Тромбоциты. Структура тромбоцитов. Функции тромбоцитов. Функции гликопротеинов. Зона золя - геля гиалоплазмы.
Добавил пользователь Morpheus Обновлено: 09.01.2026
Система гемостаза очень сложна, при этом сбалансирована так чтобы обеспечивать функциональное состояние крови, протекающей в сосудах человека, а также для того чтобы обеспечить свертывание крови в местах повреждения тканей для минимизации кровопотери и скорейшего восстановления поврежденной ткани. Система гемостаза включает в себя:
Факторы коагуляции - белки в основном вырабатываемые в печени, участвующие в каскаде коагуляции;
Тромбоциты - клеточные элементы крови, концентрирующиеся и формирующие процесс аггрегации в области повреждения и образующие первичный тромб;
Фибринолитические факторы - ферменты, участвующие в лизисе сгустка крови после процессов заживления и регенерации поврежденных тканей;
Ингибиторы - факторы коагуляции, которые помогают локализовать сгусток в месте повреждния и препятствуют развитию генерализации процесса тромбообразования в сосудах всего организма;
Клетки эндотелия - это выстилающие внутреннюю поверхность сосуда, эпителиальные клетки. В неповрежденном состоянии сосудов и тканей они обеспечивают нормальный ток крови, в случае травматизации последних, они генерируют выработку биологически активных веществ: факторов коагуляции и фибринолиза.
Тромбоциты - клетки, крови, имеющие форму диска, содержащиеся в количестве 150 - 450 тыс.1/мл3 крови. Они являются основным компонентом в образовании первичного тромба и являются важнейшим фактором нормального гемостаза. Количественные и качественные нарушения тромбоцитарного состава приводят к нарушению нормального гемостаза и склонности к кровотечениям соответственно. В данной монографии рассматривается структура, физиология и основные нарушения функций тромбоцитов. Мы не будем подробно останавливаться на тех нарушениях, связанных с уменьшением количественного состава тромбоцитов (тромбоцитопения) и нарушениях функций тромбоцитов, возникших в связи с применением некоторых лекарственных препаратов или при некоторых болезненных состояниях организма.
Клеточная структура тромбоцита
Тромбоциты трансформируются из мегакариоцитов костного мозга в процессе цитогенеза. Тромбоциты не имеют клеточного ядра. При рассматривании под микроскопом мазка крови, окрашенного по методу Райта-Гемза тромбоциты представляют собой маленькие гранулированные клетки с грубой мембраной, в количестве от 3 до 10 штук в визуальном поле. Из общего количества тромбоцитов находящихся в единицу времени в организме - 70% циркулируют в общем объеме крови, а 30% находятся в депонированном «спящем» состоянии. Жизненный цикл тромбоцитов составляет около 10 дней, затем они утилизируются ретикулоэндотелиальными клетками печени и селезенки. Средний нормальный объем тромбоцитов составляет примерно 7 + / - 4,8 унций. Средний нормальный объем является триггером в процессе тромбопоэза, а поскольку молодые тромбоциты имеют больший средний нормальный объем, но это может быть и связанно с некоторыми специфическими нарушениями функции тромбоцитов.
Метод визуального наблюдения структуры тромбоцитов под микроскопом не может в полной мере отразить всю ее сложность. Внутренняя структура тромбоцита состоит из 4 зон: периферийная зона, золево-гелевая зона, органеллы и клеточная мембрана.
Периферийная зона состоит из внешних мембран и ассоциированных с ними структур. У тромбоцитов имеется соединяющая с внешней средой система каналов, названная открытая канальцевая система (ОКС). Стенки канальцев открываются в эту зону. Через ОКС в тромбоцит поступают элементы плазмы крови и выделяются продукты генерируемые собственно тромбоцитом. Выделение тромбоцитарных продуктов через ОКС в результате активизации тромбоцита называется «реакцией выделения или высвобождения». Мембрана тромбоцита богата гликопротеинами, которые являются рецепторами для компонентов этой триггерной системы активации тромбоцита и субстратами для адгезии (склеивания) и аггрегации (сцепления) тромбоцитов. К периферийной зоне также находятся фосфолипиды мембраны. Фосфолипиды являются важным компонентом в процессе коагуляции потому, что являются матрицей для коагуляции протеинов. Фосфолипиды также являются инициирующим субстратом тромбоцитарных ферментов, продуцирующих тромбоксан А2, являющимся выжным продуктом в процессе активации тромбоцитов и мощным томбоцитарным кофактором (вызывающим аггрегацию тромбоцитов). Тромбоцитарная мембрана также переводжит сигналы с поверхности клетки во внутренние химические сигналы. Золево-гелевая зона расположена внутри периферийной зоны и участвует в формировании клеточного скелета тромбоцита. Клеточный скелет в свою очередь обуславливает дискообразную форму тромбоцита, которая после активации позволяет расширяться псевдоподиям и высвобождать содержимое грануляций. Они составляют от 30 до 50% общего тромбоцитарного белка.
Зона органелл состоит из зерен и клеточных компонентов: лизосом, митохондрий и т. д. Эти органеллы участвуют в процессе обмена веществ клетки, хранения ферментов и других необходимых для функционирования клетки веществ. В ней содержатся два отдела адениновых нуклеотидов: хранилище или резервуар твердых зерен, и метаболический или цитоплазматический резервуар. В данной зоне содержатся альфа- и твердые зерна.
Твердые зерна содержат неметаболический аденозинтрифосфат (АТФ) и аденозиндифосфат (АДФ), серотонин и кальций. Альфа-зерна содержат адгезивные белки: фибриноген, фактор Виллебранда, тромбоспондин, и витронектин. Альфа-зерна также содержат вещества содействующие росту клетки: клеточный гормон роста, тромботический фактор IV и преобразующий гормон роста клетки. Факторы коагуляции, в том числе, кининоген высокой молекулярной массы, фактор XI и активатор плазминогена ингибитор-1 также присутствуют в виде альфа-зерен.
Четвертая зона - система мембран, включающая твердую систему, состоящую из трубочек. Здесь концентрируется кальций, необходимый для контрактильных процессов трансформации тромбоцита. Там также находится гормональная система для синтеза простагландинов.
Роль тромбоцитов в процессе гемостаза
Первичный гемостаз
Процесс трансформации активированных тромбоцитов в устойчивый тромб является единым процессом, который можно разделить на четыре этапа: адгезию, аггрегацию, уплотнение и секреция.
В инертном виде тромбоциты не слипаются со стенкой сосуда. При повреждении сосудистой стенки, происходит выброс субэндотелиальных продуктов, таких как коллаген. Тромбоциты адгезируются с коллагеном и формируют гемостатически эффективный тромб. Это взаимодействие может быть инициировано фактором Виллебранда, имеющим тромбоцитарный рецептор Ib-IX. Привлечение большего числа происходит при тромбоцито-тромбоцитарном контакте, который инициируется, в основном, через фибриногеновый рецептор, GPIIb-IIIa. Адгезивный процесс по схемам тромбоцит-тромбоцит и тромбоцит-коллаген может происходить без сжатия и изменения формы клетки.
Сжатие тромбоцитов во время и после активации обуславливает сложность их формы и отрицательный заряд компонентов мембраны. Оно приводит к секреции содержимого гранул, превращая неплотно соединенное скопление тромбоцитов в однородную массу. Секреция содержимого гранул вызывается сигналом, прошедшим сквозь мембрану тромбоцита, с последующим притоком и высвобождением кальция. Это происходит без перестройки химических субстанций, содержащихся в тромбоците, что может свидетельствовать о повреждении тромбоцита. Содержимое гранул имеет высокую концентрацию внутри тромба и находится близко к поверхности первичного тромба, что обуславливает нормальный процесс коагуляции на поверхности тромбоцита. Это является основой для начала формирования фибринового сгустка - фазы вторичного гемостаза.
Внутренние биохимические процессы
Основным элементом, ответственным за дальнейшее привлечение тромбоцитов в первичный тромб являются: АДФ и тромбоксан А2. Тромбоксан А2 формируется в результате внутриклеточного метаболизма из арахидоната с трансформацией последнего в вещества под названием экосаноиды. Простимулированные тромбоциты высвобождают основные соли жирной арахидоновой кислоты, которые окисляются циклооксигеназой в эндопероксиды. Эндопероксиды затем подвергаются воздействию тромбоксанового фермента и в результате образуется тромбоксан А2. Тромбоксан А2 в жидкой состоянии является потенциальным инициатором аггрегации тромбоцитов. Он также индуцирует гидролиз фосфоинозитидов, фосфориляцию белков и увеличение свободнорадикального внутриклеточного кальция. Прием такого препарата, как аспирин приводит к необратимому ацетилированию циклооксигеназы в результате чего эндопероксиды не могут формироваться (См. рисунок 2).
Резюме
Вклад тромбоцитов в процесс гемостаза состоит в образовании первичного тромба, секреции веществ необходимых для дальнейшего привлечения тромбоцитов, предоставления подходящего поля для дальнейшего процесса коагуляции, выработке промоторов заживления эндотелия и структуры сосуда. Нарушение любого из вышеназванных процессов вызывает нарушения функции тромбоцитов, которые могут быть врожденными или приобретенными.
Условные сокращения: ЦО - циклооксигеназа; DAG - диацилглицерин; IP3 - инозиттрифосфат; ЛМЦ - легкая миозиновая цепь; КЛМЦ - киназа легкой цепи миозина; PIP2 - бифосфат фосфатдилинозит; КПС - киназа протеина С; ФЛС - фосфолипаза С; ФЛА2 - фосфолипаза А2; фВ - фактор Виллебранда; БВ - болезнь Виллебранда.
Оценка функции тромбоцитов
В начале следует попытаться оценить полноценность и завершенность первичного гемостаза путем ретроспективного анализа истории болезни и внешнего осмотра пациента. Симптомы характерные для нарушений первичного гемостаза присутствуют у большинства пациентов с нарушениями функций тромбоцитов и выражаются в кровотечениях из слизистых оболочек и подкожных кровоизлияниях. Однако симптоматика среди пациентов с различными нарушениями функций тромбоцитов различна. Кровотечений из слизистых оболочек и подкожные кровоизлияния выражаются петехиями, подкожныеми геморрагиями и экхимозами, носовыми кровотечениями, меноррагиями, кровотечениями из ЖКТ и посттравматическими кровоизлияниями.
Следует проводить подробный сбор анамнеза. Например, при носовых кровотечениях следует уточнять их частоту и продолжительность, способ остановки, наличие анемии, необходимость переливания крови, проводилось ли процедура прижигания, тампонада. Все это может помочь в оценке истинной тяжести заболевания. Особенности в сборе анамнеза существуют также при подкожных кровоизлияниях и меноррагиях. При геморрагиях следует уточнять число прокладок или тампонов, используемых в день, длительность менструального кровотечения, наличие анемии, необходимость переливания крови, гормональная терапия, гистерэктомия в попытке остановить кровотечение. Обзор всех медикаментов, в том числе прописанных, продаваемых без рецепта и растительных лекарственных средств; когда пациент начал прием этих препаратов, их влияние на течение болезненного состояния. После составления личной истории пациента, следует провести сбор семейного анамнеза. Присутствие или отсутствие сходных симптомов среди членов семьи может помочь в установке правильного диагноза и выявлению подверженных этому заболеванию.
Комплексный осмотр следует проводить, обращая особое внимание на характерные физические признаки, заключающиеся не только в кровотечениях, связанных с нарушениями функции тромбоцитов. Эти нарушения включают, но не ограничены, это: альбинизм, дефекты соединительной ткани (тонкая кожа с ненормальным рубцеванием, как при синдроме Элерса-Данло; удлиненные конечности, хрупкие суставы, и другие особенности, связанные с синдромом Марфана) Другие физические нарушения, связанные, с приобретенными нарушениями функций тромбоцитов: спленомегалия, заболевания сердца, заболевания печени и почек также следует принимать во внимание при осмотре пациента.
Сложность проводимых анализов зависит от уровня технической обеспеченности лаборатории. Минимально необходимыми являются: анализ, определяющий количество тромбоцитов и клинический анализ крови (гемоблогин, тромбоциты, лейкоциты). Результаты этих простых анализов могут обеспечить врача минимально необходимым объемом информации, в частности числом тромбоцитов. Постановка диагноза при многих нарушениях функции тромбоцитов облегчается исследованием образца периферической крови. Этот шаг является обязательным при. Присутствие шистоцитов и может свидетельствовать о микроангиопатических процессах, происходящих в организме, например гемолитическом уремическом синдроме или о тромбоцитопенической пурпуре. Обнаружение тромбоцитов увеличенных в размере, но в недостаточном количестве, при нормальной морфологии красных и белых кровяных клеток, может говорить о иммунно связаных процессах. Увеличенный размер включенных в тромбоцит гранул, отмечается при синдроме Чедиака-Хигаси. Тромбоциты увеличены в размерах при синдромах Бернара-Сулье и Мэя-Хеггелина. При синдроме Скотта-Олдрича (экзема при осмотре) тромбоциты уменьшены в размере и их число снижено. Тромбоциты приобретают сероватый оттенок при синдроме серых тромбоцитов. Однако, при большинстве нарушений функций тромбоцитов, в том числе дефектах хранения и реакции выпуска, включая тромбостению Гланцмана, число тромбоцитов и их морфология - в пределах нормы. Таким образом, в этих случаях требуется проведение дальнейших исследований.
Дополнительные лабораторные исследования, в том числе: время кровотечения и аггрегация тромбоцитов, применяются при определении функции тромбоцитов.
Будучи технически чувствительным, при правильном выполнении, анализ времени кровотечения позволят производить лишь in vivo оценку функции тромбоцитов. Определение времени кровотечения, зависти от лаборанта проводящего данное исследование и должно выполняться опытным специалистом. Нормальное время кровотечения, варьируется в зависимости от метода измерения и возраста пациента, но как правило не превышает 9 минут. Удлинение времени кровотечения при нормальном показатели числа тромбоцитов, указывает на нарушение функции тромбоцитов.
Исследования функции тромбоцитов проводятся в специально оборудованных лабораториях, специально обученным персоналом. Данные исследования предоставляют информацию необходимую для классификации заболевания. Специфические исследования функции тромбоцитов, производятся на образцах плазмы богатой тромбоцитами или цельной крови при участии особых реактивов. Полученные результаты дают возможность врачу поставить точный диагноз. Редкие реактивы, например ристоцетин в малых дозах и криопреципитат, требуются для уточнения типа болезни Виллебранда от псевдоболезни Виллебранда (дефекта тромбоцитарного рецептора Ib). Наиболее распространенными реактивами являются: АДФ, адреналин, коллаген, арахидоновая кислота, ристоцетин и тромбин. Забор крови и ее доставка в лабораторию для данного исследования должен проводиться в строгом соответствии с инструкцией, во избежание преждевременной активации тромбоцитов. Температура,
При болезни Виллебранда и синдроме Бернара-Сулье отмечаются нарушения в аггрегации с ристоцетином. При тромбастении Гланцмана аггрегационаная кривая представляет собой прямую линию со всеми реагентами, за исключением ристоцетина. Дефекты высвобождения и хранения вызывают нарушения аггрегации с АДФ, адреналином, коллагеном и арахидоновой кислотой. Существую также более специфические методы исследования в том числе структуры тромбоцита с помощью электронного микроскопа, структуры гранул, наличие и число тромбоцитарных рецепторов (Iib-IIIa или Ib), а также исследование внутриклеточных гормонов.
Тромбоциты. Структура тромбоцитов. Функции тромбоцитов. Функции гликопротеинов. Зона золя - геля гиалоплазмы.
Тромбоциты являются безъядерными образованиями. В крови основная масса тромбоцитов имеет характерную дискоидную фор¬му. Дискоидная форма поддерживается у тромбоцита циркулярным микротубулярным кольцом. У тромбоцита имеется 4 основные функциональ¬ные зоны. Первая - периферическая зона, представляет собой двухслойную фосфолипидную мембрану и пространства, прилегающие к ней с двух сторон. Золь-гель зона, является вязким матриксом цитоп¬лазмы тромбоцита, прилегая к субмембранной области. Зона органелл, включает в себя органеллы, рас¬положенные по всей цитоплазме неактивных тромбоцитов. Зона мембран, состоит из каналов плотной тубулярной системы, напоминающей структуру миоцитарного саркоплазматического ретикулума. Тромбоциты являются основой всего первичного гемостаза за счет их способности путем агрегации тромбировать повреждения в сосудах. Формируя динамичные агрегаты, тромбоциты влияют на реологические свойства крови и тем самым на состояние трофики тканей во всем организме.
1. Громнацкий Н.И., Медведев И.Н. Тромбоцитарный гемостаз у больных артериальной гипертонией с метаболическим синдромом // Международный медицинский журнал. Клиника. Диагностика. Лечение. - 2002. - № 5. - С. 413.
2. Громнацкий Н.И., Медведев И.Н. Коррекция нарушений тромбоцитарного гемостаза немедикаментозными средствами у больных артериальной гипертонией с метаболическим синдромом // Клиническая медицина. - 2003. - Т.81, № 4. - С.31-34.
3. Громнацкий Н.И., Медведев И.Н., Кондратов И.В. Изменения внутрисосудистой активности тромбоцитов больных артериальной гипертонией с метаболическим синдромом и его коррекция ловастатином (медостатиномR) // Русский медицинский журнал. - 2003. - № 5. - С.258.
4. Краснова Е.Г., Медведев И.Н. Тромбоцитарная активность гемостаза у поросят молочного питания // Ветеринарная практика. - 2011. - № 3. - С.34.
6. Медведев И.Н., Громнацкий Н.И., Наумов М.М., Беспарточный Б.Д. Способ лечения тромбоцитовпатии при метаболическом синдроме. Патент на изобретение RUS 2261705 29.01.2004.
7. Медведев И.Н., Беспарточный Б.Д., Горяинова И.А. Способ профилактики тромбоцитарных нарушений у новорожденных телят с диспепсией. Патент на изобретение RUS 2323731 27.11.2006.
8. Медведев И.Н., Беспарточный Б.Д. Способ коррекции тромбоцитопатии у лиц с метаболическим синдромом. Патент на изобретение RUS 2333758 26.02.2007.
9. Медведев И.Н., Краснова Е.Г., Завалишина С.Ю. Тромбоцитарная активность у поросят в фазу молозивного и молочного питания // Ветеринарная практика. - 2011. - № 4. - С.30.
10. Симоненко В.Б., Медведев И.Н., Кумова Т.А. Воздействие эпросаратана на агрегационную способность тромбоцитов у больных артериальной гипертонией при метаболическом синдроме // Клиническая медицина. - 2008. - Т. 86, № 4. - С. 19-21.
Тромбоциты являются безъядерными форменными элементами крови, участвующими в процессе гемостаза [4]. В крови основная масса тромбоцитов имеет характерную дискоидную форму с почти гладкой поверхностью. Диаметр их составляет от 2 до 4 мкм, площадь поверхности около 8 мкм 2 , а объем - 6-9 фл (фл - фемтолитр=10 -15 л). Дискоидная форма поддерживается у тромбоцита циркулярным микротубулярным кольцом, локализующимся у внутренней поверхности мембраны. Тромбоциты обладают двухслойной мембраной, которая по своему составу и строению несколько отличается от мембран прочих форменных элементов крови тем, что в ней больше фосфолипидов расположено асимметрично [6].
В кровяных пластинках непосредственно у внутреннего слоя мембраны находится микротубулярное кольцо, образованное белком тубулином, локализованное вдоль максимальной окружности мембраны. Тубулин занимает относительно большую поверхность, вследствие чего сохраняется дискоидная форма интактных кровяных пластинок. У дискоидных форм микротрубочки локализуются по внутреннему периметру мембраны - в случае активации они разрушаются и хаотично распределяются по цитоплазме с последующим изменением формы клетки из дискоидной в сферическую.
Тромбоциты в результате стимуляции проявляют выраженную адгезию, агрегацию и секрецию. Так, в случае соприкосновения с чужеродной поверхностью тромбоцит активируется, превращается в сфероцит, имеющий множество отростков, размер которых может существенно превышать поперечник самих тромбоцитов. В основе данных изменений формы лежит нарастание уровня Са 2+ в их цитоплазме, что ведет к деполимеризации тубулина, приводя к растворению микротубулярного кольца и ультраструктурной перестройке внутренней части тромбоцитов с формированием нитей актина. Возникновение псевдоподий обеспечивает быстрый контакт отдельных тромбоцитов между собой, замедляя кровоток в месте их активации [10].
У тромбоцита имеется 4 основные функциональных зоны. Первая - периферическая зона, представляет собой двухслойную фосфолипидную мембрану и пространства, прилегающие к ней с двух сторон. Мембранные интегральные белки проникают сквозь мембрану и обеспечивают связь с цитоскелетом кровяной пластинки. Кроме того, они выполняют функции рецепторов, каналов, насосов, участвуя в процессе активации тромбоцита. Часть интегральных протеинов, имеющих массу полисахаридных молекул, на поверхности тромбоцитов образуют внешнее покрытие липидногобислоя - гликокалекс, способный адсорбировать на себе большое количество белков. В этой связи периферическая зона кровяных пластинок осуществляет барьерную функцию, способствуя обеспечению нормальной формы тромбоцита, реализуя сквозь неё обмен веществ, активацию и весь процесс участия тромбоцитов в гемостазе [7].
Вторая зона - золь-гель зона, является вязким матриксом цитоплазмы тромбоцита, прилегая к субмембранной области. В нее входят различные белки (до 50 % от всех белков тромбоцитов). Их состояние зависит от интактного или активного состояния самого тромбоцита. Внутри зоны золь-гель находится большое число зёрен гликогена, по сути, являющихся запасом энергетического субстраттромбоцита. Также в этой зоне локализованы сократительные протеины, в связи с чем она весьма важна для ретракции агрегировавших тромбоцитов и для их реакции высвобождения [5].
Третья зона - зона органелл, включает в себя органеллы, расположенные по всей цитоплазме неактивных тромбоцитов: пероксисомы, митохондрии, 3 типа гранул хранения (α -гранулы, плотные гранулы и γ-гранулы (лизосомы)) и аппарат Гольджи.
Наиболее многочисленными являются α-гранулы. В одной клетке их может быть от 40 до 80. В их состав входит более 30 белков, необходимых для гемостаза. α-гранулы - это основной источник прокоагулянтных веществ тромбоцитов, они могут экспрессировать на своей наружной поверхности фосфолипиды с отрицательным зарядом, фактор V, GpIIb/IIIа, CD63. Кроме того, они необходимы для образования микровезикул, обладающих прокоагулянтной активностью. В α-гранулах находятся вещества, имеющие прямо противоположные свойства (ингибиторы и активаторы фибринолиза; соединения, стимулирующие и тормозящие процесс ангиогенеза), входящие в состав различных субпопуляциях α-гранул.
Как и в α-гранулах, в плотных безбелковых гранулах, находятся вещества, необходимые для процесса тромбоцитарного гемостаза - серотонин, адениновые нуклеотиды, Са 2+ , фибриноген, адреналин, фактор Виллебранда, антигепариновый фактор. В лизосомах скрыты различные гидролитические ферменты [3,5].
Четверная - зона мембран, состоит из каналов плотной тубулярной системы (ПТС), весьма напоминающей структуру миоцитарного саркоплазматического ретикулума. В ней хранится и из нее идет секреция Са 2+ , что имеет большое значение для активации тромбоцита [6].
Большие концентрации АДФ (из поврежденных эритроцитов и стенок сосудов), а также оголенные субэндотелиальные структуры быстро активируют тромбоциты, меняя их форму, способствуя возникновению выростов и отростков (псевдоподий) с выделением гранул (дегрануляция) в окружающую среду.
Процесс активации тромбоцитов сопровождается выделением ионов Са 2+ из внутриклеточных гранул, что связано с опосредуемым фосфолипазой С гидролизом фосфатидилинозитол-4,5-бисфосфата до 1,4,5-инозитолтрифосфата (ИТФ) и диацилглицерола. После этого ИТФ соединяется со специфическим рецептором, что приводит к поступлению Са 2+ внутрь кровяных пластинок, сопрягаясь с прохождением его сквозь плазматическую мембрану. Это носит название «гранулоуправляемый» кальциевый вход. Очень важна в этом процессе Са 2+ - воспринимающая молекула внутриклеточных гранул - молекула стромы взаимодействия-1 - STIM1 (stromalinteractionmolecule 1) и четыре трансмембранных белковых канала CRACM1 (Calcium-releaseactivatedcalciummodulator) или Orai1. Под воздействием STIM1 в плазмолемме происходит открытие Orai1 канала. Еще один механизм, ведущий к поступлению ионов Са 2+ в тромбоциты, непосредственно связан с рецептор-зависимым кальциевым каналом - Р2Х1, активирующим под действием диацилглицерола, возникающим в ходе распада фосфатидилинозитол-4,5-бисфосфата. При этом поступление кальция в тромбоциты сквозь плазматическую мембрану возможно благодаря TRPC (canonical transient receptor potential channel) в ходе работы натриево-кальциевого насоса [8].
Очень важным для инициации адгезии является столкновение тромбоцитов между собой и стенкой сосудов. Вместе с тем наиболее важную роль в продвижении кровяных пластинок к очагу адгезии играют сдвиговые силы, развивающиеся в ходе циркуляции крови. Под влиянием индуктора происходит активация тромбоцитов, наступающая в следующей последовательности: изменение формы кровяных пластинок, агрегация, генерация эндопероксидов, простагландинов и тромбоксана, выброс в плазму плотных гранул и α-гранул.
Выраженность агрегации в значительной степени зависит от содержания в крови молекул фибриногена и числа мономеров фибрина. Для образования устойчивой связи между двумя тромбоцитами достаточно между ними одной нити фибриногена. Процесс активации тромбоцитов, стимулированный напряжением сдвига, является одним из механизмов запуска тромбообразования под влиянием прокоагулянтов, выделяемых тромбоцитами [9].
Обычно тромбоциты способны активироваться и адгезировать к эндотелию в зонах разветвлений артерий. Р-селектин-зависимое соединение и роллинг вызывают взаимодействие кровяных пластинок синтактным эндотелием, но данный процесс никогда не бывает выраженным.
Соединение тромбоцитов с эндотелием возможно благодаря Р-селектину и PSGL-1 (P-selecting lycoproteinligand 1) на фоне активации тромбоцитов и тормозит их перемещение. В ходе роллинга отмечается также адгезия тромбоцитов с различными клетками, опосредуемая экспрессией Р-селектина. При этом, вследствие выброса оксида азота, простациклина и прочих дезагрегирующих соединений происходит распад сформировавшихся агрегатов и отвыв тромбоцитов от неповрежденного эндотелия.
На фоне повреждения сосуда развивается адгезия тромбоцитов к различным субэндотелиальным белкам. В условиях низкого напряжения сдвига, наблюдающегося при альтерации крупных артерий и вен, кровяные пластинки адгезируют прямо к оголенным коллагеновым волокнам благодаря коллагеновым рецепторам - GpVI,GpIV и GpIa/IIa. В этих условиях выраженность адгезии во многом определяется типом коллагена. В условиях контакта тромбоцитов с коллагеном V происходит примерно в 3 раза слабее, чем к коллагену I и III, и примерно в 1,5 раза слабее, чем к коллагену IV. При этом к коллагену V типа происходит адгезия в основном отдельных тромбоцитов, а к коллагену I и III наблюдается массовое прилипание с формированием крупных многослойных агрегатов. Это обеспечивает то, что только в случае глубокого повреждения сосудов с оголением коллагеновых волокон I и III типов возможно возникновение пристеночных тромбозов [1].
В условиях высокого напряжения сдвига в условиях наступления травмы мелких артерий и артериол процесс адгезии тромбоцитов обеспечивается фактором Виллебранда, находящегося в плазме и имеющего 3 активных центра - два из них соединяются с рецепторами тромбоцитов (GpIb), а один - со структурами субэндотелия.
По причине достаточно высокого содержания в субэндотелии фактора Виллебранда независимо от величины скорости сдвига, а также от уровня его в плазме и тромбоцитах в случае оголения субэндотелиальных волокон обязательно происходит адгезия тромбоцитов [9].
Кровяные пластинки адгезируют с фактором Виллебранда через рецептор GpIb/IX/V и непосредственно с коллагеном без фактора Виллебранда - через GpVI. Прочная адгезия тромбоцитов происходит через активированные интегриновые рецепторы GpIIb/IIIa (рецепторы к фибриногену) и рецептор к коллагену - α2β1, а также благодаря созданию «мостов» с участием αVβ3-интегрина, эндотелиальной ICAM-1 и GpIbα. При этом интегрины создают связь между экстрацеллюлярными мембранными протеинами и белками внутри тромбоцитов, обеспечивая эффективную двустороннюю сигнализацию.
В ходе адгезии тромбоцитов к субэндотелию они меняют свою форму, распластываются, вследствие чего развивается значительное увеличение их поверхности. Эти явления дают возможность возникнуть более обширным связям между рецепторами тромбоцитов и структурами субэндотелия с прочной фиксации на повреждённой сосудистой стенке кровяных пластинок и их агрегатов [7].
Течение агрегации помогает фактор активации тромбоцитов (РАF) и, кроме того, тромбин, всегда генерирующийся в месте повреждения сосудистой стенки. Под действием слабых индукторов (адреналин, АДФ, серотонин, фибронектинвитронектин) развивается экспрессия рецепторов на поверхности тромбоцитов к фибриногену (GpIIb/IIIa), вследствие чего при наличии в среде Са 2+ он соединяет друг с другом 2 прилегающих тромбоцита. В обеспечении данного процесса из всех адгезивных протеинов основная роль принадлежит фибриногену, являющимся главным кофактором агрегации ввиду того, что его концентрация в плазме наибольшая по сравнению с прочими белками, участвующими в адгезии и по причине достаточно большой аффинности к тромбоцитарным рецепторам (GpIb/IIa). Кроме того, симметричное строение фибриногена дает ему возможность вступать в двухсторонние связи с различными рецепторами на поверхности тромбоцитов, создавая мостики, связывающие их [2].
На начальном этапе агрегация носит обратимый характер, т.к. после нее может развиться частичное или полное разрушение агрегатов - процесс дезагрегации. Ввиду того, что связь между тромбоцитами порой непрочна, то определенное количество агрегатов способно отрываться и уноситься с кровью. Эта агрегация называется первичной, или обратимой, не обладающей возможностью полного и окончательного прекращения кровотечения даже из небольших сосудов.
Большое значение имеет вторичная (необратимая) агрегация, сопровождающаяся процессом тромбоцитарной секреции. Слабые индукторы, соединяясь со своими рецепторами, на тромбоцитах вызывают повышение цитоплазматического Са 2+ с активацией фосфолипазы А2, выщепляющей из мембран кровяных пластинок арахидоновой кислоты, которая превращается в PgG2, PgH2 и тромбоксан А2 (ТхА2), являющиеся сильными агонистами агрегации и вазоконстрикторами. Выбрасываясь из тромбоцитов PgH2PgG2 и ТхА2, повышают уровень экспрессии фибриногеновых рецепторов и усиливают сигнал, передаваемый внутрь кровяных пластинок. ТхА2 активирует фермент фосфолипазу С и стимулирует полифосфоинозитольный путь активации тромбоцитов. При этом ТхА2 способствует выделению Са 2+ в цитоплазму из плотной тубулярной системы, что активирует актом иозиновую систему и процесс фосфорилирования протеинов.
На фоне активации тромбоцитов наступает дополнительная экспрессия интегрина GpIIb/IIIa (αIIbβ3) на мембране тромбоцитов, являющегося наиболее важным для адгезии и агрегации. В этих условиях GpIb инициирует межтромбоцитарный контакт, тогда как GpIIb/IIIa сохраняет возникающие агрегаты. В этой связи главным моментом, регулирующим переход обратимой агрегации в необратимую, следует считать экспрессию GpIIb/IIIa, наступающую под действием АДФ, ТхА2 и тромбина.
Появление в крови тромбина ведет к активации протеиназоактивируемых рецепторов РАR-1 РАR-3, РАR-4, связанных с G-белками, что обеспечивает рост концентрации Са 2+ внутри кровяных пластинок и развитие их агрегации [3,7].
В результате адгезии, агрегации и ретракции из тромбоцитов выбрасываются находящиеся в гранулах биологически значимые вещества - РАF,АДФ, адреналин, фибриноген, норадреналин, ТхА2, фактор Виллебранда, фибронектин, витронектини тромбоспондин, что существенно укрепляет образовавшийся тромбоцитарный тромб. Вышедший из кровяных пластинок в ходе ретракции фактор роста способствует ускорению репарации поврежденной сосудистой стенки. Восстановлению проходимости сосуда обеспечивают выделяемые из тромбоцитарных γ-грануллизосомальные энзимы. При этом происходит синтез тромбина, стимулирующего распластывание и агрегацию тромбоцитов и вызывающий выпадение сети фибрина, в которой всегда задерживаются отдельные лейкоциты и эритроциты [10].
В ходе агрегации кровяные пластинки меняют свой состав и форму. В самом начале агрегации и возникновения первых порций тромбина поверхность тромбоцитов не испытывает значимых изменений, но в некоторых кровяных пластинках отмечаются признаки распада цитоплазмы. Спустя 40-90 сек. течения агрегации уже часть тромбоцитов становится мутной, их цитоплазма набухает, клетки плотно прилипают друг к другу, хотя повреждений мембраны найти не удаётся. Некоторые тромбоциты в этих условиях теряют гранулы, в прочих же число гранул остаётся без изменений. Спустя 2-3 мин. после появления тромбина тромбоциты набухают, их цитоплазма бледнеет, α-гранулы фрагментируются на мелкие пузырьки. В этих условиях гранулы постепенно перемещаются к центру кровяной пластинки и в ходе сокращения актомиозиновых комплексов сквозь мембрану выталкиваются в плазму. В последующем тромбоциты истончаются и несколько удлиняются. В их цитоплазме определяются только митохондрии, тогда как гранулы уже не выявляются. В этих условиях тромбоциты плотно примыкают друг к другу, а между ними находится в большом количестве фибрин. Изменения формы кровяных пластинок сопряжены с утратой мембранной асимметрии фосфолипидов. В этих условиях имеющие отрицательный заряд фосфолипиды - фосфатидилсерин, и отчасти фосфатидилэтаноламин, переходят на наружный листок мембраны тромбоцита, что обеспечивает условия, необходимые для свёртывания крови, полимеризации нитей фибрина и консолидации тромбоцитарной пробки, а в последующем ретракции тромбоцитарного конгломерата [1,9].
Таким образом, тромбоциты по праву считаются основой всего первичного гемостаза за счет их способности путем агрегации тромбировать локусы повреждения в сосудах. При формировании из них избыточного количества динамичных агрегатов могут меняться реологические свойства крови и тем самым состояние трофики тканей во всем организме.
Рецензенты:
Грушкин А.Г., д.б.н., профессор кафедры ветеринарии и физиологии животных Калужского филиала РГАУ-МСХА имени К.А.Тимирязева, г. Калуга;
Смахтин М.Ю., д.б.н., профессор кафедры биохимии Курского государственного медицинского университета, г. Курск.
Тромбоциты. Роль тромбоцитов в гемостазе.
Тромбоциты или кровяные пластинки (бляшки Биццоцеро) — имеют дисковидную форму диаметром от 2 до 5 мкм, толщиной около 0,5 мкм. В крови содержится (150-450) × 10 9 /л. Образуются в костном мозге путем отшнуровки фрагментов цитоплазмы мегакариоцита. Две трети тромбоцитов находятся в крови, остальные депонированы в селезенке (селезеночный пул): Продолжительность жизни тромбоцитов — 5-11 дней. Старые тромбоциты фагоцитируются в селезёнке, печени и костном мозге, а также служат источником питания для эндотелиоцитов большинства сосудов.
Строение. У тромбоцита нет ядра, центральная часть содержит фиолетовые гранулы, называется грануломером. Периферическая часть (гиаломер) содержит циркулярные пучки микротрубочек (необходимы для сохранения овальной формы тромбоцита), а также актин, миозин, гельзолин и другие сократительные белки, нужные для изменения формы тромбоцитов, их взаимной адгезии и агрегации, а также для ретракции образовавшегося при агрегации тромбоцитов сгустка крови. По периферии тромбоцита расположены также анастомозирующие мембранные канальцы, открывающиеся во внеклеточную среду и необходимые для секреции содержимого α‑гранул (рис. 3).
Строение тромбоцита
В цитоплазме рассеяны узкие, неправильной формы мембранные трубочки, составляющие плотную тубулярную систему; трубочки содержат циклооксигеназу (необходима для окисления арахидоновой кислоты и образования тромбоксана A2), необходимого для агрегации тромбоцитов.
В цитоплазме видны мелкие скопления гликогена и крупные гранулы нескольких типов:
α (альфа)-гранулы содержат различные белки и гликопротеины, принимающие участие в процессах свертывания крови: фибронектин (укрепляет тромб на поврежденной поверхности), фибриноген, антигепариновый фактор 4, тромбоцитарный фактор роста (стимулирует пролиферацию фибробластов, ускоряя заживление ран), фактор фон Виллебранда (WB) (после выхода в плазму способствует адгезии тромбоцитов), ингибитор активатора плазминогена (связывается с тканевым активатором плазминогена, создавая локальный антифибринолитический потенциал);
δ (дельта)-гранулы — небелковые гранулы высокой электронно-оптической плотности, содержат серотонин, гистамин, ионы Са 2+ , АДФ, АТФ. Кальций регулирует адгезию, образование тромбоксана А2, АДФ, способствует агрегации. Серотонин и катехоламины обеспечивают сокращение сосудов в местах их повреждений.
Третий тип мелких гранул — λ (лямбда)-гранулы — содержит лизосомные ферменты.
Выступающие наружу части молекул интегральных белков плазматической мембраны, богатые полисахаридными боковыми цепями (гликопротеины), создают внешнее покрытие липидного бислоя — гликокаликс. Здесь же адсорбированы факторы коагуляции и иммуноглобулины. На наружных частях гликопротеиновых молекул находятся рецепторные места. После их соединения с агонистами возникает сигнал активации, передающийся к внутренним частям периферической зоны тромбоцитов.
Так, гликопротеин Ib (GP Ib) важен для адгезии тромбоцитов, он связывается с фактором фон Виллебранда и подэндотелиальной соединительной тканью.
Гликопротеин IIb‑IIIa (GP IIb‑IIIa) — рецептор фибриногена, фибронектина, тромбоспондина, витронектина, фактора фон Виллебранда; эти факторы способствуют адгезии и агрегации тромбоцитов, опосредуя формирование между ними «мостиков» из фибриногена.
Тромбоцитарные факторы свертывания принято делить на эндогенные (образующиеся в самих тромбоцитах) и экзогенные (факторы плазмы, адсорбированные на поверхности тромбоцитов).
Эндогенные факторы тромбоцитов принято обозначать арабскими цифрами, в отличие от плазменных факторов, которые обозначаются римскими цифрами. Наиболее изучены 11 эндогенных тромбоцитарных факторов.
Фактор 1 тромбоцитов — тромбоцитарный акцелератор-глобулин — участвует в образовании протромбиназы и ускоряет образование тромбина из протромбина.
Фактор 2 тромбоцитов — акцелератор тромбина, фибринопластический фактор — ускоряет превращение фибриногена в фибрин.
Фактор 3 тромбоцитов (3ТФ) — тромбоцитарный тромбопластин, мембранный фосфолипидный фактор — представляет собой липопротеид. Служит матрицей для взаимодействия плазменных факторов гемокоагуляции, образования их активных комплексов. Необходим для эндогенного образования протромбиназы, способствующей превращению протромбина в тромбин. Фактор 3 выделяется при агрегации тромбоцитов.
Фактор 4 тромбоцитов — антигепариновый — обладает выраженной антигепариновой активностью. Снижение количества тромбоцитов повышает чувствительность крови к гепарину.
Фактор 5 тромбоцитов — свертываемый — по своим свойствам сходен с фибриногеном плазмы. Интенсивно выделяется из тромбоцитов под влиянием тромбина. Фактор 5 тромбоцитов принимает участие в агрегации тромбоцитов и тем самым способствует созданию прочного тромба.
Фактор 6 тромбоцитов — тромбостенин, представляет собой сократительный белок тромбоцитов, напоминающий актомиозин мышечных волокон. При сокращении тромбостенина происходит ретракция кровяного сгустка. При этом тромбоциты подтягиваются друг к другу, что в свою очередь приводит к сближению нитей фибрина. Сгусток обезвоживается, становится более компактным.
Фактор 7 тромбоцитов — пластиночный кофактор, котромбопластин, или активатор тромбопластина. Его аналог содержится в змеином яде. Роль котромбопластина в процессе свертывания крови в условиях нормы не ясна.
Фактор 8 тромбоцитов — антифибринолитический. Задерживает фибринолиз.
Фактор 9 тромбоцитов — фибринстабилизирующий фактор — вещество, аналогичное фактору XIII плазмы. Участвует в стабилизации фибрина (превращении растворимого фибрина в нерастворимый).
Фактор 10 тромбоцитов — серотонин, или сосудосуживающий фактор. Тромбоциты обогащаются серотонином при прохождении через сосуды желудочно-кишечного тракта и печени. Серотонин выделяется из тромбоцитов во время их агрегации, вызванной АДФ, адреналином, коллагеном. Серотонин обладает многими свойствами: усиливает сокращение сосудов и ретракцию кровяного сгустка, изменяет артериальное давление, является антагонистом гепарина; при тромбоцитопении способен нормализовать ретракцию кровяного сгустка и в присутствии тромбина ускорять переход фибриногена в фибрин.
Фактор 11 тромбоцитов — АДФ (аденозиндифосфат) — фактор агрегации тромбоцитов. При выходе на поверхность тромбоцитов АДФ способствует их склеиванию между собой. Кроме того, АДФ усиливает адгезию тромбоцитов к поврежденной стенке сосуда.
Значение тромбоцитов в организме.
1. Участвуют в гемостазе. В гемостазе тромбоциты осуществляют следующие функции:
ангиотрофическая — обеспечение жизнеспособности эндотелиальных клеток и поддержание нормальной структуры и функции стенок сосудов микроциркуляторного русла (нехватка тромбоцитов проявляется петехиальной сытью из-за резкого снижения прницаемости эндотелия);
ангиоспастическая — поддержание спазма поврежденных сосудов через секрецию серотонина, катехоламинов, β-тромбомодулина;
адгезивно-агрегационная — участие в первичном гемостазе путем образования тромбоцитарной пробки или белого тромба;
коагуляционная — участие в процессе свертывания крови и в регуляции фибринолиза (11 тромбоцитарных факторов, среди которых одним из наиболее активных является третий фактор тромбоцитов );
репаративная — ростовые факторы тромбоцитов стимулируют размножение и миграцию гладкомышечных клеток и эндотелиоцитов, что активирует процессы репарации в месте повреждения сосуда. Это обусловливает их участие в патогенезе атеросклероза, ишемической болезни сердца, реакции отторжения трансплантата, развитии опухолевых метастазов.
2. Транспортная — Депонирование и транспорт биологически активных веществ (перенос серотонина из мест синтеза и освобождения в тканях, его разрушение и др.)
3. Фагоцитоз — тромбоциты благодаря большой подвижности и образованию псевдоподий участвуют в иммунобиологических реакциях, способны фагоцитировать вирусы, иммунные комплексы и неорганические частички.
Тромбоцитоз — увеличение количества тромбоцитов в крови выше верхней границы физиологической нормы, отмечается при физическом напряжении, стрессе, после кровопотерь, при некоторых видах лейкозов и др.
Тромбоцитопения — уменьшение содержания тромбоцитов в крови ниже нижней границы физиологической нормы. Может развиваться вследствие деструкции тромбоцитов иммунной и неиммунной природы, при нарушении образования тромбоцитов (например, дефиците витамина В12, приеме некоторых лекарственных препаратов), при выраженном увеличении селезенки и др. Клинические признаки (кровоточивость десен, повышенную склонность к внутрикожным кровоизлияниям и др.) проявляются в случае снижения содержания тромбоцитов ниже 50х10 9 /л.
Тромбоцитопатия — состояние, характеризующееся нормальным количеством тромбоцитов, не способных адекватно участвовать в гемостазе (нарушения функции тромбоцитов).
Как тромбоциты ускоряют свёртывание крови
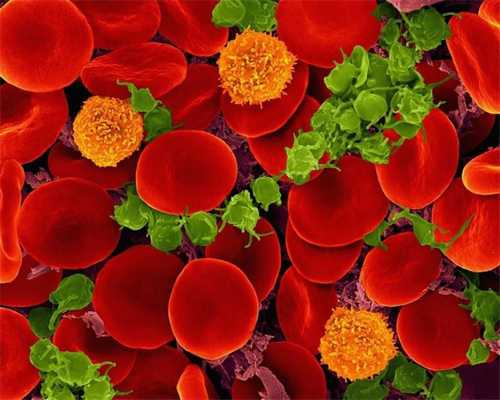
Если посмотреть на каплю крови в микроскоп (пусть это будет световой микроскоп, но достаточно мощный), то можно увидеть клетки трёх типов: многочисленные эритроциты, или красные кровяные тельца, немногочисленные, но довольно крупные лейкоциты и мельчайшие тромбоциты, которые удаётся разглядеть с некоторым трудом. Эритроциты, плотно набитые белком гемоглобином, переносят кислород: гемоглобин связывает его в лёгких и отдаёт в тканях и органах, которые в нём нуждаются. Лейкоциты — клетки иммунной системы, и они вместе с иммунными белками защищают нас от инфекций и от некоторых неинфекционных заболеваний, например от рака. Лейкоцитов существует несколько типов, отличающихся в том числе и по численности; возможно, из лейкоцитов нам попадутся Т-лимфоциты, которые целенаправленно распознают и сами уничтожают как чужеродные, так и наши собственные клетки, которым не повезло заболеть. Наконец, тромбоциты. Про тромбоциты мы знаем, что они нужны для свёртывания крови.
Кто не представляет, как работает система свёртывания крови? Уколовши палец, мы наблюдаем, как он сначала кровоточит, а потом перестаёт — образовавшийся тромб остановил кровь. Если бы кровь не свёртывалась, то разбитый нос мог бы оказаться смертельным ранением. Но, наверно, едва ли не более важная функция механизма свёртывания — предотвращение внутренних кровотечений, которые часто случаются при различных заболеваниях (например, при тяжёлой инфекции или при злокачественной опухоли). При этом система свёртывания должна быть очень точно сбалансирована: если она будет работать плохо, то пойдут неостанавливаемые кровотечения, внутренние и наружные; если же механизм свёртывания будет слишком активным, начнут формироваться тромбы, грозящие закупоркой сосудов и остановкой кровоснабжения. В медицине есть масса примеров, когда процессы свёртывания крови и тромбообразования идут не так, как надо, и не там, где надо. Причиной тому могут быть либо другие болезни, и тогда нарушения системы свёртывания крови — это просто сопутствующий симптом, либо же сами эти нарушения представляют собой отдельные, самостоятельные заболевания (вроде небезызвестной гемофилии или болезни Виллебранда).
Бороться с аномалиями системы свёртывания можно по-разному, и сейчас есть медицинские средства, которые позволяют эффективно регулировать её работу. Но чтобы такие средства работали ещё лучше, чтобы сделать их ещё более совершенными, нужно как можно точнее знать, как на молекулярно-клеточном уровне устроен механизм свёртывания крови. Его изучают уже более ста лет, и сейчас его схему можно найти в любом школьном учебнике; правда, схему эту большинство из нас старается забыть, как страшный сон: ещё бы, около двух десятков белков, соединённых стрелками, — кто-то кого-то активирует, кто-то кого-то ингибирует. Однако если рассматривать свёртывание по этапам, то всё становится более или менее понятно.
Стоит сразу сказать, что собственно свёртывание — лишь часть более общего процесса гемостаза (от греч. haimatos — кровь, stasis — остановка). И этот процесс начинается как раз с тромбоцитов. Они происходят от мегакариоцитов — гигантских клеток костного мозга. От зрелых мегакариоцитов «отшнуровываются» куски цитоплазмы, которые и становятся безъядерными клетками тромбоцитами (хотя, учитывая их происхождение и отсутствие ядра, более корректно называть их просто тельцами крови или кровяными пластинками). Тромбоциты циркулируют по крови, пока не «заметят» брешь в сосуде. Сигналом для них служит соединительнотканный белок коллаген. Он обычно спрятан внутри стенки сосуда, но при её повреждении оказывается лицом к лицу с тромбоцитами и другими белками крови. На мембране тромбоцитов есть специальный рецептор, который хватает коллаген и заставляет кровяные пластинки прилипнуть к месту повреждения. Тут в дело вступает один из факторов свёртывания под названием «фактор фон Виллебранда». Это гликопротеин (его молекула состоит из белковой и углеводной частей), который помогает другим рецепторам тромбоцитов зацепиться за торчащий из стенки сосуда коллаген. Благодаря фактору фон Виллебранда тромбоциты не только прочнее взаимодействуют с местом повреждения, но и дополнительно активируются — подают молекулярные сигналы другим тромбоцитам и белкам свёртывания, меняют внешнюю форму и активно слипаются друг с другом. В результате на стенке кровеносного сосуда появляется затычка из тромбоцитов.
Одновременно с формированием тромбоцитарной пробки происходит процесс собственно свёртывания крови — свёртывания в строгом смысле слова. В нём участвует множество белков плазмы крови, большинство из них — ферменты-протеазы, то есть белки, отщепляющие куски от других белков. Если до расщепления «жертва» протеазы была неактивным белком-ферментом, то после расщепления фермент активируется и, если он сам протеаза, тоже может кого-то расщепить. Суть ферментативных реакций, которые идут во время свёртывания, в том, что белки активируют друг друга, и в итоге всё заканчивается появлением активного белка фибрина, который быстро полимеризуется, превращаясь в нити — фибриллы. Из нитей фибрина формируется фибриновый сгусток, дополнительно укрепляющий тромбоцитовую «затычку», — образуется тромбоцитарно-фибриновый тромб. Когда сосуд восстанавливается, тромб рассасывается.
Оба этапа — и формирование тромбоцитарной пробки, и свёртывание крови с участием плазматических факторов-ферментов — подчиняются множеству регуляторов. Для организма важно, чтобы система гемостаза работала как можно более точно, и многостадийность как раз помогает выполнять тонкую настройку: на каждом этапе, на каждой реакции ферменты и другие молекулы, задействованные в процессе, проверяют, не ложный ли сигнал к ним пришёл и действительно ли есть необходимость в тромбе. Естественно, тромбоциты и факторы свёртывания теснейшим образом связаны друг с другом и тромбоциты нужны не только для того, чтобы первыми заткнуть брешь в сосуде. Во-первых, они также выделяют белки, которые ускоряют восстановление стенки сосуда. Во-вторых, что особенно важно, кровяные пластинки нужны ещё для того, чтобы ферменты свёртывания продолжали работать.
После запуска процесса гемостаза мембрана некоторых тромбоцитов изменяется особым образом, так что теперь на неё могут садиться ферменты реакций свёртывания: после приземления на такие тромбоциты они начинают работать намного быстрее. Что при этом происходит, удалось выяснить лишь относительно недавно. Активированные тромбоциты, то есть те, которые почувствовали повреждение сосуда, бывают двух форм: простые (агрегирующие) и сверхактивированные (прокоагулянтные). Простые агрегирующие тромбоциты отчасти похожи на амёбы: они образуют выпячивания мембраны, похожие на ножки, которые помогают им лучше сцепляться друг с другом, и становятся более плоскими, как бы растекаясь по поверхности. Такие клетки формируют основное тело тромба. Сверхактивированные тромбоциты ведут себя иначе: они приобретают сферическую форму и увеличиваются в несколько раз, становясь похожими на воздушные шарики. Они не просто укрепляют тромб, но и стимулируют реакцию свёртывания, почему их и называют прокоагулянтными.
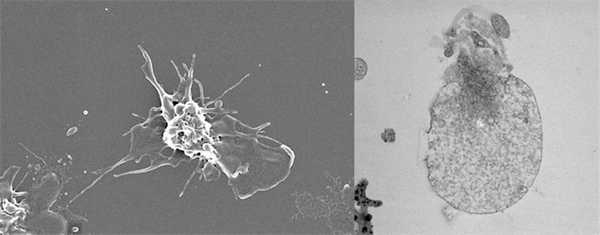
Почувствовав, что стенка кровеносного сосуда повреждена, тромбоциты активируются. Обычные активированные тромбоциты (слева; фото со сканирующего электронного микроскопа) делаются плоскими и образуют, подобно амёбам, многочисленные выросты мембраны; сверхактивированные, или прокоагулянтные, тромбоциты (справа; фото с просвечивающего электронного микроскопа) превращаются в шар и становятся площадкой для сборки и активации ферментативных комплексов аппарата свёртывания крови. Фото Михаила Пантелеева, МГУ
Как одни тромбоциты становятся простыми, а другие — сверхактивированными? Известно, что в прокоагулянтных тромбоцитах очень высок уровень кальция (ионы кальция вообще один из главных регуляторов гемостаза) и что у них выходят из строя митохондрии. Связаны ли эти изменения в клеточной физиологии со сверхактивацией тромбоцитов?
В прошлом году Фазли Атауллаханов * , директор Центра теоретических проблем физико-химической фармакологии РАН, вместе с Михаилом Пантелеевым, заведующим лабораторией молекулярных механизмов гемостаза центра и профессором кафедры медицинской физики физического факультета МГУ, опубликовали в журнале Molecular BioSystems статью с описанием модели митохондриального некроза как особой формы клеточной смерти. Мы знаем, что клетка может погибнуть в результате апоптоза, включив программу самоуничтожения (при апоптозе всё происходит по плану и с минимальным беспокойством для клеток-соседей), или в результате некроза, когда гибель случается быстро и незапланированно, например из-за разрыва наружной мембраны или из-за масштабных внутренних неприятностей, вроде вирусной или бактериальной инфекции.
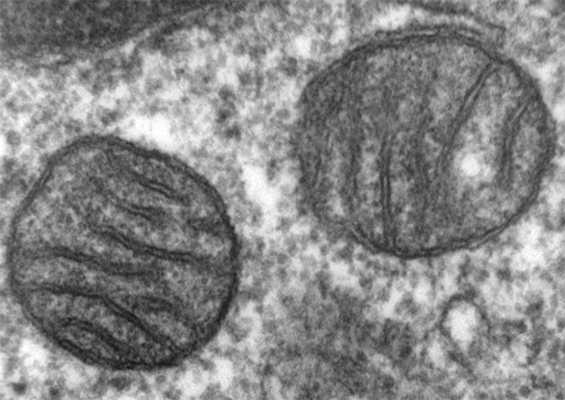
Поперечный срез через митохондрии клетки лёгкого. Митохондрии выглядят как цистерны, окружённые двойной мембраной; поперечные полосы на снимке — впячивания их внутренней мембраны, на которой сидят ферменты энергетического обмена веществ. Фото: Louisa Howard/Wikimedia Commons
В чём особенность митохондриального некроза? Митохондрии, как известно, служат источниками энергии для любой нашей клетки: в митохондриях происходит кислородное окисление «питательных» молекул, а освобождённая при этом энергия запасается в удобной для клетки форме. Побочным продуктом при работе с кислородом оказываются агрессивные кислородные радикалы, которые могут испортить любую биомолекулу. Сами митохондрии стараются уменьшать концентрацию радикалов и не выпускать их из себя в клеточную цитоплазму.
При митохондриальном некрозе происходит следующее: митохондрии вбирают в себя кальций, и в какой-то момент, когда кальция становится слишком много, они разрушаются, выплёскивая в цитоплазму и кальций, и активные формы кислорода. В результате в клетке распадается внутриклеточный белковый скелет и клетка сильно увеличивается в объёме, превращаясь в шар. (Как мы помним, шарообразная форма характерна для сверхактивированных тромбоцитов.) Кроме того, и ионы кальция, и активные формы кислорода активируют фермент скрамблазу, который перебрасывает фосфатидилсерин — один из липидов цитоплазматической мембраны — из внутреннего слоя мембраны в наружный. И вот на такую модифицированную мембрану округлившихся тромбоцитов, обогащённую фосфатидилсерином, прилипают некоторые важные факторы свёртывания: здесь они собираются в комплексы, активируются, и в результате реакция свёртывания ускоряется в 1000-10 000 раз.
В новой статье, опубликованной в июне этого года в Journal of Thrombosis and Haemostasis, Михаил Пантелеев, Фазли Атауллаханов и их коллеги описывают эксперименты, которые полностью подтверждают такую модель активации тромбоцитов: кровяные пластинки стимулировали тромбином, одним из белков системы свёртывания, после чего митохондрии наполнялись ионами кальция, а в митохондриальных мембранах появлялись поры. Проницаемость митохондрий увеличивалась, и в какой-то момент, когда изменение проницаемости делалось необратимым, весь запасённый кальций оказывался в цитоплазме и запускал процесс «переформатирования» наружной мембраны.
Получается следующая картина: тромбоциты, подчиняясь внешним активаторам, впитывают кальций. Из их цитоплазмы кальций переходит в митохондрии. В самой цитоплазме уровень ионов кальция то повышается, то понижается (осциллирует), но в митохондриях он неуклонно растёт, и наступает момент, когда они уже не могут удерживать кальциевые ионы внутри себя. Весь кальций (с кислородными окислителями) выходит в цитоплазму и включает фермент, перебрасывающий липиды в цитоплазматической мембране тромбоцита. В результате на поверхности сверхактивированного и, очевидно, доживающего свои последние минуты тромбоцита собираются ферментативные комплексы, ускоряющие реакцию свёртывания.
Почему же не все тромбоциты становятся сверхактивированными — прокоагулянтными? Вероятно, потому, что для активации требуется сумма сигналов от разных регуляторов. Мы уже сказали, что тромбоциты чувствительны к тромбину, который плавает в плазме крови, а в начале статьи говорили, что одним из первых активирующих сигналов для кровяных пластинок служит коллаген из повреждённой стенки сосуда. Коллаген и тромбин действительно сильные активаторы, но кроме них тромбоциты «прислушиваются» и к некоторым другим молекулам. Степень активации зависит от количества разных входных сигналов, и превращение в прокоагулянтную форму, очевидно, происходит тогда, когда суммарный сигнал извне оказывается для конкретного тромбоцита особенно сильным.
Практические аспекты полученных результатов понятны каждому: чем больше подробностей узнаем про свёртывание крови, тем скорее научимся управлять этим процессом, ускоряя или замедляя его в соответствии с медицинскими показаниями.
Читайте также:
- Признаки опухоли, гемангиомы на КТ глаза и орбиты
- Возбудитель Лайм боррелиоза. Механизм развития Лайм-боррелиоза
- Частота резекции легкого с пневмоперитонеумом. Пневмоперитонеум в дооперационном периоде
- Рубцовая алопеция. Причины выпадения волос и диагностика
- Закрытый перелом: причины, симптомы и лечение