
Чем может быть представлен генетический аппарат вирусов
Вирусам, как и всем живым организмам, свойственны наследственность и изменчивость. Основной особенностью вирусного генома является то, что наследственная информация у вирусов может быть записана как на ДНК, так и на РНК. Геном ДНК-содержащих вирусов двухнитевой (исключение составляют парвовирусы, имеющие однонитевую ДНК), несегментированный и проявляет инфекционные свойства. У вирусов, принадлежащих к родам PoxvirusиHepadnavirusгеном представлен двумя цепочками ДНК разной длины. Геном большинства РНК-содержащих вирусов однонитевой (исключение составляют реовирусы и ретровирусы, обладающие двунитевыми геномами) и может быть сегментированным (представители родовRetrovirus,Orthomyxovirus,ArenavirusиReovirus) или несегментированным.
У другой группы вирусов РНК не способна транслировать генетическую информацию непосредственно на рибосомы и функционировать как иРНК. Такие РНК служат матрицей для образования иРНК, т.е. при репликации первоначально синтезируется матрица (+РНК) для синтеза –РНК. Такой тип РНК определяют как минус-нить и обозначают –РНК (негативный геном). У вирусов этой группы репликация РНК отличается от транскрипции по длине образующихся молекул: при репликации длина РНК соответствует материнской нити, а при транскрипции образуются укороченные молекулы иРНК. Молекулы +РНК проявляют инфекционность, а –РНК не проявляют инфекционные свойства и для воспроизведения должны транскрибироваться в +РНК.
Исключение составляют ретровирусы, которые содержат однонитевую +РНК, служащую матрицей для вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). При помощи этого фермента информация переписывается с РНК на ДНК, в результате чего образуется ДНК-провирус, интегрирующий в клеточный геном.
Так же как и у прочих форм жизни нуклеиновые кислоты вирусов подвержены мутациям. Фенотипически мутации вирусного генома проявляются изменениями в антигенной структуре, неспособности вызывать продуктивную инфекцию в чувствительной клетке, термостабильностью, изменением размера и формы бляшек, образуемых под агаровым покрытием. Большинству мутаций присущи реверсии к дикому типу, причем каждая мутация имеет характерную частоту реверсий, которую можно точно измерить. У вирусов выделяют спонтанные и индуцированные мутации.
Скорость спонтанного мутагенеза в ДНК-геномах значительно ниже (10 -8 – 10 -11 на каждый включенный нуклеотид), чем у РНК-геномных (10 -3 – 10 -4 на каждый включенный нуклеотид). Более высокая частота спонтанных мутаций связана с низкой точностью репликации РНК-геномов, которая вероятно связана с отсутствием у РНК-репликаз корректирующей активности, свойственной ферментам, реплицирующим ДНК. Наиболее часто спонтанные мутации наблюдаются у ретровирусов, что связано с более высокой частотой сбоев в обратной транскрипции, не способных к самокоррекции.
Индуцированные мутации у вирусов получают при действии различных химических и физических мутагенов, которые подразделяют на действующие invivoиinvitro.
Вирусные мутации классифицируют по изменениям фенотипа и генотипа. По фенотипическим проявлениям мутации вирусов разделяют на четыре группы:
Мутации, не имеющие фенотипического проявления.
Летальные мутации, т.е. полностью нарушающие синтез или функцию жизненно важных белков и приводящие к утрате способности к репродукции.
Условно летальные мутации, т.е. мутации с потерей способности синтезировать определенный белок или с нарушением его функции только в определенных условиях.
Мутации, имеющие фенотипическое проявление, например изменение размеров бляшек под агаровым покрытием или термостабильности.
По изменению генотипа мутации подразделяют на точечные (локализующиеся в индивидуальных генах) и генные (затрагивающие более обширные участки генома).
Заражение вирусами чувствительных клеток носит множественный характер, т.е. в клетку проникает сразу несколько вирионов. При этом вирусные геномы в процессе репликации могут кооперироваться или интерферировать. Кооперативные взаимодействия между вирусами представлены генетическими рекомбинациями, генетической реактивацией, комплементацией и фенотипическим смешиванием.
Генетическая рекомбинация чаще встречается у ДНК-содержащих вирусов или РНК-содержащих вирусов с фрагментированным геномом (вирус гриппа). При генетической рекомбинации происходит обмен между гомологичными участками вирусных геномов.
Генетическая реактивация наблюдается между геномами родственных вирусов с мутациями в разных генах. При перераспределении генетического материала формируется полноценный геном.
Комплементация происходит когда один из вирусов, инфицирующих клетку, в результате мутации синтезирует нефункциональный белок. Немутантный вирус, синтезируя полноценный белок, восполняет отсутствие его у мутантного вируса.
Фенотипическое смешивание происходит при смешанном заражении чувствительной клетки двумя вирусами, когда часть потомства приобретает фенотипические признаки, присущие двум вирусам, при неизменном генотипе.
При множественном инфицировании чувствительной клетки между вирусами могут возникать интерферирующие взаимодействия. Интерфернцией вирусов называют состояние невосприимчивости к вторичному заражению клетки, уже инфицированной вирусом. При гетерологической интерференции инфицирование одним вирусом полностью блокирует возможность репликации второго вируса в пределах одной клетки. Механизмы гетерологической интерференции связаны с угнетением адсорбции другого вируса путем блокирования или разрушения специфических рецепторов, а так же с ингибированием трансляции мРНК любой гетерологичной мРНК в инфицированной клетке. Кроме того, первичное заражение может индуцировать образование интерферона, ингибирующего репликацию второго вируса.
Гомологическая интерференция, т.е. интерференция между гомологичными вирусами, характерна для многих вирусов, особенно при повторных пассажах invitroи при высокой множественности инфицирования. В таких условиях образуется много дефектных вирусных частиц, обычно не способных к репродукции. Однако размножение дефектных вирусов возможно при совместном заражении с полноценным вирусом (вирус-помощник). При этом дефектный вирус может вмешиваться в репликативный цикл вируса-помощника и образовывать дочерние дефектные интерферирующие (ДИ) вирусные частицы. ДИ-частицам присущи три основных свойства: дефектность (повреждение в важных генах), способность к интерференции (ДИ-частицы препятствуют репликации полноценного вируса или других гомологичных вирусов) и способность к самообогащению за счет стандартного вируса. Циркуляция ДИ-частиц и коинфекция с полноценным вирусом вызывают вялотекущие, длительные формы инфекции.
Кроме взаимодействий, происходящих между вирусами, при смешанной инфекции происходят также взаимодействие между вирусом и клеткой-хозяином. При взаимодействии клеток с ДНК-содержащими вирусами может происходить вирусная трансформация клетки. В результате трансформации у клеток изменяются морфологические, биохимические и ростовые характеристики, может появляться способность к опухолевому росту. Представляет интерес трансформация клеток под действием РНК-геномных ретровирусов. У ретровирусов трансформация и репликация не являются взаимоисключающими, поскольку трансформированные клетки способны реплицировать вирус. Геномы трансформирующих вирусов обычно интегрируют с геномом трансформируемой клетки.
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Неклеточные формы жизни: вирусы
Вирус (лат. virus — яд) — неклеточные структуры с упорядоченной организацией, содержащие генетический материал (ДНК или РНК), упакованный в белковую оболочку, или капсид. Вирусы являются внутриклеточными паразитами на генетическом уровне. Они способны проникать в клетки живых организмов и в них размножаться. Для построения своих новых частиц они используют химические вещества и энергию клетки-хозяина. Форма вируса (палочковидная, сферическая, нитевидная) зависит от характера взаимодействия нуклеиновой кислоты с белковой оболочкой.
Вирусы в природе распространены повсеместно. Они паразитируют на всех группах организмов. С вирусной инфекцией связаны многие заболевания человека, в том числе крайне опасные (вирусный гепатит, СПИД, полимиелит, грипп и другие).
Вирусы существуют в двух формах: покоящейся (внеклеточной) и репродуцирующейся (внутриклеточной).
Вирусы условно делятся на простые и сложные.
- Простые. В составе таких вирусов только нуклеиновые кислоты (РНК и ДНК) и белок.
- Сложные. Нуклеотид этих вирусов состоит из белка и только РНК, также они могут содержать липопротеидную мембрану, углеводы и ферменты. К группе сложных вирусов относят так называемые ретровирусы . У них обнаружен такой фермент, как обратная транскриптаза.
Особенности вирусов
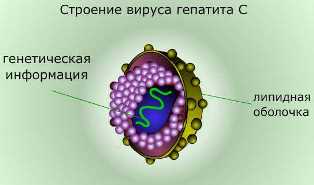
1. Тело вируса не имеет клеточного строения.
2. Вирусы могут существовать только как внутриклеточные паразиты и не могут размножаться вне клеток организма хозяина.
3. В вирусах содержится один тип нуклеиновых кислот — либо РНК, либо ДНК (все клеточные организмы содержат и ДНК, и РНК одновременно). Отсутствуют рибосомы.
Генетический аппарат вирусов представлен различными формами нуклеиновых кислот, такого разнообразия нет ни у одной из других форм жизни. У всех живых организмов, кроме вирусов, генетический аппарат состоит из двунитевой молекулы дезоксирибонуклеиновой кислоты (ДНК), а рибонуклеиновая кислота (РНК), выполняющая в клетках роль переносчика информации, всегда однонитевая.
У вирусов же существуют все возможные варианты устройства генетического аппарата: одно- и двунитевая РНК, одно- и двунитевая ДНК.
При этом и вирусная РНК, и вирусная ДНК могут быть либо линейными, либо замкнутыми в кольцо.
4. Отсутствует обмен веществ. Вирусом используется энергия, получаемая за счет обмена веществ в клетках хозяина. Имеют очень ограниченное число собственных ферментов, используют обмен веществ хозяина, его ферменты, энергию, полученную при обмене веществ в клетках хозяина.
Вирусы состоят из нуклеиновой кислоты, окруженной белками одного или нескольких типов; некоторые вирусы имеют также липидсодержащую внешнюю оболочку.
Белки вирусов выполняют тот же ряд важнейших функций, что и в клетке, в том числе структурную, транспортную, ферментативную, защитную.
Одной из основных особенностей строения вирусов является белковая оболочка (капсула), в которую заключен генетический материал вируса. При этом вирусы не имеют собственных белоксинтезирующих систем, а используют для этого системы клетки хозяина.
Вирусный белок выполняет защитную роль и весьма важную функцию — он отвечает за прикрепление вируса к поверхности клеток, после чего вирус проникает внутрь клетки и начинает размножаться в ней. Поэтому фрагмент белка, связывающийся с клеточной мембраной, остается неизменным.
Гемагглютинин — поверхностный белок вируса гриппа, обеспечивающий способность вируса присоединяться к клетке-хозяину.
Нейраминидаза — поверхностный белок вируса гриппа, отвечающий, во-первых, за способность вирусной частицы проникать в клетку, и, во-вторых, за способность вирусных частиц выходить из клетки после размножения.
Нуклеокапсид — генетический материал (РНК) вируса заключенный в белковую оболочку (капсулу).
Как результат присутствия в молекулах нескольких функциональных групп белки обладают высокой реактивной способностью и амфотерными свойствами.
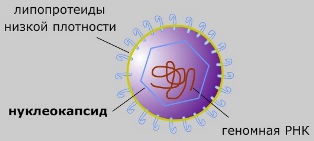
Капсид (уклеокапсид)— это внешняя оболочка вируса, состоящая из белков.
- Защита генетического материала (ДНК или РНК) вируса от механических и химических повреждений.
- Определение потенциала к заражению клетки.
- Прикрепление к клеточной мембране, разрыв мембраны и внедрение в клетку генетического материала вируса.
Капсиды большинства вирусов имеют спиральную или икосаэдрическую симметрию. В случае спиральной симметрии (например, у вируса табачной мозаики) составные части капсида формируют цилиндр из уложенных по спирали белковых глобул, внутри которого находится генетический материал вируса. В случае икосаэдрической симметрии (например, у многих бактериофагов) образуется квази-сферическая структура капсида.
Структурный анализ основных типов капсидов используется в классификации вирусов.
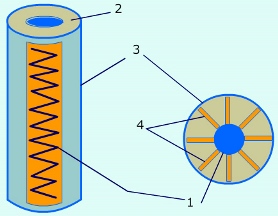
Дополнительная липопротеидная оболочка образована из плазматической мембраны клетки-хозяина и встречается только у сравнительно больших вирусов (грипп, герпес).
Схематичное строение вируса: 1 — сердцевина (однонитчатая РНК); 2 — белковая оболочка (капсид); 3 — дополнительная липопротеидная оболочка; 4 — капсомеры (структурные части капсида)
В отличие от других организмов вирусы не имеют рибосом и ферментов, катализирующих образование макроэргических фосфатов, метаболизм белков, углеводов и жиров.
Вирусы размножаются только внутри зараженных клеток и поэтому относятся к облигатным внутриклеточным паразитам.
Вирусные гены обычно кодируют белки, необходимые для репликации нуклеиновой кислоты и сборки вирусов.
Вироиды — субвирусные инфекционные агенты, возбудители некоторых заболеваний (в первую очередь у растений). Представляют собой высокоструктурированные кольцевые фрагменты РНК, реплицируемые клеточной РНК-полимеразой. Белков вироиды не кодируют. Вироиды были открыты и названы в 1971 году Теодором О. Динером.
Вирусоиды похожи на вироиды, но включены в структуру вируса — помощника и реплицируются только с его помощью.
Человек может заразиться прионами, содержащимися в пище, так как они не разрушаются ферментами пищеварительного тракта. Беспрепятственно проникая через стенку тонкого кишечника, они в конечном итоге попадают в центральную нервную систему. Так переносится новый вариант болезни Крейтцфельдта — Якоба, которой люди заражаются после употребления в пищу говядины, содержащей нервную ткань из голов скота, больных бычьей губчатой энцефалопатией (BSE, коровье бешенство).
Мозговое вещество, изъеденное прионом коровьего бешенства. Дырки и обширные пустые участки располагаются на месте бывших нервных клеток
Прионы могут проникать в тело и парентеральным (через внедрение патогенных микроорганизмов в организм человека или животного минуя пищевой тракт (через кровяное русло, кожу, конъюнктиву глаза, подкожно, внутримышечно, внутрибрюшинно) путем. Были описаны случаи заражения при внутримышечном введении препаратов, изготовленных из человеческих гипофизов (главным образом гормоны роста для лечения карликовости), а также заражение мозга инструментами при нейрохирургических операциях, поскольку прионы устойчивы к применяемым в настоящее время термическим и химическим методам стерилизации.
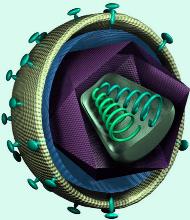
Ретровирусы — это вирусы с необычным способом репликации генетического материала. Для цикла репродукции этого большого семейства вирусов характерен обратный поток генетической информации: вместо обычной транскрипции (т. е. переписывания) дезоксирибонуклеиновой кислоты (ДНК) в рибонуклеиновую кислоту (РНК), как это происходит в клетке при реализации генетической информации, их геномная РНК переписывается в ДНК (обратная транскрипция).
Ретровирусы (лат. retro — обратно, назад и virus — яд) — РНК — содержащие вирусы, цикл размножения которой проходит через стадию двухцепочечной ДНК. В каждой вирусной частице имеются две копии вируса. В составе генома помимо генов, кодирующих структуру белков капсида, имеется ген, кодирующий фермент обратную транскриптазу, который осуществляет синтез ДНК на РНК. Обратная транскриптаза вместе с РНК упаковывается в вирусную частицу.
В цитоплазме клетки-хозяина фермент превращает одноцепочечную РНК в двухцепочечную ДНК, которая встраивается в геном хозяина. Провирусная ДНК транскрибируется как матрица для синтеза вирусных белков и в качестве генома при упаковке вирусных частиц.
Члены семейства ретровирусов вызывают ряд тяжелых заболеваний животных и человека. К наиболее изученным вирусам относятся вирусы лейкемии птиц, мышей, кошек и приматов, а также вирусы иммунодефицита кошек, обезьян и человека. Вирус иммунодефицита человека (ВИЧ) вызвал пандемию ВИЧ-инфекции и СПИДа (синдрома приобретенного иммунодефицита) во всем мире.
Первый Lentivirinae ( lente — медленно) — ленивый ретровирус был открыт в 1904 году, когда французы А. Балле и А. Карре обнаружили фильтрующийся агент — вирус инфекционной анемии лошадей. Затем были открыты другие лентивирусные инфекции сельскохозяйственных животных. Типичными лентивирусами являются давно изученые вирус висны у овец, кошачий вирус иммунодефицита, вирус артрита у коз. Кроме сходства по своему строению эти вирусы вызывают однотипные патологии. Например, заражение овец вирусом висны приводит к длительному хроническому заболеванию, которое тянется порой до 2 и более лет. Но затем, также как и при инфицировании ВИЧ, неизбежно наступает летальный исход.
ВИЧ-1 и ВИЧ-2 — единственные патогенные для человека представители подсемейства Lentivirinae. Говоря о медленном течении лентивирусных инфекций, их обычно сравнивают с острыми вирусными инфекциями (например, с гриппом), но не с инфекциями, вызываемыми другими ретровирусами. В то же время клиническая картина острой лихорадочной фазы ВИЧ-инфекции напоминает проявления многих классических острых инфекций.
Лекция 7
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ГЕНОМА ВИРУСА
Вирусы являются одним из излюбленных объектов молекулярной генетики благодаря простому строению и малой молекулярной массе их геномов, которая в 10 6 раз меньше массы генома эукариотической клетки. Организация генетического аппарата у ряда вирусов, например у SV40, настолько сходна с таковой генов эукариотической клетки, что получила название минихромосомы. Минихромосома широко используется для изучения организации и репликации ДНК.
Число генов у вирусов значительно варьирует: от 3—4 генов у просто устроенных вирусов (парвовирусы) до 150 генов и больше у сложно устроенных (вирус оспы). Геном вирусов животных является гаплоидным, за исключением ретровирусов, которые имеют диплоидный геном, представленный двумя идентичными молекулами РНК. У вирусов с фрагментарным геномом (вирусы гриппа, реовирусы) каждый фрагмент обычно представляет собой один ген.
Так же, как и геном эукариотической клетки, ДНК-геном ряда вирусов животных имеет мозаичную структуру, при которой смысловые последовательности чередуются с неинформативными последовательностями. Механизм сплайсинга при формировании иРНК широко распространен и среди вирусов, имеющих ядерную локализацию транскрипции (адено-, папова-, герпесвирусы), поскольку ферменты, осуществляющие сплайсинг, находятся в ядре. Однако сплайсинг был обнаружен и у РНК-содержащих вирусов. Например, у вирусов гриппа происходит сплайсинг транскриптов 7-го и 8-го генов; в результате сплайсинга и сдвига рамки трансляции продуктами каждого из этих генов являются по два уникальных белка.
В составе генов ДНК-содержащих вирусов есть регуляторные участки, в том числе промотор, контролирующие функцию структурных генов. Сильными промоторами являются концы многих вирусных ДНК, представляющие собой длинные концевые повторы, сильный промотор имеют гены тимидинкиназы вирусов оспы и герпеса. Эти промоторы используются в генной инженерии для усиления транскрипции изучаемого гена.
СПОСОБЫ УВЕЛИЧЕНИЯ ИНФОРМАЦИОННОЙ ЕМКОСТИ ВИРУСНОГО ГЕНОМА
У многих вирусов молекулярная масса синтезирующихся белков превышает теоретически рассчитанную. Этот феномен объясняется наличием у вирусов механизмов, позволяющих получить развернутую генетическую информацию при максимальной экономии генетического материала; подобные механизмы выработаны в процессе эволюции вирусов как генетических паразитов.
Способами увеличения генетической информации являются: 1) двукратное считывание одной и той же иРНК, но с другого инициирующего кодона; 2) сдвиг рамки трансляции; 3) сплайсинг; 4) транскрипция с перекрывающихся областей ДНК и др.
Трансляция может происходить без сдвига рамки и со сдвигом рамки. Генетический код является триплетным, это означает, что три нуклеотида, составляющих триплет, или кодон, кодируют одну аминокислоту. В том случае, если триплеты сохранены и генетический код не изменился, то при трансляции с двух разных инициирующих кодонов будут синтезироваться полипептиды, представляющие собой укороченную копию первого полипептида (трансляция без сдвига рамки).
В том случае, если произошел сдвиг на один или два нуклеотида, образуются новые триплеты (кодоны) и появляется новый генетический код. В этом случае одна молекула иРНК может транслироваться с образованием двух уникальных белков, т. е. таких белков, у которых нет идентичных аминокислотных последовательностей.
Сплайсинг со сдвигом рамки широко используется у ряда вирусов (вирусы гриппа, парамиксовирусы, буньявирусы, аденовирусы, паповавирусы, парвовирусы и др.). Например, все три иРНК аденоассоциированного вируса образуются при транскрипции одного гена и имеют общий З'-конец; самая короткая иРНК образуется путем сплайсинга и транслируется с образованием трех структурных белков, остальные две иРНК транслируются с образованием неструктурных белков. В результате сплайсинга и сдвига рамки иРНК 7-го и 8-го генов вируса гриппа транслируются с образованием двух белков: полипептидов М, и М2 (продукты 7-го гена) и NSi и NS2 (продукты 8-го гена). Белки NSt и NS2 имеют лишь первые 10 идентичных аминокислот, а затем — уникальные аминокислотные последовательности. Один и тот же ген парамиксовирусов (вирус Сендай) кодирует два уникальных белка: структурный белок Р и неструктурный белок С.
Одним из способов экономии генетического материала является нарезание полипептида-предшественника на участки разной длины, в результате чего образуются разные полипептиды с перекрывающимися аминокислотными последовательностями. Подобный механизм нарезания имеет место у аденоассоциированных вирусов и у SV40.
Таким образом, число реальных генов превосходит молекулярную массу генома. Основанный на длине генома расчет числа генов неизменно приведет к ошибочным результатам. Более точные представления о числе генов можно получить путем биохимического и генетического анализов.
ОСНОВНЫЕ ПРОЦЕССЫ, КОНТРОЛИРУЮЩИЕ НАСЛЕДСТВЕННОСТЬ И ИЗМЕНЧИВОСТЬ ВИРУСОВ
Модификации. Модификациями называются не наследуемые (фенотипические) изменения у вирусов, обусловленные клеткой-хозяином. Эти изменения лежат в основе адаптации вируса к новому хозяину и преодоления зависимого от хозяина ограничения. Модификации нуклеиновых кислот вирусов осуществляют клеточные ферменты, ответственные за ограничение (рестрикцию) репродукции вируса.
Мутации. В основе изменчивости вирусов лежат мутации, т. е. изменения состава и последовательностей нуклеотидов вирусного генома. Мутации происходят у всех вирусов, независимо от того, является ли их генетическим аппаратом ДНК или РНК. В результате мутаций отдельные вирионы могут приобретать новые свойства. Дальнейшая судьба таких вирусов зависит от естественного отбора, сохраняющего популяцию, наиболее приспособленную к условиям существования.
Мутации могут иметь разные последствия. В одних случаях они ведут к изменению фенотипических проявлений в нормальных условиях. Например, увеличивается или уменьшается размер бляшек под агаровым покрытием; увеличивается или ослабляется нейровирулентность для определенного вида животных; вирус становится более чувствительным к действию химиотерапевтического агента и т. п.
В других случаях мутация является летальной, так как вследствие ее нарушается синтез или функция жизненно важного вирусспецифического белка, например вирусной полимеразы.
В некоторых случаях мутации являются условно летальными, так как вирусспецифический белок сохраняет свои функции в определенных, оптимальных для него, условиях и теряет эту способность в неразрешающих (непермиссивных) условиях. Типичным примером таких мутаций являются температурно-чувствительные (temperature sensitive) — ts-мутации, при которых вирус теряет способность размножения при повышенных температурах (39—42° С), сохраняя эту способность при обычных температурах выращивания (36—37° С).
По своему механизму мутации могут быть тоже разными. В одних случаях происходит делеция, т. е. выпадение одного или нескольких нуклеотидов, в других случаях происходит встраивание одного или нескольких нуклеотидов, а в некоторых случаях — замена одного нуклеотида другим.
Мутации могут быть прямыми и обратными. Прямые мутации меняют фенотип, а обратные мутации — реверсии — его восстанавливают. Возможны истинные реверсии, когда обратная мутация происходит в месте первичного повреждения, и псевдореверсии, если мутация происходит в другом участке дефектного гена (интрагенная супрессия) или в другом гене (экстрагенная супрессия). Реверсия не является редким событием, так как ревертанты обычно более приспособлены к данной клеточной системе. Поэтому при получении мутантов с заданными свойствами, например вакцинных штаммов, приходится считаться с возможной их реверсией к дикому типу.
Мутации носят случайный характер и объясняются статистическими законами.
В качестве физических мутагенов наиболее часто применяется ультрафиолетовое облучение, так как его энергия сопоставима с энергией химических связей. Реже применяются более жесткие виды облучения — рентгеновское и γ-облучение, а также обработка вирусных суспензий нейтронами, протонами, электронами и ядрами гелия, так как они вызывают сильные разрушения вирусных геномов и их инактивацию.
В качестве химических мутагенов применяют аналоги оснований (бромурацил, бромдезоксиуридин, 2-аминопурин, нитрозогуанидин и пр.), алкилирующие и флуоресцирующие соединения (профлавин), интеркалирующие агенты (актиномицин, этидий бромид), азотистую кислоту, гидроксиламин и многие другие.
ГЕНЕТИЧЕСКИЕ И НЕГЕНЕТИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ МЕЖДУ ВИРУСАМИ
Как в естественных, так и в экспериментальных условиях одна клетка может быть заражена не одним, а несколькими вирусами. В процессе такой смешанной инфекции могут иметь место различные формы взаимодействия как между вирусными геномами, так и между продуктами генов. При взаимодействии геномов могут наблюдаться такие формы генетических взаимодействий, как множественная реактивация, рекомбинация, пересортировка генов, кросс-реактивация, гетерозиготность. При взаимодействии на уровне продуктов генов могут иметь место негенетические взаимодействия: комплементация, интерференция, фенотипическое смешивание и др.
Множественная реактивация. Вирусная инфекция может возникнуть при заражении клетки несколькими ви-рионами с поврежденными геномами вследствие того, что функцию поврежденного гена может выполнять вирус, у которого этот ген не поврежден. Этот феномен был вначале обнаружен на бактериофагах и получил название множественной реактивации. В основе множественной реактивации лежит кооперативный процесс, при котором вирионы с поражением разных генов дополняют друг друга путем генетической рекомбинации, в результате чего репродуцируется исходный неповрежденный вирус. Эффективность множественности реактивации зависит от многих причин: степени повреждения генома вирионов, числа проникших в клетку вирионов, концентраций их в определенных участках клетки, аутоинтерференции поврежденных вирионов. Для множественной реактивации важное значение имеет расстояние между вирионами с поврежденными геномами внутри клетки. Обработка вирионов двухвалентными ионами металлов, ведущая к их агрегации, усиливает множественную реактивацию.
Рекомбинация. Генетической рекомбинацией называют обмен генетическим материалом, происходящий между родительскими вирусами. Возможен обмен полными генами (межгенная рекомбинация), так и участками одного и того же гена (внутригенная рекомбинация). Образующийся вирус-рекомбинант обладает свойствами, унаследованными от разных родителей.
Обычно рекомбинируемые штаммы обладают характерными признаками, которые обозначаются как маркеры. Например, были получены рекомбинанты между вирусами полиомиелита, обладающие повышенной устойчивостью и повышенной чувствительностью к гуанидину, разной нейровирулентностью, разной устойчивостью к повышенной температуре, разной чувствительностью к ингибиторам сывороток лошадей и коров и т. п. Для получения рекомбинантов используют штаммы, содержащие два или большее число маркеров.
Тест рекомбинации применяют для генетических исследований вирусов. С его помощью возможно построение генетических карт вирусов, в которых определяется, в каких участках генома произошли мутации, а также в условных единицах измеряется расстояние между разными мутациями.
Пересортировка генов. Вариантом рекомбинации является феномен, получивший название пересортировки генов. Она наблюдается при генетических взаимодействиях между вирусами, имеющими сегментированный геном. Образующиеся при этом гибридные формы вирусов называют реассортантами. Реассортанты вирусов гриппа получают при совместном культивировании вирусов с разными генами гемагглютинина и нейраминидазы. В этом случае из общего потомства путем нейтрализации соответствующих антигенов можно выделить интересующие исследователя варианты.
Существуют определенные группировки (констелляции или созвездия) генов, которые в данной системе клеток более стойки и делают вирус более жизнеспособным.
Сходные процессы пересортировки генов имеют место у вирусов гриппа типов А, В и С и у других вирусов с фрагментарным геном — у буньявирусов, аренавирусов (однонитчатые РНК) и реовирусов (ротавирусов) (двунитчатая РНК). Однако эти процессы не столь интенсивны и доступны изучению, как у вирусов гриппа.
Гетерозиготность. При совместном культивировании двух штаммов вируса может происходить формирование вирионов, содержащих в своем составе два разных генома или по крайней мере один полный геном и часть второго генома. Это явление названо гетерозиготностью.
Комплементация. Комплементация (дополнение) является таким видом негенетического взаимодействия при смешанной инфекции двумя вирусами, которое стимулирует репродукцию обоих партнеров или одного из них, но не изменяет генотипы вирусов. Принцип комплементации заключается в том, что вирус снабжает партнера недостающими компонентами, обычно белками, структурными или неструктурными.
Комплементация широко распространена среди вирусов и встречается как между родственными, так и неродственными вирусами. Феномен тесно связан с проблемой дефектности вирусов.
Поскольку в вирусной популяции помимо стандартных обычно присутствуют дефектные неинфекционные вирусные особи, в частности дефектные частицы, утратившие часть генетического материала, комплементация имеет место в инфекционном цикле многих вирусов и заключается в том, что члены популяции снабжают друг друга продуктами генов, которые дефектны у партнеров. Отличие комплементации от генетической рекомбинации заключается в отсутствии обмена генетическим материалом.
Комплементация встречается и у неродственных вирусов, принадлежащих к разным семействам. Одним из семейств, вирусы которого наиболее часто участвуют в комплементации, является семейство аденовирусов. В одних системах аденовирусы могут действовать как дефектные вирусы, в других — как помощники. Например, в культуре клеток почек макак резусов аденовирусы могут репродуцироваться только в присутствии SV40, который является в данном случае вирусом-помощником. В других системах сами аденовирусы действуют как вирусы-помощники, а вирусом-сателлитом является аденоассоци-ированный вирус, относящийся к семейству парвовирусов. Репродукция этого вируса полностью зависит от комплементирующего действия аденовирусов. Вирус гепатита В является помощником для дельта-агента, который покрывается его наружным белком — HBs-антигеном. Сочетание обоих вирусов обнаружено при наиболее тяжелых формах гепатита.
Возможна не только межцистронная, но и внутрицистронная комплементация в том случае, когда один ген кодирует несколько белков.
Фенотипическое смешивание. При совместном культивировании двух вирусов может наблюдаться феномен фенотипического смешивания, когда геном одного вируса бывает заключен в капсид, состоящий частично или полностью из белков другого вируса.
Фенотипическое смешивание наблюдается при смешанной инфекции,многими вирусами, причем эти вирусы могут быть как близкими друг другу (например, вирусы гриппа А и В или разные серологические подтипы вируса гриппа А), так и весьма далекими (онковирусы и рабдовирусы).
РЕСТРИКТАЗЫ И ФИЗИЧЕСКИЕ КАРТЫ ВИРУСОВ
Подлинную революцию в физическом картировании геномов вирусов произвело применение рестриктаз и секвенирование вирусных геномов. Рестриктазы имеют исключительное значение в молекулярной генетике вообще и генетической инженерии в частности. Их открытие (1968—1970 гг.) впервые дало возможность специфически расщеплять ДНК на строго определенные фрагменты, доступные для препаративного выделения и анализа.
Рестриктазы или эндодезоксирибонуклеазы — это просто организованные белки, являющиеся ферментами, широко распространенными среди прокариотов и участвующими в генетических процессах. В отличие от экзонук-леаз, отщепляющих концевые нуклеотиды или свободные остатки фосфорной кислоты, эндонуклеазы расщепляют молекулу ДНК изнутри, обычно — в местах, где преобладают пиримидиновые основания. Рестриктазы характеризуются высоковыраженной специфичностью, распознавая строго определенные последовательности нуклеотидов в двунитчатой ДНК.
Число новых рестриктаз стремительно нарастает и со временем, по-видимому, будут обнаружены рестриктазы, узнающие любую последовательность нуклеотидов.
Использование разных рестриктаз позволяет получать фрагменты разной величины, которые затем разделяются и анализируются путем электрофореза в агарозных или полиакриламидных гелях. Сочетание рестрикционного анализа с другими методами позволяет составить физические карты геномов вирусов. Физические карты вирусных геномов обозначают взаимное расположение генов, их границы, локализацию начала репликации, промоторов, лидеров, экзонов и интронов, сигнальных последовательностей и других генетических элементов.
Генетический код для синтеза белков вируса SV-40 записан не на одной, а на обеих нитях ДНК, а транскрипция разных генов идет в разных направлениях.
В настоящее время полностью расшифрованы нуклеотидные последовательности отдельных генов и целых геномов методом секвенирования (от англ. sequence — последовательность). Если речь идет о РНК-содержащих вирусах, то предварительным условием для дальнейшего их анализа является переписка РНК на ДНК с помощью РНК-зависимой ДНК-полимеразы (обратной транскриптазы), после чего генетический материал может быть подвергнут рестрикционному анализу.
Проектное задание к модулю
В качестве проектного задания студентам предлагается написание рефератов по следующим темам:
1. Природа дефектных вирусных геномов. Вирусы-сателлиты.
2. Вирусная интерференция
3. Необычные свойства ретровирусов
4. Трансформация клетки опухолеродными ДНК-вирусами
5. Классификация и основные свойства вирусов гриппа
6. Индукция специфического иммунного ответа на вирусы
7. Особенности репродукции пикорнавирусов
8. Семейство тогавирусов. Особенность репродукции и инфекционного процесса.
9. Вирус клещевого энцефалита.
10. Вирус бешенства
11. Вирусная персистенция
12. Основные свойства парамиксовирусов
13. Вирус кори. Биология возбудителя. Особенности патогенеза
14. Аденовирусы, их репликация и связь с другими вирусами
16. Пути распространения вирусных болезней растений.
17. Прионы. Возбудители или провокаторы или….
18. Вироиды как вирусоподобные инфекционные агенты.
19. Вирусы, вызывающие респираторные инфекции. Сравнительная характеристика
20. Место вирусов в биосфере.
21. Бактериофаги. Их морфологическое многообразие и взаимодействие с бактериями.
22. Атипичная пневмония и возможность происхождения новых вирусов
23. Роль вирусов в возникновении злокачественных опухолей
24. Особенности транскрипции РНК- содержащих вирусов.
25. Морфогенез вирусов или морфологические превращения в процессе репродукции.
Тест рубежного контроля
Читайте также: