Ранний эмбриогенез. Развитие зародыша. Амниотический пузырь. Амниотическая жидкость. Оболочки.
Добавил пользователь Morpheus Обновлено: 09.01.2026
ПЛОДНЫЕ ОБОЛОЧКИ — оболочки, возникающие на ранней стадии эмбриогенеза, развивающиеся в соответствии с потребностями зародыша (плода) и обеспечивающие условия его развития. Плодные оболочки являются составной частью последа (см.). К плодным оболочкам относятся амнион и хорион, развивающиеся из внезародышевых эмбриональных компонентов, а также слизистая оболочка матки, подвергающаяся во время беременности преобразованию в децидуальную оболочку (см.).
Амнион (греч, amnion чаша для жертвенной крови; син.: водная оболочка, амниотическая оболочка) — внутренняя Плодная оболочка, образующая полость, содержащую околоплодные воды (см.), в которой развивается плод. Зачаток амниона образуется из внезародышевой эктодермы и внезародышевой мезодермы (см. Зародышевые листки). Из эктодермы развивается эпителий амниона, из мезодермы — его соединительнотканная основа.
Макроскопически амнион представляет собой тонкую, почти прозрачную оболочку, которая прилегает к хориону, выстилает внутреннюю поверхность плаценты, переходит на пуповину, покрывая ее в виде футляра и сливается в области пупка с внешними покровами зародыша. Микроскопически амнион состоит из 5 слоев: внутренняя его поверхность покрыта эпителием, под которым располагаются базальная мембрана, компактный слой, фибробласты, спонгиозный слой. Эпителий амниона на ранних стадиях развития плоский, в последующем он становится однорядным кубическим и цилиндрическим. При этом внеплацентарная зона амниона выстлана кубическим эпителием, а плацентарная — цилиндрическим эпителием, где наряду с цилиндрическими клетками встречаются пузырькообразные. С функцией пузырькообразных клеток связывают в основном образование околоплодных вод. Клетки эпителия амниона в период секреции становятся выше, в их цитоплазме образуются вакуоли, к-рые сливаются, оттесняют к периферии ядро, вызывают разрыв оболочки клетки; содержимое вакуолей вместе с частью цитоплазмы переходит в полость амниона. Разрушенные клетки слущиваются и замещаются новыми. Цитоплазма клеток эпителия амниона содержит липиды, полисахариды, гликозаминогликаны, а также многочисленные ферменты типа липазы, пептидаз, трипсина, а также гликолитические ферменты. Базальная мембрана располагается под эпителием в виде узкой эозинофильной бесклеточной массы. Компактный слой представлен гомогенной массой, лишенной клеток, отличающейся от базальной мембраны более бледной окраской. Слой фибробластов самый толстый и состоит из фибробластов, располагающихся в густой сети коллагеновых, ретикулярных волокон и межклеточного вещества; иногда в нем встречаются гистиоциты. Спонгиозный слой амниона связан посредством соединительнотканных волокон и межклеточного вещества с гладким хорионом. Амнион содержит антигены (А и В), соответствующие групповым антигенам, содержащимся в крови плода.
Хорион (греч, chorion послед) — оболочка плодного происхождения, возникающая на ранней стадии эмбриогенеза (стадии морулы и бластоцисты) на основе трофобласта и мезобласта. После имплантации оплодотворенной яйцеклетки на поверхности трофобласта (см.) появляются первичные ворсины, к 12-му дню внутриутробного развития в них появляются элементы мезенхимы — начало возникновения хориона. К концу 2-й нед. беременности образуются вторичные ворсины хориона, погруженные в межворсинчатое пространство. Ворсины хориона имеют двухслойный эпителиальный покров: синцитий и цитотрофобласт (слой клеток Лангханса); строма их состоит из мезенхимы, в которой находятся фибробласты и кровеносные сосуды. В течение первых недель беременности ворсины хориона распределяются по поверхности плодного яйца равномерно. С конца II месяца беременности ворсины в области капсульной децидуальной оболочки постепенно атрофируются и исчезают, хорион в этой области становится гладким. Ворсины в области базальной децидуальной оболочки разрастаются, формируя плаценту (см.). Гладкий хорион внутренней поверхностью прилегает к амниону, наружной — к децидуальной оболочке. В нем различают четыре слоя: клеточный, ретикулярный, псевдобазальную мембрану, трофобласт.
Клеточный слой гладкого хориона хорошо дифференцируется в ранние сроки беременности, в поздние сроки в П. о. он нередко отсутствует. Ретикулярный слой является наиболее прочным, в нем содержатся фибробласты и клетки Кащенко — Хофбауэра. Псевдобазальная мембрана—тонкий однородный слой, не содержащий клеток. Трофобласт неотчетливо отделяется от прилежащей децидуальной оболочки. По данным Ю. В. Гулъкевича, клетки трофобласта располагаются в несколько слоев (5—10), отличаются полиморфизмом, цитоплазма их содержит значительное количество рибонуклеопротеидов, высокомолекулярные полисахариды, гликоген, а также щелочную фосфатазу, неспецифические эстеразы и другие ферменты. Структура и функциональная активность трофобласта сохраняются до конца беременности. Полагают, что трофобласт гладкого хориона синтезирует хорионический гонадотропин (см.).
В соединительной ткани гладкого хориона и прилежащей к нему децидуальной оболочке найдены кислые гликозаминогликаны, напр, гиалуроновая к-та и хондроитинсульфаты, а также различные гликопротеиды и гликоген. Групповые антигены плода в гладком хорионе обычно отсутствуют, поэтому ткань его не оказывает иммунизирующего влияния на организм беременной.
Функциональное значение
Важнейшей функцией эпителия амниона является секреция околоплодных вод, определяющих важнейшие условия развития плода, а также выведение продуктов его обмена, попадающих в околоплодные воды. Резорбция вод происходит через эпителий амниона, далее жидкость проникает в спонгиозный слой, граничащий с гладким хорионом, и удаляется через ткань гладкого хориона. Секреция и резорбция околоплодных вод определяют образование вод в количестве, необходимом для обеспечения жизнедеятельности плода на всех стадиях его развития. Удаление вместе с околоплодными водами продуктов метаболизма указывает на участие П. о. в обеспечении гомеостаза развивающегося плода.
Хорион выполняет трофическую, выделительную, дыхательную и защитную функции.
Данные об активности ферментных систем в амнионе и гладком хорионе указывают также на наличие процессов параплацентарного обмена, осуществляемого через П. о.
П. о. вместе с околоплодными водами играют существенную роль в течении родов (см.). При родовых схватках околоплодные воды устремляются в сторону внутреннего отверстия шейки матки, где располагается нижний отдел П. о. Эта часть оболочек, внедряющаяся вместе с околоплодными водами в канал шейки матки, носит название плодного пузыря.
Плодный пузырь имеет обычно сферическую форму и разрывается в родах в конце периода раскрытия шейки матки.
Патология
С изменением функций П. о. связано нарушение процессов секреции и резорбции околоплодных вод, преимущественно амниона. Оно может привести к недостаточной продукции вод — маловодию (см.) или избытку их — гидроамниону (см. Многоводие). Эти нарушения так же, как и амниотические сращения, перетяжки между амнионом и различными частями плода (см. Амниотические нити, перетяжки, сращения), нередко сопутствуют порокам развития плода (см. Пороки развития). К нередким видам патологии П. о. относят воспалительный процесс, возникающий в амнионе и хорионе (хориоамнионит), который характеризуется симптомами интоксикации и гноевидными выделениями из родовых путей.
При чрезмерной плотности П. о. плодный пузырь приобретает цилиндрическую или грушевидную форму (под напором околоплодных вод), разрыв оболочек может быть запоздалым, что иногда может способствовать преждевременной отслойке плаценты. Плоский плодный пузырь, возникающий вследствие малого количества передних околоплодных вод, тормозит процесс раскрытия шейки матки в первом периоде родов, а также своевременный разрыв оболочек плодного пузыря.
К нередким видам акушерской патологии относится преждевременный (до начала родовой деятельности) и ранний (до полного раскрытия шейки матки) разрыв плодных оболочек (см. Преждевременное отхождение вод). Разрыв оболочек плодного пузыря (своевременный, несвоевременный) зависит от структуры
П. о., степени повышения внутриматочного давления, а также скорости и характера формирования самого плодного пузыря.
Библиография:
Гулькевич Ю. В., Маккавеева М. Ю. и Никифоров Б. И. Патология последа человека и ее влияние на плод, Минск, 1968; Жемкова 3. П. и Tопчиева О. И. Клинико-морфологическая диагностика недостаточности плаценты, с. 9 и др., Л., 1973; Dallenbach-Hellweg С. a. Nette G. Morphological and histochemical observations on trophoblast and decidua of the basal plate of the human placenta at term, Amer. J. Anat., v. 115, p. 309, 1964; Fox H. Pathology of the placenta, L. a. o., 1978; Handbuch der speziellen pathologischen Anatomie und Histologie, hrsg. v. F. Henke u. O. Lubarsch, Bd 7, T. 5, B. u. a., 1967.
Общая характеристика эмбрионального развития: зигота, дробление, гаструляция, гисто- и органогенез. Зародышевые оболочки плода. Взаимоотношение материнского организма и плода.
В онтогенезе различают два периода — эмбриональный и постэмбриональный. Для высших животных и человека принято деление на пренатальный, или антенатальный (до рождения), и постнатальный (после рождения). Предложено также выделить предзиготный период, предшествующий образованию зиготы.
Предзиготный период развития связан с образованием гамет (гаметогенез). Процессы, характеризующие овогенез, приводят к образованию гаплоидного набора хромосом и формированию сложных структур в цитоплазме. В яйцеклетках происходит накопление желтка. В зависимости от количества желтка и характера его распределения различают яйца трех основных типов: изолецитальные, телолецитальные и центролецитальные.
Изолецитальные яйца содержат немного желтка, и он распределен равномерно по всей клетке. Такие яйца встречаются у иглокожих низших хордовых, млекопитающих. Телолецитальныеяйца характерны для моллюсков, земноводных, рептилий, птиц, содержат большое количество желтка, сосредоточенного на одном из полюсов — вегетативном. Противоположный полюс, содержащий ядро и цитоплазму без желтка, называется анимальным. В центролецитальных яйцах желток находится в центре клетки, а цитоплазма расположена на периферии (яйца насекомых). Различное строение яиц связано с приспособлением к условиям развития и закрепилось в процессе эволюции.
У животных, которые в постэмбриональный период проходят стадию личинок (иглокожие, насекомые, амфибии), яйца содержат сравнительно немного желтка. Личинки покидают яйцевые оболочки до окончания развития и продолжают его вне яйца. У многих животных с неличнночным типом онтогенеза яйца телолешггальные. У животных с внутриутробным типом развития (млекопитающие) яйца бедны желтком, и он распределен в них равномерно.
В предзнготный период развития в яйце накапливаются рибосомальная и информационная РНК, различные участки цитоплазмы приобретают отличия по химическому составу, образуется ряд структур. Многие из них заметны благодаря присутствию различных пигментов. Под клеточной мембраной образуется кортикальный слой цитоплазмы, содержащий гранулы гликогена. Яйцо приобретает полярность: вегетативный и анимальный полюса.
Эмбриональный период,или эмбриогенез (гр. етЬгуоп — зародыш), начинается с образования зиготы. Окончание этого периода при разных типах онтогенеза связано с различными моментами развития: при личиночном типе — с выходом из яйцевых оболочек, при неличиночном — с выходом из зародышевых оболочек, при внутриутробном — с моментом рождения.
Эмбриональный периодделится на стадии зиготы, дробления, бластулы, образования зародышевых листков, гисто- и органогенеза. Зародыши млекопитающих и человека до образования зачатков органов принято называть эмбрионом, а в дальнейшем плодом.
Зигота, образующаяся в результате слияния женской и мужской гамет, представляет собой одноклеточную стадию развития многоклеточного организма. Участки цитоплазмы яйца, содержащие зерна желтка, митохондрии, пигменты, видны на живых объектах, поэтому в зиготе удалось проследить значительные перемещения цитоплазмы.
В неоплодотворенных яйцах морского ежа в кортикальной, области равномерно расположены зерна красного пигмента. После оплодотворения они перемещаются и образуют красный пояс ниже экватора, в то время как анимальный и вегетативный полюса обесцвечиваются. Таким образом, создаются три зоны цитоплазмы: в ани-малыюй части яйца — непигментированная, ниже экватора — пигментированная, на вегетативном полюсе — бесцветная. В дальнейшем из цито-плазматического материала верхней бесцветной зоны формируется эктодерма, из пигментированной зоны — энтодерма, из нижней — элементы мезодермы.
У ряда видов животных уже в зиготе осуществляется интенсивный синтез белка, матрицей для которого на начальных стадиях развития служит и РНК, синтезированная во время овогенеза, но одновременно синтезируется и новая РНК.
Дробление. Начальный этап развития оплодотворенного яйца (зиготы) носит название дробления. Характер дробления обусловлен типом яйцеклетки. В изолецитальном, бедном желтком оплодотворенном яйце ланцетника, первая борозда дробления в виде щели начинается на анимальном полюсе и постепенно распространяется в продольном меридиональном направлении к вегетативному, разделяя яйцо на две клетки — 2 бластомера. Вторая борозда проходит перпендикулярно первой — образуются 4 бластомера. Третья борозда проходит экваториально: возникает 8 бластомеров. В результате последующих дроблений в меридиональных и экваториальных плоскостях образуется 16, 32, 64 и т. д. бластомеров. Клетки, расположенные на вегетативном полюсе, несколько крупнее, чем на анимальном.
В результате ряда последовательных дроблений формируются группы клеток, тесно прилегающих друг к другу. У некоторых животных такой зародыш напоминает ягоду шелковицы или малины. Он получил название морулы (лат. morum — тутовая ягода).
У млекопитающих желтка в яйцах мало, поэтому дробление полное, но также неравномерное. В различных бластомерах оно идет сразным ритмом, и можно наблюдать стадии 2, 3, 6, 7, 9, 10 и т. д. бластомеров. Одни из них (светлые) располагаются по периферии, другие (темные) находятся в центре. Из светлых клеток образуется окружающий зародыш трофобласт, клетки которого выполняют вспомогательную функцию и непосредственно в формировании тела зародыша не участвуют. Клетки трофобласта обладают способностью растворять ткани, благодаря чему зародыш внедряется в стенку матки. Далее клетки трофобласта отслаиваются от клеток зародыша, образуя полый пузырек. Полость трофобласта заполняется жидкостью, диффундирующей в нее изтканей матки. Зародыш в это время имеет вид узелка, расположенного на внутренней стенке трофобласта. В результате дальнейшего дробления зародыш принимает форму диска, распластанного на внутренней поверхности трофобласта.
В процессе дробления увеличивается число бластомеров, однако бластомеры не вырастают до размеров исходной клетки, а с каждым дроблением становятся мельче Эчо объясняется тем.что митотические циклы дробящейся зиготы не имеют типичной интерфазы пресинтетический период (G1) отсутствует, а синтетический (S) начинается еще в телофазе предшествующего митоза. Во время дробления митозы следуют быстро друг за другом, и к концу периода весь зародыш лишь ненамного крупнее зиготы. В это время бластомеры уже отличаются по характеру цитоплазмы и могут разниться по содержанию желтка и размерам, что накладывает отпечаток на их дальнейшее развитие и дифференцировку.
Дробление яйца заканчивается образованием бластулы. Отметим, что в зиготе и бластомерах ядерно-плазменное соотношение нарушено в пользу цитоплазмы. В клетках бластулы устанавливается типичное для каждого вида животных ядерно-плазменное со: отношение. Начиная с бластулы, клетки зародыша принято называть не бластомерами, а эмбриональными клетками. У ланцетника бластула образуется по достижении зародышем 128 клеток. В силу накопления продуктов жизнедеятельности бластомеров между ними появляется полость (бластоцель, или первичная полость). При полном равномерном дроблении (как у ланцетника) бластула имеет форму пузырька со стенкой в один слой клеток, который назван бластодермой. Стадию бластулы проходят зародыши всех типов животных.
Гаструляция. У всех многоклеточных животных следующим за бластулой этапом развития является гас-труляция, которая представляет собой сложный процесс перемещения эмбрионального материала с образованием двух или трех слоев тела зародыша,называемых зародышевыми листками. В процессе гаструляции следует различать два этапа: а) образование экто-и энтодермы (двуслойный зародыш); б) образование мезодермы (трехслойный зародыш). У животных с изоле-цитальным типом яиц гаструляция идет путем инвагинации, т. е. впячи-вания. Вегетативный полюс бластулы впячивается внутрь наподобие стенки продырявленного резинового мяча. Противоположные полюса бластодермы почти смыкаются, так что бластоцель либо исчезает полностью, либо остается в виде незначительной полости, а из шара возникает двухслойный зародыш.
Внешний слой клеток носит название наружного листка, или эктодермы (гр. есtos—снаружи, derma—кожа), внутренний слой—внутреннего листка, или энтодермы (гр. еntos—внутри). Полость называется гастроцелем, или первичной кишкой, а вход в кишку получил наименование бластопора, или первичного рта. Края его сближаются, образуя верхнюю и нижнюю губы. У пер-вичноротых (к ним относится большинство типов беспозвоночных) бластопор превращается в дефинитивный (окончательный) рот, у вторичноротых (иглокожие и хордовые) из него формируется анальное отверстие либо он зарастает, а рот образуется на противоположном конце тела.
Гаструляция происходит не только путем инвагинации. Другими ее способами являются деляминация (расслоение), эпиболия (обрастание) и иммиграция (проникновение внутрь).
Образование гаструлы путем иммиграции характерно для кишечнополостных. Этот способ заключается в массовом активном перемещении клеток бластодермы в бластоцель. ЭпиболияВстречается у животных, имеющих телолецитальные яйца. При этом способе гаструляции мелкие клетки анимального полюса обрастают и покрывают снаружи крупные, богатые желтком клетки вегетативного полюса, которые становятся внутренним слоем. При делями-нации клетки зародыша делятся параллельно его поверхности, образуя наружный и внутренний зародышевые листки.
Гистогенез и органогенез. Гистогенез — процесс образования тканей, органогенез — формирование органов. Диффгренцированный на три эмбриональных листка зародышевый материал дает начало всем тканям и органам. Из эктодермы развиваются ткани нервной системы, очень рано обособляющиеся. У хордовых она первоначально имеет форму нервной пластин-кч. Эта пластинка растет интенсивнее остальных участков эктодермы и затем прогибается, образуя желобок. Размножение клеток продолжается, края желобка смыкаются, возникает нервная трубка, которая тянется вдоль тела от переднего конца к заднему. На переднем конце нервной трубки путем дальнейшего роста и днфферен-цировки формируется головной мозг. Отростки нервных клеток центральных отделов нервной системы образуют периферические нервы. Кроме того, из эктодермы развиваются наружный покров кожи — эпидермис и его производные (ногти, волосы, сальные и потовые железы, эмаль зубов, вослринимающие клетки органов зрения, слуха, обоняния и т. п.).
Из энтодермы развивается эпителиальная ткань, выстилающая органы дыхательной, частично мочеполовой и пищеварительной систем, в том числе печень и поджелудочную железу.
Миотом дает начало скелетной мускулатуре, нефрогонотом—органам выделения и половым железам (гонадам). Клетки, образующие висцеральные и париетальные листки спланхнотома, являются источником эпителиальной выстилки вторичной погости тела — целома. За счет элементов склеротома развиваются хрящевая, костная и соединительная ткани, образующие вокруг хорды осевой скелет. Дерматом дает начало соединительной ткани кожи, а спланхнотом — соединительной ткани внутренних органов, кровеносным сосудам, гладкой мускулатуре кишок, дыхательных и мочеполовых путей. В образовании сердца принимает участие также висцеральный листок спланхнотома. Железы внутренней секреции имеют различное происхождение: одни из них (эпифиз, часть гипофиза) развиваются из закладок нервной системы, другие— из эктодермы. Надпочечники и половые железы являются производными мезодермы.
Органогенез завершается в основном к концу эмбрионального периода развития. Однако дифференцировка и усложнение органов продолжаются и в постэмбрионалыюм онтогенезе. Описанные процессы связаны не только с активным клеточным размножением первичных эмбриональных закладок, но и с их значительным перемещением, изменением формы тела зародыша, образованием отверстий и полостей, а также с формированием ряда временных зародышевых (провизорных) органов.
В процессе эмбрионального развития у позвоночных формируется ряд зародышевых оболочек, которые играют большую роль в развитии эмбриона. Самая примитивная из зародышевых оболочек — это желточный мешок, который впервые появляется у рыб. Желточный мешок представляет собой мембрану, пронизанную сетью кровеносных сосудов; он расположен снизу под эмбрионом, вокруг желтка, и служит для переноса питательных веществ из желтка к зародышу. Желточный мешок имеет очень большое значение у птиц п у рептилий, яйца которых содержат много желтка.
Несмотря на то, что у млекопитающих желток почти или совсем отсутствует, желточный мешок у них все же развивается и в нем образуются первые кровяные островки, а затем и кровеносные сосуды эмбриона, но на более поздних стадиях зародышевого развития эта оболочка превращается вмаленький рудиментарный орган.
Вторая зародышевая оболочка — амнион — формируется вокруг эмбриона у наземных позвоночных; амнион наполнен амниотической жидкостью, в которую погружен зародыш. Таким образом, эмбрионы наземных позвоночных находятся в водной среде, несмотря на то что взрослые животные обитают на суше. Амниотическая жидкость служит опорой для нежного зародыша и защищает его от повреждений. Третья оболочка — аллантоис — образуется из заднего конца первичной кишки.
У пресмыкающихся и птиц эта оболочка расположена вокруг амниона и желточного мешка и прилегает к скорлупе яйца. Аллантоис у этих животных функционирует в качестве эмбриональных легких, поглощая кислород и выделяя углекислый газ через скорлупу яйца. В птичьем яйце, вскрытом в конце периода насиживания, можно видеть непосредственно под скорлупой эту пронизанную кровеносными сосудами зародышевую оболочку. У млекопитающих аллантоис разрастается и, соприкасаясь со стенкой матки, образует часть плаценты. Кислород и питательные вещества из крови матери проходят через плаценту и по пупочному канатику переносятся эмбриональной кровью в организм зародыша. Продукты жизнедеятельности эмбриона проходят через плаценту и поступают в кровь матери.
Четвертая оболочка — хорион — не играет существенной роли в развитии эмбрионов птиц и рептилий; хорион в яйцах этих животных примыкает непосредственно к скорлупе яйца, но практически не функционирует. У млекопитающих роль хориона, напротив, очень важна: он прорастает слизистую матки и адсорбирует питательные вещества на той, самой ранней стадии формирования зародыша, когда еще не развились ни кровеносные сосуды, ни аллантоис. Хорион также принимает участие в образовании плаценты; кроме того, в нем вырабатывается гормон, играющий большую роль в нормальном развитии эмбриона.
Взаимоотношение материнского организма и плода. У млекопитающих и человека яйцеклетка бедна желтком, поэтому провизорные приспособления развивающегося организма имеют свои особенности. Желточный мешок закладывается на ранних этапах эмбриогенеза, но не развивается, а постепенно редуцируется, расслаивается. Аллантоис также не развит. Зачаток его входит в состав нового специфического провизорного органа — пупочного канатика.
Функцию наружной зародышевой оболочки выполняет хорион, или ворсинчатая оболочка, названная так вследствие развития на ее поверхности большого числа выростов, ворсинок. Ворсинки хориона врастают в слизистую оболочку матки — специального органа материнского организма, присущего только млекопитающим. Место наибольшего разветвления ворсинок хориона и наиболее тесного контакта их со слизистой оболочкой матки носит название детского места, или плаценты.
Связь тела зародыша с плацентой осуществляется через пуповину или пупочный канатик, содержащий кровеносные сосуды. Кровеносные капилляры тела зародыша разветвляются в ворсинках хориона. Так устанавливается плацентарное кровообращение. Кровь матери не смешивается с кровью плода; она омывает ворсинки хориона, но никогда не проникает в капилляры плода. Через плаценту плод снабжается питательными веществами, кислородом и освобождается от продуктов жизнедеятельности. При этом важная роль принадлежит эпителиальным клеткам, образующим хорион и его ворсинки. Вместе с клетками стенок сосудов эпителий хориона образует специфический клеточный барьер; микроорганизмы и ряд веществ из кровотока матери в норме не поступают в кровоток плода. Нарушение плацентарного барьера, как правило, ведет к расстройству нормального развития плода, к патологии беременности. Плацента не является барьером для ряда лекарственных веществ, в том числе наркотиков, производственных и пищевых ядов, чужеродных белков и антител. Изучение биологических особенностей связи организма плода и матери у высших млекопитающих, а ,следовательно, и у человека, имеет большое значение и лежит в основе правильной организации медицинской службы вобласти охраны материнства.
1. Оплодотворение. Ранний эмбриогенез.
Оплодотворении - процесс слияния зрелых мужской (сперматозоид) и женской (яйцеклетка) половых клеток, в результате чего возникает зигота, несущая генетическую информацию как отца, так и матери.
Созревание половых клеток.
Сперматогенез совершается в извитых семенных канальцах мужских гонад. Он завершается в период половой зрелости образованием зрелых сперматозоидов, обладающих способностью к оплодотворению. Полному созреванию предшествует процесс редукционного деления, в результате которого в ядре сперматозоида содержится гаплоидный набор хромосом.
Сперматозоиды бывают двух видов: носители половых Х- и Y-хромосом. При слиянии с яйцеклеткой сперматозоида, являющегося носителем половой Х-хромосомы, из образующейся зиготы развивается эмбрион женского пола, при слиянии сперматозоида, имеющего половую Y-хромосому, возникает зародыш мужского пола (яйцеклетка всегда является носительницей половой Х-Хромосомы).
Зрелый сперматозоид имеет длину до 50—60 мкм и состоит из головки, шейки и хвостовой части. Головка сперматозоида, имеющая овальную форму, содержит ядро, окруженное тонким слоем протоплазмы. Шейка имеет протоплазму, содержащую видоизмененную центросому, которая способствует процессу дробления оплодотворенной яйцеклетки. Хвостик состоит из протоплазмы и выполняет двигательные функции. В результате колебательных движений хвостовой части сперматозоиды способны совершать самостоятельные движения в половом тракте женщины со скоростью 2—3 мм/мин. Способность к движениям сперматозоиды получают после их контакта с секретом семенных пузырьков и предстательной железы - семенная жидкость, или спермы. Семенная жидкость имеет сложный состав и содержит фруктозу, белковые вещества, протеазы, кислую фосфатазу, лимонную кислоту и биологически активные вещества — простагландины. Семенная жидкость обладает выраженной антигенной активностью (антигены обнаружены в самих сперматозоидах и в жидкой части спермы). В процессе оплодотворения происходит нейтрализация антигенов сперматозоидов.
При половом сношении во влагалище женщины изливается в среднем около 3—5 мл спермы, в которой содержится 300—500 млн сперматозоидов, некоторые из которых имеют те или иные морфологические либо функциональные отклонения. Часть сперматозоидов, в том числе и неполноценных, остается во влагалище и подвергается фагоцитозу. Вместе со сперматозоидами во влагалище попадают и другие составные части спермы, при этом особая роль принадлежит простагландинам. Под их влиянием происходит активация сократительной активности матки и маточных труб, что очень важно для нормального транспорта гамет. Из влагалища, имеющего у здоровой женщины кислую среду (кислая среда неблагоприятна для жизнедеятельности сперматозоидов), последние быстро поступают в цервикальную слизь, которая во время полового акта под влиянием сокращений мышц шейки матки выделяется из цервикального канала, и продвигаются слизи по направлению к матке (спустя 1-2 мин после полового акта сперматозоиды обнаруживаются в матке, а через 1,5-2 часа они достигают концевых отделов маточных труб, где и происходит их слияние с яйцеклеткой). Транспорт сперматозоидов в матку, а затем и в маточные трубы в основном обеспечивается сокращениями гладкой мускулатуры этих органов, высокой кинетической активностью сперматозоидов, движением микроворсин реснитчатого эпителия эндоцервикса и током жидкости в просвете маточной трубы.
В верхних отделах полового тракта женщины начинается процесс - капацитация спермы - приобретение ею благодаря сложным изменениям способности к оплодотворению.
Яйцеклетка. Развитие связано с ростом и развитием первичных фолликулов, находящихся в корковом слое яичников. Перед оплодотворением зрелая яйцеклетка содержит ядро, со всех сторон окружена лучистым венцом, отчетливо заметна блестящая оболочка. Уже через несколько минут после разрыва фолликула (овуляции) яйцеклетка попадает в полость маточной трубы. Этому процессу способствует ряд факторов: "захват" ее фимбриями маточной трубы со стороны яичника, в котором произошла овуляция, направление тока фолликулярной жидкости при разрыве фолликула и др. Способность яйцеклетки к оплодотворению в среднем составляет 24 ч.
Оплодотворение. Яйцеклетка, попавшая в ампулярный отдел маточной трубы, быстро окружается большим количеством сперматозоидов. Сперматозоиды, окружающие яйцеклетку, начинают пенетрировать в клетки лучистого венца в области воспринимающего бугорка. Процесс пенетрации обусловлен наличием ряда ферментов, которые содержатся как в головке сперматозоида, так и в трубной жидкости. Сразу же после слияния мембран половых клеток происходит кортикальная реакция яйцеклетки, являющаяся составной частью обеспечения блока полиспермии. После этого хромосомы зиготы вступают в первое митотическое деление, которое наступает через 24 ч после начала оплодотворения. Ядро оплодотворенной яйцеклетки (зиготы) содержит диплоидный набор хромосом (46).
После оплодотворения (через 24 ч) начинается дробление оплодотворенной яйцеклетки. Первоначально дробление имеет синхронный характер. Через 12 ч от начала возникновения 2 бластомеров возникают 4 бластомера и т.д. К 96 ч от момента слияния ядра сперматозоида с ядром яйцеклетки зародыш состоит из 16-32 бластомеров (стадия морулы). На этой стадии оплодотворенное яйцо (зигота) попадает в матку.
Поскольку дробящаяся яйцеклетка не обладает самостоятельной подвижностью, ее транспорт определяется взаимодействием сократительной активности маточной трубы (основной фактор), движениями цилиарного эпителия эндосальпинкса и капиллярным током жидкости в направлении от ампулярного конца маточной трубы к матке.
Транспорт яйцеклетки по маточной трубе находится под воздействием гормонов желтого тела - прогестерона и эстрогеннов. Под влиянием относительно низкого содержания прогестерона и более высокой концентрации в крови эстрогенов повышается тонус ампулярно-перешеечного отдела трубы. В результате яйцеклетка задерживается в ампулярном отделе, где происходит ее оплодотворение и начинается процесс клеточного деления с образованием бластомеров. В дальнейшем происходит постепенное продвижение оплодотворенной яйцеклетки по перешейку маточной трубы к матке. Под влиянием нарастающих концентраций прогестерона желтого тела сократительная функция маточных труб приобретает перистальтический характер, при этом волны сокращений направлены в сторону матки. Происходит расслабление трубно-маточного соединена, и яйцеклетка из маточной трубы попадает в полость матки.
Имплантация плодного яйца. Проделав путь по маточной трубе в течение 4 сут, плодное яйцо на стадии морулы попадает в матку, где превращается в бластоцисту. Стадия бластоцисты характеризуется тем, что бластомеры подвергаются определенным изменениям. Часть бластомеров, более крупных по своим размерам, образует так называемый эмбриобласт, из которого в дальнейшем развивается эмбрион. Другая часть клеток, более мелких и располагающихся по периферии плодного яйца, образует питательную оболочку - трофобласт. В дальнейшем наиболее развитая часть трофобласта превращается в плаценту. В полости матки бластоциста приближается к месту имплантации (нидации). Локализация имплантации имеет свои закономерности и в значительной степени определяется местными особенностями эндометрия. Обычно бластоциста имплантируется в области передней или задней стенки матки.
Затем начинается погружение бластоцисты в эндометрий, который к этому времени превращается в децидуальную оболочку - видоизмененный функциональным слой эндометрия.
Процесс имплантации, который в среднем продолжается около 2 дней, сопровождается выраженными гемодинамическими сдвигами местного характера. Вблизи места имплантации бластоцисты отмечается расширение кровеносных сосудов и образование синусоидов, представляющих собой расширенные капилляры и венулы. Этим процессам принадлежит большая роль в процессах обмена между материнским организмом и зародышем.
Ранний эмбриогенез. После имплантации происходит быстрое развитие как зародыша, так и его оболочек. На трофобласте образуются ворсинки, которые на ранних стадиях онтогенеза равномерно покрывают всю поверхность бластоцисты. Первичные ворсины являются бессосудистыми образованиями. Постепенно наружный слой трофобласта утрачивает клеточные границы, превращаясь в синцитий (симпласт). Внутренний слой трофобласта сохраняет клеточное строение и называется цитотрофобластом.
Одновременно с трофобластом развивается и эмбриобласт. Этот процесс особенно активизируется после имплантации. Клетки эмбриобласта дифференцируются в два узелка: эктобластический и эндобластический. Вскоре в центральных частях этих узелков образуются полости, в результате чего эктобластический узелок превращается в эктобластический пузырек, а энобластический узелок - в эндобластический пузырек. Эктобластический пузырек затем превращается в амниотическую полость, а эндобластический - в желточный мешок. Из клеточных скоплений эктобласта и эндобласта, расположенных между амниотическим и желточным пузырьком, образуется зародышевый щиток (зародыш).
На ранних стадиях развития происходит дифференцировка мезенхимы. С одной стороны клетки мезенхимы оттесняются на периферию - к трофобласту. С другой стороны происходит скопление мезенхимальных клеток вокруг амниотического и желточного пузырьков, а также около зародыша. В результате этого стенки пузырьков становятся двухслойными, а сам зародыш дифференцируется в экто-, эндо- и мезодерму. Эти три зародышевых листка служат исходным материалом для формирования в дальнейшем всех органов и тканей эмбриона и плода.
По мере дальнейшего внутриутробного развития происходит быстрое увеличение размеров амниотического пузырька, который превращается в амниотическую полость. В полости наблюдается быстрое накопление прозрачной (амниотической) жидкости, при этом стенка амниотического пузырька приближается к ворсинчатой оболочке и, наконец, сливается с ней. Зародыш начинает вворачиваться в полость амниона, а желточный пузырек атрофируется.
Одновременно с развитием плодных оболочек из каудального отдела первичной кишки зародыша образуется выпячивание - аллантоис. По аллантоису сосуды зародыша "подрастают" к ворсинчатой оболочке, врастая затем в каждую ворсину. В результате этого бессосудистый хорион васкуляризируется.
После завершения начальных этапов онтогенеза эмбрион окружен амниотической жидкостью и тремя оболочками: децидуальной, ворсинчатой и водной.
Децидуальная оболочка представляет собой трансформированный в связи с беременностью функциональный слой эндометрия. К моменту имплантации эндометрий находится в секреторной фазе. Он состоит из двух слоев: компактного и спонгиозного. Компактный слой, обращенный в полость матки, содержит выводные протоки маточных желез и клетки стромы эндометрия, превратившиеся во время беременности в децидуальные. Спонгиозный слой в основном состоит из желез.
В соответствии с локализацией плодного яйца в децидуальной оболочке можно различить три части:
• выстилающая полость матки;
• покрывающая плодное яйцо со стороны полости матки;
• расположенная между плодным яйцом и стенкой матки. По мере роста зародыша утолщается, в ней развиваются сосуды и она превращается в материнскую часть плаценты. В нее также врастают ворсины хориона, вокруг образуются межворсинчатые пространства, в которые изливается кровь из материнских сосудов и орошает поверхность ворсин.
Децидуальная оболочка является для плода питательным (осуществляется на ранних этапах внутриутробного развития) и защитным (высокая фагоцитарная активноеть) слоем.
Амнион (водная оболочка) обращена к плоду. Она выстилает плаценту и переходит на пуповину, сливаясь в области пупочного кольца с кожей плода. Микроскопически амнион представляет собой тонкую полупрозрачную мембрану. На ранних стадиях развития эпителий амниона представлен крупным полигональными клетками, с 3-го месяца беременности он становится кубическим. Амнион вместе с гладким хорионом принимает активное участие в обмене околоплодных вод, а также в параплацентарном обмене.
Ранний эмбриогенез
После имплантации происходит быстрое развитие как зародыша, так и его оболочек. На трофобласте образуются ворсинки, которые на ранних стадиях онтогенеза равномерно покрывают всю поверхность бластоцисты. Первичные ворсины являются бессосудистыми образованиями. Постепенно наружный слой трофобласта утрачивает клеточные границы, превращаясь в синцитий (симпласт). Внутренний слой трофобласта сохраняет клеточное строение и называется цитотрофобластом. Более подробно дальнейшие процессы дифференциации трофобласта описаны в следующем разделе (см. раздел 3.2).
Одновременно с трофобластом развивается и эмбриобласт. Этот процесс особенно активизируется после имплантации. Клетки эмбриобласта диффе-ренцируются в два узелка: эктобластический и эндобластический. Вскоре в центральных частях этих узелков образуются полости, в результате чего эктобластический узелок превращается в эктобластический пузырек, а эндобластический узелок - в эндобластический пузырек (рис. 3.6). Эктобластический пузырек затем превращается в амниотическую полость, а эндобластический - в желточный мешок. Из клеточных скоплений эктобласта и эндобласта, расположенных между амниотическим и желточным пузырьком, образуется зародышевый щиток (зародыш).
На ранних стадиях развития происходит и дифференцировка мезенхимы. С одной стороны клетки мезенхимы оттесняются на периферию - к трофобласту. С другой стороны происходит скопление мезенхимальных клеток вокруг амниотического и желточного пузырьков, а также около зародыша. В результате этого стенки пузырьков становятся двухслойными, а сам зародыш дифференцируется в экто-, эндои мезодерму. Эти три зародышевых листка служат исходным материалом для формирования в дальнейшем всех органов и тканей эмбриона и плода.
По мере дальнейшего внутриутробного развития происходит быстрое увеличение размеров амниотического пузырька, который превращается в амниотическую полость. В полости наблюдается быстрое накопление прозрачной жидкости (амниотическая жидкость), при этом стенка амниотического пузырька приближается к ворсинчатой оболочке и, наконец, сливается с ней. Зародыш начинает вворачиваться в полость амниона, а желточный пузырек атрофируется.
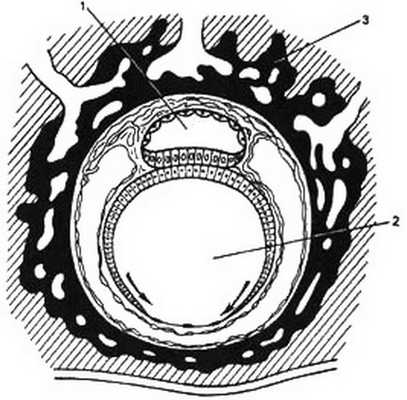
Рис. 3.6. Ранние стадии развития эмбриона. 1 - эктобластический пузырек (амнион); 2 - эндобластический пузырек (желточный мешок); 3 - ворсины трофобласта.
Децидуальная оболочка представляет собой трансформированный в связи с беременностью функциональный слой эндометрия. К моменту имплантации эндометрий находится в секреторной фазе. Он состоит из двух слоев: компактного и спонгиозного. Компактный слой, обращенный в полость матки, содержит выводные протоки маточных желез и клетки стромы эндометрия, превратившиеся во время беременности в децидуальные. Спонгиозный слой в основном состоит из желез.?
В соответствии с локализацией плодного яйца в децидуальной оболочке можно различить три части:
1. выстилающая полость матки (decidua parietalis);
2. покрывающая плодное яйцо со стороны полости матки (decidua capsularis);
3. расположенная между плодным яйцом и стенкой матки (decidua basalis). Указанные взаимоотношения различных частей децидуальной оболочки представлены на рис. 3.7.
В процессе роста плодного яйца decidua parietalis и decidua capsularis растягиваются, истончаются и приближаются друг к другу. На 4-5-м месяце беременности плодное яйцо занимает уже всю полость матки, и decidua parietalis сливается с decidua capsularis. Наоборот, decidua basalis значительно гипертрофируется и превращается в материнскую часть плаценты. В этой части плаценты находятся многочисленные ворсины хориона, вокруг которых формируется межворсинчатое пространство.
Децидуальная оболочка является для плода питательным и защитным слоем: трофическая функция ее в основном осуществляется на ранних этапах внутриутробного развития, защитная роль наиболее полно проявляется высокой фагоцитарной активностью. Децидуальная ткань лизирует микроорганизмы и инактивирует их токсины, принимает также участие в синтезе углеводов, липидов и белков. В ней происходит синтез пролактина и простагландинов. Таким образом, децидуальной оболочке принадлежит очень важная роль в имплантации и дальнейшем развитии эмбриона и плода.
Амнион (водная оболочка) обращена к плоду. Она выстилает плаценту и переходит на пуповину, сливаясь в области пупочного кольца с кожей плода. Макроскопически амнион представляет собой тонкую полупрозрачную мембрану. В процессе эмбриогенеза амнион развивается из эктобластического пузырька. Из эктодермы формируется эпителий амниона, из мезодермы - соединительнотканная основа.
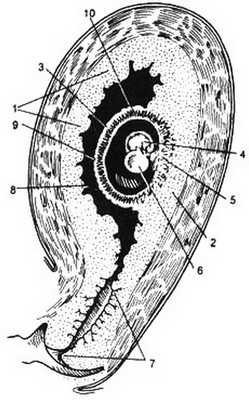
Рис. 3.7. Беременность малого срока. 1 - decidua parietalis; 2 - decidua basalis; 3, 9 - decidua capsularis; 4 - эмбрион в амниотическом мешке; 5 - chorion frondosum; 6 - желточный мешок; 7 - цервикальный канал; 8 - целомическая полость; 10 - ворсины хориона.
На ранних стадиях развития эпителий амниона представлен крупными полигональными клетками, с 3-го месяца беременности он становится кубическим. Эпителиальные клетки амниона содержат липиды, полисахариды, протеины, фосфорные соединения, а также ряд ферментов, участвующих в процессах метаболизма и обмене стероидных гормонов.
Амнион вместе с гладким хорионом принимает активное участие в обмене околоплодных вод, а также в параплацентарном обмене. По своим физическим свойствам плодные оболочки отличаются друг от друга. Так как амниотическая оболочка очень плотная и выдерживает давление в несколько раз большее, чем гладкий хорион, в родах разрыв гладкого хориона наступает раньше, чем амниона.
Научная электронная библиотека

Плодные оболочки - временные образования, окружающие плод в матке. Аллантоис - является производным энтодермы и висцерального листка спланхнотома. В стенке спланхнотома развиваются кровеносные сосуды, в него открывается урахус, по которому в аллантоис поступает фетальная моча. Хорион возник из энтодермы и париетального листка спланхнотома, покрывает трофобласт, который участвует в образовании плаценты. Амнион - произошёл из эктодермы и париетального листкаспланхнотома. По этой причине изучение формы, величины и топографииполостей амниона и аллантоиса имеет биологическое и практическое значение для козоводства.
Возникают они в период обособления зачатков тканей и органов, что сопровождается повышенной потребностью зародыша, предплода, плода в белках, липидах, углеводах и минеральных веществах. С ростом и развитием зародыша, предплода, плода этих веществ в маточном молоке не хватает, для защиты и удовлетворения этих потребностей формируются плодные оболочки и плацента.
Общепризнанно, что зародыш, предплод и плод на протяжении всего внутриутробного развития находится в полости амниона, а аллантоис с жидкостью расположен вентрально и с боков плода, в виде двух слепых мешков, повторяющих форму полостей рогов матки.
На 25 сутки беременности коз аллантоис по объему превосходит амнион, он имеет вытянутую форму, заполнен жидкостью и занимает всю полость плодовместилища матки. Из тела матки он переходит в свободный рог, где фиксируется связкой в его верхушке. Связка тонкая, нитевидная и имеет желтоватый оттенок (М.С. Сеитов,2001). Таким образом, амнион снаружи покрыт аллантоисом, жидкость, содержащаяся в нём, прозрачная, слегка студнеобразной консистенции, защищает зародыш от механических воздействий. Околоплодные оболочки тонкие, прозрачные, через них просматривается зародыш, имеющий подковообразную форму, масса оболочки равна 1,0 ± 0,09 г, длина аллантоиса 115,4 ± 2,1 м, а амниона - 8,0 ± 0,44 мм. Зародыш занимает почти весь амниотический пузырь (рис. 7).
На 30 сутки связки аллантоисного пузыря уже хорошо выражены, но рыхлые и при натягивании выходят из маточного отверстия яйцеводов в виде тонких шнуровидных образований желтого цвета.

Рис. 7. Беременность 25 суток. Эмбрион защищен не только амниотической, но и аллантоисной оболочками
Масса околоплодных оболочек, без их содержимого, увеличивается в 7,5 раз, а длина - в 2,6 раза по сравнению 25-суточной беременностью. Отмечается увеличение объемов как аллантоисной, так и амниотической жидкости. Однако линейные размеры амниотического пузыря, по сравнению с предыдущим этапом, возрастают несколько интенсивнее, так длина в 3,1, а ширина - в 3,5 раза. Зародыш занимает большую часть полости амниона.
На 35 сутки беременности амниотический пузырь увеличивается в объеме и приближается к стенке аллантоиса. В этом месте аллантоис слепо, инвагинирует, объем его несколько уменьшается при общем увеличении аллантоисной жидкости. И всё же аллантоис с жидкостью прикрывает предплод со стороны спины. Через оболочки пузырей хорошо просматривается предплод, занимающий большую часть объема амниона. Масса оболочек по сравнению с предыдущим этапом увеличивается в 1,3, длина аллантоиса - в 1,3, амниона - в 1,4 и ширина его - в 1,8 раза, то есть просматривается опережающий рост амниона (рис. 8).
На 45 сутки беременности коз стенки аллантоиса смыкаются над предплодом, а в области первичной инвагинации амнион имеет шаровидно-овальную форму. Его стенка тонкая, прозрачная, по этому хорошо виден предпдод. Покоится он на короткой пуповине в небольшом количестве амниотической жидкости. Масса оболочек по сравнению с предыдущим этапом увеличивается в 4,3, длина аллантоиса - в 1,4, амниона - в 1,6 и ширина его - в 1,5 раза
(рис. 9).

Рис. 8. Беременность 35 суток. Аллантоисная оболочка спадается в области расположения амниона

Рис. 9. Беременность 45 суток. Обособление амниотической оболочки
С 60 дней беременности происходит ускорение роста плода, объема амниотической жидкости, аллантоис занимает превалирующее положение в плодном пузыре, а аллантоисный пузырь лежит с боковых сторон от амниона, причём подавляющая часть аллантоиса с жидкостью выдавливается в свободный рог матки. Такое взаимоотношение плодных оболочек уже остается до конца плодоношения. Масса плодных оболочек по сравнению с 45-дневной беременностью увеличивается в 2, длина аллантоиса в 1,1 раза, а амниотического пузыря - в 3,1, а ширина - в 2,3 раза. При этом длина пуповины увеличивается, что позволяет плоду свободно перемещаться в жидкости амниона (рис. 10).

Рис. 10. Беременность два месяца. Плод в дорсальной части защищен только амниотической оболочкой
В 90 дней беременности масса плодных оболочек, по сравнению с предыдущим этапом, увеличивается в 1,6, длина аллантоиса - в 1,3, амниона - в 3,0 раза, ширина последнего - в 1,5 раза, в 120 дней, соответственно, в 2,0; 1,1; 1,4 и 1,1 раза.
В 150 дней беременности масса плодных оболочек возрастает в 1,6 раза, длина аллантоиса - в 1,07, длина амниона - 1,1 и шири-
на - в 1,06 раза (рис. 11).

Рис. 11. Беременность три месяца. Плод защищен значительным слоем околоплодной жидкости
Из краткого анализа следует, что аллантоисный пузырь к концу беременности перестает выполнять роль дополнительного жидкостного буфера для плода, по этой причине он располагается в области средних боковых поверхностей плода и в свободном роге, что оправдано с биологической точки зрения. В 1,5-2,0 месяца беременности зародыш, предплод и ранний плод чрезвычайно чувствительны к механическим воздействиям, ткани студнеобразной консистенции и органы очень ранимы. В 100 % случаев мы наблюдали в эти этапы петехии и энхимозы. Они чаще располагались в области головы, крестца, спины, реже на боковых поверхностях, конечностях и носили прижизненный характер.
Читайте также:
- Характеристика подпольно произведенного амфетамина (наркотика)
- Профилирование экспрессии генов для прогноза рака молочной железы
- Атипичная пневмония
- Профилактика постренальной формы острой почечной недостаточности. Принципы профилактики постренальной формы опн.
- Калий и кислотно-основное равновесие крови. Гиперкалиемия