Морфофункциональные системы клетки. Покровная система клетки - плазмолемма.
Добавил пользователь Валентин П. Обновлено: 28.01.2026
Плазмолемма (plasmalemma), или внешняя клеточная мембрана, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку. Химический состав плазмолеммы. Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм и, таким образом,является самой толстой из клеточных мембран. Основу строения плазмолеммы составляет двойной слой липидных молекул билипидная мембрана, в которую местами включены молекулы белков, также имеется надмембранный слой гликокаликс, структурно связанный с белками и липидами билипидной мембраны, и в некоторых клетках имеется подмембранный слой.
Строение билипидной мембраны:Каждый монослой ее образован в основном молекулами фосфолипидов и, частично, холестерина. При этом в каждой липидной молекуле различают две части: гидрофильную головку и гидрофобные хвосты. Гидрофобные хвосты липидных молекул связываются друг с другом и образуют билипидный слой. Гидрофильные головки билипидного слоя соприкасаются с внешней или внутренней средой. Билипидная мембрана, а точнее ее глубокий гидрофобный слой, выполняет барьерную функцию, препятствуя проникновению воды и растворенных в ней веществ, а также крупных молекул и частиц.
Белковые молекулы встроены в билипидный слой мембраны локально и не образуют сплошного слоя.
По локализации в мембране белки подразделяются на:
интегральные пронизывают всю толщу билипидного слоя;
полуинтегральные включающиеся только в монослой липидов (наружный или внутренний);
прилежащие к мембране, но не встроенные в нее.
Функции плазмолеммы. Эта мембрана выполняет ряд важнейших клеточных функций, ведущими из которых являются функция р а з г р а н и ч е н и я цитоплазмы с внешней средой, функции р е ц е п ц и и и т р а н с п о р т а различных веществ как внутрь клетки, так и из нее.
По выполняемой функции белки плазмолеммы подразделяются на:
Находящиеся на внешней поверхности плазмолеммы белки, в также гидрофильные головки липидов обычно связаны цепочками углеводов и образуют сложные полимерные молекулы гликопротеиды и гликолипиды. Именно эти макромолекулы и составляют надмембранный слой — гликокаликс. В неделящейся клетке имеется подмембранный слой, образованный микротрубочками и микрофиламентами.
Значительная часть поверхностных гликопротеидов и гликолипидов выполняют в норме рецепторные функции, воспринимают гормоны и другие биологически активные вещества. Такие клеточные рецепторы передают воспринимаемые сигналы на внутриклеточные ферментные системы, усиливая или угнетая обмен веществ и тем самым оказывают влияние на функции клеток. Клеточные рецепторы, а возможно и другие мембранные белки, благодаря своей химической и пространственной специфичности, придают специфичность данному типу клеток данного организма и составляюттрансплантационные антигеныилиантигены гистосовместимости.
Помимо барьерной функции, предохраняющей внутреннюю среду клетки, плазмолемма выполняет транспортные функции, обеспечивающие обмен клетки с окружающей средой.
Различают следующие способы транспорта веществ:
пассивный транспорт способ диффузии веществ через плазмолемму (ионов, некоторых низкомолекулярных веществ) без затраты энергии;
активный транспорт веществ с помощью белков-переносчиков с затратой энергии (аминокислот, нуклеотидов и других);
везикулярный транспорт через посредство везикул (пузырьков), который подразделяется на эндоцитоз транспорт веществ в клетку, и экзоцитозтранспорт веществ из клетки.
В свою очередь эндоцитоз подразделяется на:
фагоцитоз захват и перемещение в клетку крупных частиц (клеток или фрагментов, бактерий, макромолекул и так далее);
пиноцитоз перенос воды и небольших молекул.
Процесс фагоцитоза подразделяется несколько фаз:
адгезия (прилипание) объекта к цитолемме фагоцитирующей клетки;
поглощение объекта путем образования вначале углубления (инвагинации), а затем и образования пузырьков — фагосомы и передвижения ее в гиалоплазму
Характеристика надмембранного слоя (гликокаликса) : сложные полимерные макромолекулы, образованные цепочками углеводов, связанными с находящимися на внешней поверхности плазмолеммы белками и гидрофильными головками липидов. Значительная часть поверхностных гликопротеидов и гликолипидов выполняет в норме рецепторные функции. Характеристика подмембранного слоя клеточной оболочки: Образован периферическим (кортикальным) слоем цитоплазмы и содержащимися в нём элементами цитоскелета клетки, включающим актиновые микрофиламенты, промежуточные филаменты и микротрубочки. Сокращение сети микрофиламентов, связанных с белками плазмолеммы, способствует формированию псевдоподий и выростов цитоплазмы, перемещению клетки в пространстве.
8.Функциональная и структурная характеристика различных видов соединений. Простые соединения. Сложные соединения: плотные, соединения, щелевые соединения (нексусы), промежуточные соединения, десмосомы, пальцевидные соединения.
Межклеточные соединения делятся на п р о с т ы е и с л о ж н ы е . Простое межклеточное соединение— сближение плазмолемм соседних клеток на расстояние 15—20 нм (рис. 8). При этом происходит взаимодействие слоев гликокаликса соседних клеток. Гликопротеиды соседних клеток при образовании простого контакта «узнают» клетки одного типа. Наличие этих белков-рецепторов (кадгерины, интегрины и др.) характерно для определенных тканей. Они реагируют только с соответствующими им клетками.
Пальцевидные соединения (интердигитации) образуются за счет взаимной инвагинации (впячивания) обеих плазмолемм в начале в одном, а затем в другом. Это один из трех видов контактов между кардиомиоцитами.
Сложные межклеточные соединения представляют собой небольшие парные специализированные участки плазматических мембран двух соседних клеток. Они подразделяются на запирающие (изолирующие), сцепляющие (заякоривающие) и коммуникационные (объединяющие) контакты. К з а п и р а ю щ и м (изолирующим) относится плотный контакт . В этом соединении принимают участие специальные интегральные белки, расположенные на поверхности соседних клеток, образующие подобие ячеистой сети.Эта ячеистая сеть окружает в виде пояска весь периметр клетки, соединяясь с такой же сетью на поверхности соседних клеток. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды. Этот тип соединений характерен для клеток однослойных эпителиев и эндотелия. К с ц е п л я ю щ и м , или заякоривающим, соединениям относятся адгезивный (сцепляющий) поясок и десмосомы. Общим для этой группы соединений является то, что к участкам плазматических мембран со стороны цитоплазмы подходят фибриллярные элементы цитоскелета, которые как бы заякориваются на их поверхности. Адгезивный поясок(промежуточный контакт)-опоясывает апикальную часть клетки однослойных эпителиев.Клетки связаны интегральными гликопротеидами,к которым примыкает слой примембранных белков.
Факальный контакт характерен для фибробластов.Соединяется не с соседней клеткой,а с элементами внеклеточного субстрата.В образовании этого контакта участвуют микрофиламенты.
Десмосомы-парные структуры,диаметр-0,5 мкм.Прилежит слой белков.В состав входят десмоплакины.С внешней стороны плазмолеммы соединяются с помощью десмоглеинов. Функция:механическая связь между клетками.(В клетках эпителиев,сердечных и гладких мышц)
Полудесмосомы-связь эпителиальных клеток с базалбной мембраной.
Коммуникационные соединения(щелевые контакты и синапсы):
Щелевые контакты(нексусы)- 0,5-3 мкм.В структуре плазмолемм соседних клеток располагаются коннексоны(Все ткани).Функция:перенос ионов и мелких молекул от клетки к клетке.
Синапсы-характерны для нервной ткани.Встречаются между двумя нейронами.Участки контактов двух клеток,специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому.
Лекция № 6. Эукариотическая клетка: цитоплазма, клеточная оболочка, строение и функции клеточных мембран
Цитоплазма — обязательная часть клетки, заключенная между плазматической мембраной и ядром; подразделяется на гиалоплазму (основное вещество цитоплазмы), органоиды (постоянные компоненты цитоплазмы) и включения (временные компоненты цитоплазмы). Химический состав цитоплазмы: основу составляет вода (60-90% всей массы цитоплазмы), различные органические и неорганические соединения. Цитоплазма имеет щелочную реакцию. Характерная особенность цитоплазмы эукариотической клетки — постоянное движение (циклоз). Оно обнаруживается, прежде всего, по перемещению органоидов клетки, например хлоропластов. Если движение цитоплазмы прекращается, клетка погибает, так как, только находясь в постоянном движении, она может выполнять свои функции.
Гиалоплазма (цитозоль) представляет собой бесцветный, слизистый, густой и прозрачный коллоидный раствор. Именно в ней протекают все процессы обмена веществ, она обеспечивает взаимосвязь ядра и всех органоидов. В зависимости от преобладания в гиалоплазме жидкой части или крупных молекул, различают две формы гиалоплазмы: золь — более жидкая гиалоплазма и гель — более густая гиалоплазма. Между ними возможны взаимопереходы: гель превращается в золь и наоборот.
Функции цитоплазмы:
- объединение всех компонентов клетки в единую систему,
- среда для прохождения многих биохимических и физиологических процессов,
- среда для существования и функционирования органоидов.
Клеточные оболочки
Клеточные оболочки ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной (синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом (образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой (образован целлюлозой).
Строение мембран
Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны. Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде.
Помимо липидов в состав мембраны входят белки (в среднем ≈ 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки (расположены на наружной или внутренней поверхности липидного бислоя), 2) полуинтегральные белки (погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки (пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).
Строение мембраны: А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Молекулы белков, углеводов и липидов подвижны, способны перемещаться в плоскости мембраны. Толщина плазматической мембраны — примерно 7,5 нм.
Функции мембран
Мембраны выполняют такие функции:
- отделение клеточного содержимого от внешней среды,
- регуляция обмена веществ между клеткой и средой,
- деление клетки на компартаменты («отсеки»),
- место локализации «ферментативных конвейеров»,
- обеспечение связи между клетками в тканях многоклеточных организмов (адгезия),
- распознавание сигналов.
Важнейшее свойство мембран — избирательная проницаемость, т.е. мембраны хорошо проницаемы для одних веществ или молекул и плохо проницаемы (или совсем непроницаемы) для других. Это свойство лежит в основе регуляторной функции мембран, обеспечивающей обмен веществ между клеткой и внешней средой. Процесс прохождения веществ через клеточную мембрану называют транспортом веществ. Различают: 1) пассивный транспорт — процесс прохождения веществ, идущий без затрат энергии; 2) активный транспорт — процесс прохождения веществ, идущий с затратами энергии.
При пассивном транспорте вещества перемещаются из области с более высокой концентрацией в область с более низкой, т.е. по градиенту концентрации. В любом растворе имеются молекулы растворителя и растворенного вещества. Процесс перемещения молекул растворенного вещества называют диффузией, перемещения молекул растворителя — осмосом. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Можно выделить следующие виды пассивного транспорта: 1) простая диффузия — транспорт веществ непосредственно через липидный бислой (кислород, углекислый газ); 2) диффузия через мембранные каналы — транспорт через каналообразующие белки (Na + , K + , Ca 2+ , Cl - ); 3) облегченная диффузия — транспорт веществ с помощью специальных транспортных белков, каждый из которых отвечает за перемещение определенных молекул или групп родственных молекул (глюкоза, аминокислоты, нуклеотиды); 4) осмос — транспорт молекул воды (во всех биологических системах растворителем является именно вода).
Необходимость активного транспорта возникает тогда, когда нужно обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется особыми белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ. К активному транспорту относят: 1) Na + /К + -насос (натрий-калиевый насос), 2) эндоцитоз, 3) экзоцитоз.
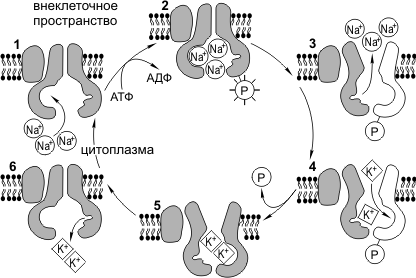
Работа Na + /К + -насоса. Для нормального функционирования клетка должна поддерживать определенное соотношение ионов К + и Na + в цитоплазме и во внешней среде. Концентрация К + внутри клетки должна быть значительно выше, чем за ее пределами, а Na + — наоборот. Следует отметить, что Na + и К + могут свободно диффундировать через мембранные поры. Na + /К + -насос противодействует выравниванию концентраций этих ионов и активно перекачивает Na + из клетки, а K + в клетку. Na + /К + -насос представляет собой трансмембранный белок, способный к конформационным изменениям, вследствие чего он может присоединять как K + , так и Na + . Цикл работы Na + /К + -насоса можно разделить на следующие фазы: 1) присоединение Na + с внутренней стороны мембраны, 2) фосфорилирование белка-насоса, 3) высвобождение Na + во внеклеточном пространстве, 4) присоединение K + с внешней стороны мембраны, 5) дефосфорилирование белка-насоса, 6) высвобождение K + во внутриклеточном пространстве. На работу натрий-калиевого насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки. За один цикл работы насос выкачивает из клетки 3Na + и закачивает 2К + .
Эндоцитоз — процесс поглощения клеткой крупных частиц и макромолекул. Различают два типа эндоцитоза: 1) фагоцитоз — захват и поглощение крупных частиц (клеток, частей клеток, макромолекул) и 2) пиноцитоз — захват и поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Явление фагоцитоза открыто И.И. Мечниковым в 1882 г. При эндоцитозе плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму структур, отграниченных от цитоплазмы одиночной мембраной. К фагоцитозу способны многие простейшие, некоторые лейкоциты. Пиноцитоз наблюдается в эпителиальных клетках кишечника, в эндотелии кровеносных капилляров.
Экзоцитоз — процесс, обратный эндоцитозу: выведение различных веществ из клетки. При экзоцитозе мембрана пузырька сливается с наружной цитоплазматической мембраной, содержимое везикулы выводится за пределы клетки, а ее мембрана включается в состав наружной цитоплазматической мембраны. Таким способом из клеток желез внутренней секреции выводятся гормоны, у простейших — непереваренные остатки пищи.
Перейти к лекции №5 «Клеточная теория. Типы клеточной организации»
Перейти к лекции №7 «Эукариотическая клетка: строение и функции органоидов»
Строение и функции плазмолеммы (цитолеммы)
1. Цитология наука о строении, развитии и жизнедеятельности клеток. Следовательно, цитология изучает закономерности структурно-функциональной организации первого (клеточного) уровня организации живой материи. Клетка является наименьшей единицей живой материи, обладающей самостоятельной жизнедеятельностью и способностью к самовоспроизведению. Субклеточные образования (ядро, митохондрии и другие органеллы) хотя и являются живыми структурами, но не обладают самостоятельной жизнедеятельностью.
Клетка элементарная единица живого, состоящая из цитоплазмы и ядра и являющаяся основой строения, развития и жизнедеятельности всех животных и растительных организмов.
Основные компоненты клетки:
По соотношению ядра и цитоплазмы (ядерно-цитоплазматическое отношение) клетки подразделяются на:
· клетки ядерного типа объем ядра преобладает над объемом цитоплазмы;
· клетки цитоплазматического типа цитоплазма преобладает над ядром.
По форме клетки бывают:
· круглыми (клетки крови);
· кубическими или цилиндрическими (клетки разных эпителиев);
· отростчатыми (нервные клетки) и другие.
Большинство клеток содержат одно ядро, однако могут быть в одной клетке 2, 3 и более ядер многоядерные клетки. В организме имеются структуры (симпласты, синтиций), содержащие несколько десятков или даже сотен ядер. Однако эти структуры образуются или в результате слияния отдельных клеток (симпласты), или в результате неполного деления клеток (синцитий). Морфология этих структур будет рассмотрена при изучении тканей.
Структурные компоненты цитоплазмы животной клетки:
· Плазмолемму, окружающую цитоплазму, нередко рассматривают как одну из органелл цитоплазмы.
Плазмолемма оболочка животной клетки, ограничивающая ее внутреннюю среду и обеспечивающая взаимодействие клетки с внеклеточной средой.
Плазмолемма имеет толщину около 10 нм, и состоит на 40 % из липидов, на 5—10 % из углеводов (в составе гликокаликса), и на 50—55 % из белков.
Функции плазмолеммы:
· рецепторная или антигенная;
· образование межклеточных контактов.
Основу строения плазмолеммы составляет двойной слой липидных молекулбилипидная мембрана, в которую местами включены молекулы белков, также имеется надмембранный слой гликокаликс, структурно связанный с белками и липидами билипидной мембраны, и в некоторых клетках имеется подмембранный слой.
Строение билипидной мембраны
Каждый монослой ее образован в основном молекулами фосфолипидов и, частично, холестерина. При этом в каждой липидной молекуле различают две части: гидрофильную головку и гидрофобные хвосты. Гидрофобные хвосты липидных молекул связываются друг с другом и образуют билипидный слой. Гидрофильные головки билипидного слоя соприкасаются с внешней или внутренней средой. Билипидная мембрана, а точнее ее глубокий гидрофобный слой, выполняет барьерную функцию, препятствуя проникновению воды и растворенных в ней веществ, а также крупных молекул и частиц.
На электроннограмме в плазмолемме четко определяются три слоя наружный и внутренний электронноплотные, промежуточный с низкой электронной плотностью.
Белковые молекулы встроены в билипидный слой мембраны локально и не образуют сплошного слоя. По локализации в мембране белки подразделяются на:
· интегральные пронизывают всю толщу билипидного слоя;
· полуинтегральные включающиеся только в монослой липидов (наружный или внутренний);
· прилежащие к мембране, но не встроенные в нее.
По выполняемой функции белки плазмолеммы подразделяются на:
Значительная часть поверхностных гликопротеидов и гликолипидов выполняют в норме рецепторные функции, воспринимают гормоны и другие биологически активные вещества. Такие клеточные рецепторы передают воспринимаемые сигналы на внутриклеточные ферментные системы, усиливая или угнетая обмен веществ и тем самым оказывают влияние на функции клеток. Клеточные рецепторы, а возможно и другие мембранные белки, благодаря своей химической и пространственной специфичности, придают специфичность данному типу клеток данного организма и составляют трансплантационные антигены или антигены гистосовместимости.
Различают следующие способы транспорта веществ:
· пассивный транспорт способ диффузии веществ через плазмолемму (ионов, некоторых низкомолекулярных веществ) без затраты энергии;
· активный транспорт веществ с помощью белков-переносчиков с затратой энергии (аминокислот, нуклеотидов и других);
· везикулярный транспорт через посредство везикул (пузырьков), который подразделяется на эндоцитоз транспорт веществ в клетку, и экзоцитозтранспорт веществ из клетки.
В свою очередь эндоцитоз подразделяется на:
· фагоцитоз захват и перемещение в клетку крупных частиц (клеток или фрагментов, бактерий, макромолекул и так далее);
· пиноцитоз перенос воды и небольших молекул.
Процесс фагоцитоза подразделяется несколько фаз:
· адгезия (прилипание) объекта к цитолемме фагоцитирующей клетки;
· поглощение объекта путем образования вначале углубления (инвагинации), а затем и образования пузырьков — фагосомы и передвижения ее в гиалоплазму
Морфофункциональная характеристика клетки: общие понятия о клетке и ее функциях; плазмолемма, цитоплазма и ее компоненты, органеллы.
Клетка- элементарная биологическая система, способная к самообновлению, самовоспроизведению и развитию. Клеточные структуры лежат в основе строения растений и животных. Каким бы многообразным ни представлялось строение организмов, в основе его лежат сходные структуры - клетки.
Клетка обладает всеми свойствами живой системы:
- осуществляет обмен веществом и энергией;
- размножается и передает по наследству свои признаки;
- реагирует на внешний сигналы (раздражители);
Живое содержимое клетки, протоплазма, отделяется от окружающей среды плазматической мембраной (плазмолеммой) и, в некоторых случаях, прочной клеточной стенкой. Протоплазма представляет собой коллоидную неоднородную массу с множеством различных органелл и параплазматических включений. Последние только условно причисляются к живой протоплазме и содержат вещества, подлежащие накоплению или выделению.
I. Цитология с эмбриологией
1.Структурно- химический состав и молекулярная организация плазмолеммы
Плазмолемма, или внешняя клеточная мембрана- поверхностная периферическая структура, ограничивающая клетку снаружи и обеспечивающая ее непосредственную связь с внеклеточной средой.
Функции плазмолеммы:
1) разграничительная (барьерная);
5) образование межклеточных контактов
Химический состав веществ плазмолеммы: белки, липиды, углеводы. Основу плазмолеммы составляет липо-протеиновый комплекс. Она имеет толщину около 10 нм и, таким образом, является самой толстой из клеточных мембран.
Строение плазмолеммы:
1) двойной слой липидных молекул, составляющий основу плазмолеммы, в которую местами включены молекулы белков;
2) надмембранный слой гликокаликс (Находящиеся на внешней поверхности плазмолеммы белки и гидрофильные головки липидов связаные с цепочками углеводов).
Толщина этого слоя около 3—4 нм, он обнаружен практически у всех животных клеток, но степень его выраженности различна. Гликокаликс представляет собой ассоциированный с плазмолеммой гликопротеиновый комплекс, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы В гликокаликсе могут располагаться белки, не связанные непосредственно с билипидным слоем.
Как правило, это белки-ферменты, участвующие во внеклеточном расщеплении различных веществ, таких как углеводы, белки, жиры и др.
3) подмембранный слой. Подмембранный (кортикальный) слой плазмолеммы образован упорядоченной сетью поперечно связанных белковых нитей из актина и актинсвязанных белков (прежде всего филамина), которая выстилает изнутри поверхность плазматической мембраны.
Молекулярное строение плазмолеммы описывается жидкостно-мозаичной моделью, согласно которой она состоит из липидного бислоя, в который погружены молекулы белков.
Липидный бислой представлен преимущественно молекулами фосфолипидов (таких как лецитин и цефалин), состоящими из двух длинных неполярных (гидрофобных) цепей жирных кислот и полярной (гидрофильной) головки. В состав большинства мембран входит также холестерин.
Мембранные белки составляют более 50% массы мембран. Они удерживаются в липидном бислое за счет гидрофобных взаимодействий с молекулами липидов. Белки обеспечивают специфические свойства мембраны и играют различную биологическую роль: структурных молекул, ферментов, переносчиков и рецепторов. Мембранные белки подразделяются на 2 группы: интегральные и периферические.
Периферические белки непрочно связаны с поверхностью мембраны и обычно находятся вне липидного бислоя.
Интегральные белки либо полностью, либо частично погружены в липидный бислой; либо часть белков целиком пронизывает всю мембрану (трансмембранные белки).
2.Надмембранный (гликокаликс) и подмембранный (кортикальный) компоненты плазмолеммы. Особенности строения и функции. Мембранные рецепторы
Гликокаликс — «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов, гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции, а также участвует в обеспечении избирательности транспорта веществ и пристеночном (примембранном) пищеварении. Наличие гликокаликса характерно для клеток животных (в отличие от прокариотов, растений и грибов, где его нет.)
Гликокаликс представляет собой молекулярное сито, пропускающего или не пропускающего молекулы, в зависимости от их величины, заряда и других параметров. В слое гликокаликса располагаются пищеварительные ферменты, как поступающие туда из полости кишечника, так и синтезированные энтероцитами (общее название ряда клеток эпителия кишечника). Толщина гликокаликса равна приблизительно 15—40 нм на боковой поверхности энтероцита и 50—100 нм — на апикальной. Гликокаликс, микроворсинки и апикальная мембрана вместе называются исчерченной каёмкой.
Кортикальный слой образован микротрубочками и сократимыми, актиновыми микрофиламентами, которые являются частью цитоскелета клетки. Подмембранный слой обеспечивает поддержание формы клетки, изменения клеточной поверхности, участвует в эндо- и экзоцитозе, секреции, движении.
Мембранные рецепторы являются преимущественно гликопротеинами,
которые расположены на поверхности плазмолеммы клеток и обладают
способностью высокоспецифически связываться со своими лигандами. Они
выполняют ряд функций:
(1) регулируют проницаемость плазмолеммы
(2) регулируют поступление некоторых молекул в клетку
(3) действуют как датчики, превращая внеклеточные сигналы во
(4) связывают молекулы внеклеточного матрикса с цитоскелетом, эти
рецепторы, называемые интегринами, играют важную роль в формировании
контактов между клетками и клеткой и компонентами межклеточного вещества.
Рецепторы, связанные с каналами, взаимодействуют с сигнальной
молекулой (нейромедиатором), которая временно открывает или закрывает
воротный механизм, в результате чего инициируется или блокируется
транспорт ионов через канал.
Каталитические рецепторы включают внеклеточную часть (собственно
рецептор) и цитоплазматическую часть, которая функционирует как
протеинкиназа (посредством таких рецепторов на клетки воздействуют
инсулин и некоторые факторы роста).
Рецепторы, связанные с G-белками - трансмембранные белки, ассоциированные с ионным каналом или ферментом, - состоят из рецептора,
взаимодействующего с сигнальной молекулой (первый посредник), и G-белка
(гуанозин трифосфат-связывающего регуляторного белка), который передает сигнал на связанный с мембраной фермент или ионный канал, вследствие чего активируется второй внутриклеточный посредник - чаще всего циклический АМФ (цАМФ) или Са 2+ .
Читайте также: