествуют явления дегенерации.
Дегенерация
Дегенерация - это патологический процесс, характеризующийся распадом миелиновой оболочки нервных волокон на ограниченных участках при относительной сохранности осевых цилиндров.
При дегенерации нерва происходят биохимические и биофизические изменения. Вес нерва и содержание в нем воды увеличиваются, двоякопреломляемость нарушается, понижается концентрация липоидов мякотной оболочки (сфигномиелина, цереброзидов, свободного холестерина). Изменяется активность различных ферментов.
Шванновская оболочка при дегенерации нерва также изменяется. Уже через 48 часов после травматизации нерва в шванновских клетках происходит набухание ядра, увеличение хроматина, разрастание цитоплазмы. Через 5—6 дней становятся заметны митозы, образуются синцитиальные протоплазматические тяжи с несколькими ядрами, так называемые бюнгеровские ленты. В конце первой недели полибласты скапливаются на поверхности дегенерирующих нервных волокон, затем внедряются в них, превращаясь в макрофаги.
При валлеровской дегенерации распад периферического отрезка поврежденного нерва начинается одновременно на всем протяжении (Б. С. Дойников, Р. Кахаль). Мнения авторов, изучавших процессы дегенерации в периферическом отрезке нерва, о непосредственной причине распада различны.
При повреждении тела нервной клетки она обычно погибает и фагоцитируется микроглиальными элементами. Фагоциты - клетки микроглии, которые обладают способностью поглощать погибшие части нейронов. Если повреждается (в результате перетяжки, травмы и пр.) аксоннервной клетки, то в теле соответствующего нейрона наступает ряд характерных изменений. Во-первых, наблюдаетсяхроматолиз, т.е. разрушение и растворение субстанции Ниссля, представляющей собой шероховатую эндоплазматическую сеть со скоплением рибосом в теле нейрона. Одновременно вследствие потери воды размеры тела нервной клетки и её ядра могут уменьшаться, цитоплазма вакуолизируется, ядро занимает краевое положение и меняет форму. Число нейрофибрилл в клетке уменьшается, они делаются тоньше и плохо различимыми. Центральный и периферический отрезки перерезанного аксона, его мякотная и безмякотная оболочки претерпевают распад; на некотором расстоянии от места поврежедения миелин растворяется. Вся эта картина получила название “первичной реакции Ниссля”, или ретроградной клеточной дегенерации, а для центрального и периферического отрезка аксона - травматической дегенерации. Особенно сложно протекают изменения в периферическом отрезке перерезанного аксона или, если речь идёт о нерве, в периферическом отрезке нерва. Эти изменения называются вторичной, или валлеровской, дегенерацией нервных волокон. Во время валлеровской дегенерации периферические отрезки аксонов, потерявшие связь с телом нервной клетки, являющейся их трофическим центром, распадаются и полностью дегенерируют. Миелиновая оболочка распадается; миелин собирается в капли, в которых иногда ещё можно проследить обломки периферических аксонов.
Аксональная дегенерация возникает, если в теле клетки нарушается синтез необходимых аксону веществ или при нарушении выработки энергии в митахондриях, вследствие чего прекращается быстрый антероградный аксоплазматический ток в аксоне. При аксональной дегенерации в нейроне наблюдается повышенный уровень кальция, который активизирует систему кальпаинов, способных запускать различные патологические процессы.
При аксональной дегенерации, прежде всего, страдает утолщенная часть аксона, затем дегенерация распространяется в проксимальном направлении. Сильнее всего повреждаются самые длинные аксоны, поэтому симптомы аксональной дегенерации ощущаются, прежде всего, в стопах и в кистях.
Дегенеративный процесс, распространившийся на тело клетки, приводит к ее гибели. Если же воздействие факторов, приведших к аксональной дегенерации, прекратилось до гибели клетки, то происходит восстановление аксонов. Регенерация происходит медленно и может занять от нескольких месяцев до года.
Дегенерация и регенерация нервных волокон. Регенерация нервных окончаний
В отличие от кожной раны, которая заживает по первичному натяжению, если ее края привести в соприкосновение, место пореза нерва первичным натяжением не срастается, даже в тех случаях, когда наложены немедленно эпиневральные швы.
Срастание разорванных частей нерва и восстановление его функций происходит путем длительного процесса регенерации нервных волокон, причем новообразованные нервные волокна растут из центрального участка поврежденного нерва. Такой точки зрения придерживаются отечественные и зарубежные нейрогистологи (Б. С. Дойников, Б. И. Лаврентьев, Р. Кахаль, И. Ф. Иванов, А. Н. Голиков). Немногочисленные представители теории клеточных цепей допускают возможность аутогенной регенерации за счет дифференцировки элементов шванновского синцития периферического отрезка.
Свойство нервов восстанавливать свою анатомическую целостность и физиологическую функцию проведения нервных возбуждений вырабатывалось в результате длительного развития и совершенствования организма животных. В этом приспособительном процессе, по-видимому, большую роль сыграли повреждающие факторы — травмы, в результате которых нарушалась анатомическая целостность нервных стволов конечностей и других периферических нервов. Именно нервные стволы и периферические смешанные нервы обладают наиболее высокой степенью регенерации, в то время как спинной и головной мозг этим свойством обладают значительно меньшей степени. Однако «нет никакого сомнения в том, что в поврежденном спинном и головном мозгу человека может происходить некоторая, хотя бы и абортивная регенерация» (У. Уитль, 1955). Опыт лечения больных с полным перерывом спинного мозга показал, что «спинной мозг человека не способен к регенерации, если у больного с полным перерывом этого органа позже в какой-то степени восстанавливается функция, то это следует объяснить неточным диагнозом, т. е. наличием фактически неполного разрыва спинного мозга, а не его регенерацией» (Мейфильд, 1953).
Известно, что нарушение иннервации внутренних органов неизбежно вызывает их дисфункции, а также снижает местный иммунитет. Например, ухудшение иннервации органов мочеполовой сферы вследствие остеохондроза может способствовать развитию таких заболеваний как простатит и аденома простаты у мужчин и гинекологические заболевания у женщин.
При повреждении нервов наблюдаются двигательные, чувствительные, вазомоторные, секреторные и трофические расстройства в разнообразных сочетаниях. Они могут быть сгруппированы в следующие синдромы:
1. Синдром полного нарушения проводимости нерва (синдром полного перерыва), который проявляется полным выпадением функции нерва. При этом отсутствуют боли спонтанные и при давлении на нервный ствол ниже уровня повреждения; сосудистые и трофические нарушения невелики.
2. Синдром раздражения, проявляется самопроизвольными болями различной интенсивности и развитием вегетативных и трофических расстройств.
3. Синдром сдавления нерва, проявляющийся:
а) быстрым прогрессированием симптомов раздражения и выпадения при гематоме или отеке, неправильном наложении гипсовой повязки, или бурным инфекционным процессом;
б) медленным развитием этой симптоматики при сдавлении рубцом.
4. Синдром частичного нарушения проводимости нерва - сочетание различных компонентов других симптомов.
5. Синдром регенерации, определяемый на основании анализа динамики симптомов на протяжении от 1 месяца после повреждения до нескольких лет. Этот синдром характеризуется медленно прогрессирующим, обычно частичным восстановлением двигательной и чувствительной функции нерва; одним из наиболее ранних признаков регенерации являются парестезии (ощущение ползания мурашек) в зоне анестезии, что указывает на прорастание молодых аксонов.
Процесс самовосстановления нервной системы состоит из двух составляющих: пластичности и регенерации.
Пластичность означает, что функции погибших нейронов берут на себя другие нервные клетки, увеличиваясь в размерах и формируя новые межнейронные связи, или дендриты, которые компенсируют утраченные функции. Регенерация означает образование новых нейронов, или нейрогенез.
А раз восстановление нервных клеток возможно в принципе, весь вопрос в том, чтобы найти средства, способные ускорить этот процесс. В частности, ученые лондонского университета Королевы Марии выяснили, что продукты с высоким содержанием жирных кислот Омега-3 ускоряют регенерацию нервных клеток и улучшают передачу нервных сигналов от головного и спинного мозга к другим нервным тканям.
Я выбрал эту тему, потому что медицинское и социальное значение травм периферической нервной системы весьма велико. Частота повреждений периферических нервов составляет от 1.5 до 10\% от всех травм. При этом, изолированное повреждение нерва наблюдается относительно редко: у 15\% повреждены магистральные сосуды, у 25-45.2\% трубчатые кости. Наиболее часто сочетанные повреждения отмечаются на верхних конечностях. При огнестрельных переломах костей конечностей нервы повреждаются в 10.6 (для бедренной) - 35.6\% (для плечевой). В 60\% случаев повреждения нервов приводят к развитию инвалидности.
Цель реферата: получить начальные знания о структуре нерва, изучить процессы дегенерации нервных волокон и нервных окончаний.
1.5.4. Регенерация периферических нервов
При разрушении участка периферического нерва в течение недели наступает восходящая дегенерация проксимальной (ближайшей к телу нейрона) части аксона с последующим некрозом как аксона, так и шванновской оболочки. На конце аксона формируется расширение (ретракционная колба). В дистальной части волокна после его перерезки отмечается нисходящая дегенерация с полным разрушением аксона, распадом миелина и последующим фагоцитозом детрита макрофагами и глией (рис. 1.5.7, 1.5.8).
Начало регенерации характеризуется сначала пролиферацией шванновских клеток, их передвижением вдоль распавшегося волокна с образованием клеточного тяжа, лежащего в эн-доневральных трубках. Таким образом, шван-новские клетки восстанавливают структурную целостность в месте разреза. Фибробласты также пролиферируют, но медленнее шванновских клеток. Указанный процесс пролиферации шванновских клеток сопровождается одновременной активацией макрофагов, которые первоначально захватывают, а затем лизируют оставшийся в результате разрушения нерва материал.
Следующий этап характеризуется прорастанием аксонов в щели, образованные шваннов-скими клетками, проталкиваясь от проксимального конца нерва к дистальному. При этом от ретракционной колбы в направлении дистальной части волокна начинают отрастать тонкие веточки (конусы роста). Регенерирующий аксон растет в дистальном направлении со скоростью 3—4 мм/сут вдоль лент из шванновских клеток (ленты Бюгнера), которые играют направляющую роль. В последующем наступает дифференциация шванновских клеток с образова-
Глава 1. Клетка и ткани
t t tt ft
J
Уолеров- Тер- Транс-ская миналь- нейронная ная
Рис. 1.5.7. Термины, используемые при описании
различных типов дегенерации нейронов и нервных
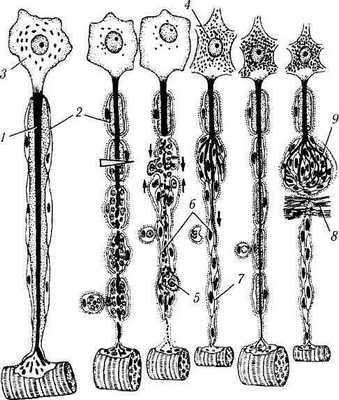
Рис. 1.5.8. Регенерация миелинового нервного волокна:
а — после перерезки нервного волокна проксимальная часть аксона (/) подвергается восходящей дегенерации, миелиновая оболочка (2) в области повреждения распадается, перикарион (3) нейрона набухает, ядро смещается к периферии, хромафильная субстанция (4) распадается; б—дистальная часть, связанная с иннервируемым органом, претерпевает нисходящую дегенерацию с полным разрушением аксона, распадом миелиновой оболочки и фагоцитозом детрита макрофагами (5) и глией; в — лем-моциты (6) сохраняются и митотически делятся, формируя тяжи — ленты Бюгнера (7), соединяющиеся с аналогичными образованиями в проксимальной части волокна (тонкие стрелки). Через 4—6 недель структура и функция нейрона восстанавливается, от проксимальной части аксона дистально отрастают тонкие веточки (жирная стрелка), растущие вдоль ленты Бюгнера; г — в результате регенерации нервного волокна восстанавливается связь с органом-мишенью и регрессирует ее атрофия; д — при возникновении преграды (8) на пути регенерирующего аксона компоненты нервного волокна формируют травматическую неврому (9), которая состоит из разрастающихся веточек аксона и леммоцитов
нием миелина и окружающей соединительной ткани. Коллатерали и терминали аксонов восстанавливаются в течение нескольких месяцев. Регенерация нервов происходит только при условии отсутствия повреждения тела нейрона, небольшом расстоянии между поврежденными концами нерва, отсутствии между ними соединительной ткани. При возникновении преграды на пути регенерирующего аксона развивается ампутационная нейрома. Регенерация нервных волокон в центральной нервной системе отсутствует.
Афанасьев Ю. И., Юрина Н. А. Гистология, ци тология и эмбриология. — М.: Медицина, 1999. — 744 с.
Билич Г., Катинас Г. С, Назарова JJ. В. Цито логия. — Спб., 1999.— 216 с.
Быков В. JI. Цитология и общая биология. — Спб.: СОТИС, 1999.— 519 с.
Быков В. JI. Частная гистология человека. — Спб.: СОТИС, 1997.— 300 с.
Дудел Д., Циммерман Л. Физиология чело века: В 4 т. / Пер. с англ.; под ред. Р. Шмидта и Г. Тевса. — Т. 2. — М.: Мир, 1985.— 240 с.
Луцик О. Д., 1ванова А.Й., Кабак К-С. Псто- лопя людини.—Льв1в: Мир, 1992. — С. 399.
Леей А., Сикевиц И. Структура и функция клет ки. — М.: Мир, 1971, —583 с.
Хэм А., Кормак Д. Гистология / Пер. с англ. — М.: Мир, 1982,— 1350 с.
Елисеев В. Г. Гистология.—М.: Медицина, 1972.— С. 612.
10. Toda K-, Fitzpatrlck Т. В. The origin of melano-somes, in Kawamura T. (eds): Biology of normal and Abnormal melanocytes // Tokyo, University of Tokyo press. — 1971. — P. 265—267.
U.Szabo G., Gerald А. В., Pathak M. A. Racial differences in the fate of meanosomes in human epidermis // Nature. — 1969.— Vol. 222. — P. 1081 — 1082.
Wolff K- Melanocyte-Keratinocyte interactions in vivo: The fate of melanosomes // Yale J Biol Med. — 1973.—Vol. 46.— P. 384—396.
Ramirez F., Pereira L. The fibrillins // Int J Bio- chem Cell Biol. — 1999. — Vol. 31. —P. 255—259.
Kielty С M., Shuttleworth С A. Fibrillin-contain- ing microfibrils: structure and function in health and di sease // Int J Biochem Cell Biol. — 1995. —Vol. 27. — p. 747—760.
Sakai L. Y., Keene D. R., Engvall E. Fibrillin, a new 350-kd glycoprotem, is a component of extracellu lar microfibrils // J Cell Biol. — 1986. — Vol. 103. — P. 2499.
Wright D. W., McDaniels С N., Swasdison S. Immunisation with undenatured bovine zonular fibrils results in monoclonal antibodies to fibrillin // Matrix Biol. — 1994.— Vol. 14.— P. 41—49.
17'. Thurmond F.A., Trotter J. A. Morphology and biomechanics of the microfibrillar network of sea cucumber dermis // J Exp Biol. — 1996. — Vol. 199. — P. 1817—1828.
McConnell СМ., DeMont M.E., Wright G. M. Microfibrils provide non-linear elastic behaviour in the abdominal artery of the lobster Homarus americanus // J. Physiol.—1997.—Vol. 499. — P. 513—526.
Rosenbloom J., Abrams W. R., Mecham B. Extra cellular matrix 4: the elastic fibre // FASEB J. — 1993.— Vol. 7.— P. 1208—1218.
Davis Е.С., Mecham R. P. Intracellular trafficking of tropoelastin // Matrix Biol. — 1998. — Vol. 17. — P. 245—254.
2\.Mayne R., Mayne P. R., Baker J. R. Fibrilllin-1 is the major protein present in bovine zonular fibrils // Invest Ophthalmol Vis Sci. — 1997. — Vol. 38.— P. 1399—1411.
Wheatley H.M., Traboulsi E.I., Flowers В. Е. Immunohistochemical localization of fibrillin in human ocular tissues // Arch Ophthalmol. — 1995. — Vol. 113. — P. 103—109.
Walacce R. N., Streeten B. W., Hanna R. B. Ro tary shadowing of elastic system microfibrils in the ocu lar zonule, vitreous, and ligamentum nuchae // Curr Eye Res. — 1991, — Vol. 10.— P. 99—108.
Wright D. W., Mayne R. Vitreous humor of chicken contains two fibrillar systems: an analysis of their structure // J. Ultra Mol. Struct. Res. — 1988. — Vol. 100. — P. 224—234.
Bishop P., Ayad S., Reardon A. Type VI collagen is present in human and bovine vitreous // Graefes Arch. Clin. Exp. Ophthalmol. — 1996.— Vol. 234.— P. 710—713.
Lee В., Godfrey M., Vitale E. Linkage of Mar- fan syndrome and a phenotypically related disorder to two different fibrillin genes // Nature.— 1991.— Vol. 352. — P. 330—338.
Zhang H., Apfelroth S. D., Ни W. Structure and expression of fibrillin-2, a novel microfibrillar compo nent preferentially located in elastic matrices // J. Cell Biol. — 1994.— Vol. 124.— P. 855—863.
Zhang H., Ни W., Ramirez F. Developmental expression of genes suggests heterogeneity of extracel lular microfibrils // J. Cell Biol. — 1995. — Vol. 129. — P. 1165—1176.
Mir S., Wheatley H. M., Maumenee-Hussels I. E. A comparative histologic study of the fibrillin microfibril lar system in the lens capsule of normal subjects and subjects with Marfan syndrome // Invest Ophthalmol Vis Sci. — 1998. — Vol. 39. — P. 84—93.
30. Farnsworth P. N. В., Burke P. Three-dimensional architecture of the suspensory apparatus of the lens of the rhesus monkey // Exp. Eye Res. — 1977. — Vol. 25. — P. 563.
3\.Pessier A. P., Potter K-A. Ocular pathology in bovine Marfan syndrome with demonstration of altered fibrillin immunoreactivity in explanted ciliary body cells // Lab Invest. — 1996. — Vol. 75. — P. 87—95.
Kielty СМ., Davies 5., Phillips J. Marfan synd rome expression and microfibrillar abnormalities in a family with predominant ocular defects // J. Med. Ge net. — 1994. — Vol. 31. —P. 1—5.
Izquierdo N. J., Traboulsi E., Enger C. Glauco ma in the Marfan syndrome // Trans Am Ophthalmol Soc. — 1992. — Vol. 90.— P. 111 — 122.
Izquierdo N. J., Traboulsi E. /., Enger C. Strabis mus in the Marfan syndrome // Am. J. Ophthalmol. — 1994.—Vol. 117.— P. 632—635.
Allen R., Straatsma В., Apt L. Ocular manifesta tions of the Marfan syndrome // Trans. Am. Acad. Oph thalmol. Otolaryngol. — 1967. — Vol. 71.— P. 18—38.
Ramsay M.S., Fine B.S., Shields J.A. The Mar fan syndrome. A histopathologic study of ocular findings //Am. J. Ophthalmol.—1973. —Vol. 76. —P. 103—116.
Cross H.E., Jensen A. D. Ocular manifestations in the Marfan syndrome and homocystinuria // Am J. Ophthalmol. — 1973. —Vol. 75. — P. 405—419.
Freissler K., Kuchle M., Naumann G. О. Н. Spon taneous dislocation of the lens in pseudoexfoliation syndrome // Arch Ophthalmol. — 1995. — Vol. 113.— P. 1095—11.
Schlutzer-Schrehardt U., Naumann G.O.H. A histopathologic study of zonular instability in pseudo- exfoliation syndrome // Am. J. Ophthalmol.— 1994. — Vol. 118.— P. 730—743.
Schlutzer-Schrehardt U., Stumer J. P., Reme С. Е. The fibrillin-containing microfibrillar network in the trabecular meshwork of normal and glaucomatous eyes // Invest Ophthalmol Vis Sci. — 1997. — Vol. 38.— P. 2117—2126.
Читайте также: