
Днк эукариот прокариот и вирусов
Презентация была опубликована 6 лет назад пользователемВера Бабина
Презентация на тему: " Лекция 9. Организация геномов вирусов, прокариот, эукариот Мяндина Галина Ивановна д.б.н., профессор." — Транскрипт:
1 Лекция 9. Организация геномов вирусов, прокариот, эукариот Мяндина Галина Ивановна д.б.н., профессор
3 Понятия генома и ДНК тождественны Геном - это совокупность всех молекул ДНК в клетке организма данного вида Геном человека состоит из 3.1 x 10 9 пар нуклеотидов (п.н.) ДНК
4 Геномы прокариот и эукариот Хромосомная ДНК содержится: Прокариоты - в нуклеоиде Эукариоты - в ядре Внехромосомная ДНК: Прокариоты – плазмиды, Эукариоты - ДНК митохондрий (мтДНК) и хлоропластов, кольцевые ДНК в нуклеоплазме (эписомы) В хромосомах эукариот содержится 95% всей ДНК, в митохондриях – 5%
5 Геном – это генетическая характеристика вида геном Homo sapiens включает: 24 ДНК хромосом (22 аутосомы, Х- и Y- хромосомы) одна ДНК митохондрий (мтДНК)
6 Содержание ДНК в клетках организмов разных видов (С-парадокс) Название видап.н./геном E. coli4.5 x 10 6 Homo sapiens3.1 x 10 9 Drosophila1.7 x 10 8 Maize2.0 x 10 9 Arabidopsis7.0 x 10 7 Lily1.0 x 10 11
7 Принцип организации генетического материала в клетке Существует несоответствие между длиной молекулы ДНК и размером клетки Поэтому ДНК может существовать в виде компактной структуры в строго ограниченном участке клетки Суммарная длина молекул ДНК в клетке человека -1,8 м, диаметр ядра – 5 мкм
8 Хромосома прокариот - кольцевая суперспирализованная двухцепочечная ДНК
9 Упрощенная генетическая карта хромосомы E.coli Расстояние между генами прокариот измеряют в минутах (цифры внутри кольца) гены (опероны) обозначены символами
10 Плазмиды – внехромосомные ДНК бактерий Электронная микрофотография открытых кольцевых молекул ДНК плазмид Кольцевые молекулы ДНК Являются репликонами и могут существовать самостоятельно Конъюгативные плазмиды содержат гены переноса и обеспечивают процесс конъюгации у бактерий Плазмиды содержат гены, обеспечивающие дополнительные свойства бактериальной клетки (например, устойчивость к антибиотикам)
11 Степень конденсации (плотность упаковки) хромосом = размер молекулы ДНК длина хромосомы Самая короткая хромосома человека: 4,6Х10 7 п.н. ДНК ( мкм), В митозе ее размер составляет 2 мкм Степень конденсации = 14000/2 = 7000
12 Нуклеосома – элементарная единица структуры хроматина эукариот
13 Уровни организации хроматина эукариот
14 Уровни организации хроматина метафазная хромосома – 1400 нм участок метафазной хромосомы – 700 нм; Нуклеомеры- субъединицы толстой фибриллы нм хроматиновая фибрилла – 30 нм; нуклеосомы – 11 нм двойная спираль ДНК – 2 нм
15 Хромосомная ДНК эукариот состоит из участков Уникальные последовательности представлены в одной-двух копиях на геном (45%) Повторяющиеся последовательности (повторы) -55%: различают высоко повторояющиеся и умеренно повторяющиеся последовательности ДНК Кодирующие последовательности составляют 1,5% - 2% генома человека
16 Инвертированные повторы (палиндромы) Одинаковые последовательности ДНК, взаимно противоположной ориентации в цепях ДНК А А Т Т Г Т А Ц А А Т Т Т Т А А Ц А Т Г Т Т А А
17 Тандемные и диспергированные повторы Тандемные повторы расположены блоками (в тандеме) во многих участках хромосом эукариот, например: ТТАГГГ… ТТАГГГ ТТАГГГ//ТТАГГГ Диспергированные повторы не объединены в блоки и рассеяны по всему геному В зависимости от размера повторенного фрагмента различают: сателлиты, минисателлиты и микросателлиты
18 Повторы ТандемныеДиспергированные сателлиты LINE (повторяющаяся единица имеет длину п.н) минисателлиты SINE (повторяющаяся единица – 50 – 300 п.н.) микросателлиты
19 При изучении генома человека мы узнали: геном человека содержит 3 млрд.164 млн. 700 т.п.н. общее число генов составляет – Функции 50% генов неизвестны Наибольшее количество генов содержит хромосома 1 (2968 генов), наименьшее – Y- хромосома (231 ген). наши ДНК совпадают на 99,9%, т.е. индивидуальные отличия составляют 0,1% генома человека 1,5% генома человека кодируют белки повторяющиеся последовательности занимают 50% генома человека
20 последовательности ДНК человека Геном человека Экзоны 1,5% Интроны, промоторы, энхансеры, сайленсеры 24% Уникальные некодирующие последовательности 15% Повторы, содержащие МГЭ 44% Повторы, не содержащие МГЭ 15%
21 Внехромосомная ДНК эукариот содержится в митохондриях и пластидах электронная микрофотография мтДНК мыши
22 Митохондриальная ДНК человека содержит 37 структурных генов
23 Характеристика мтДНК наследуется по материнской линии гены мтДНК не имеют интронов нет системы репарации нет гистонов генетический код отличается от ядерного большую часть белков митохондрий кодируют хромосомные гены мутации генов митохондрий – причина митохондриальных болезней человека
24 Мобильные генетические элементы (МГЭ) это последовательности ДНК с непостоянной локализацией Транспозиция МГЭ обеспечивается особенностями их структуры и наличием белков- ферментов (транспозазы и резольвазы)
25 Свойства МГЭ Содержат гены транспозиции и осуществляют перенос генов Не являются репликонами не могут существовать самостоятельно Являются биологическими мутагенами Изменяют активность генов Обеспечивают рекомбинацию хромосом и эволюцию геномов
26 Мобильные элементы генома Прокариоты IS элементы и транспозоны Эукариоты Транспозоны и ретротранспозоны
27 механизмы транспозиции МГЭ Cut – paste: вырезание МГЭ и его встраивание в другой участок ДНК Copy – paste: образование копии МГЭ (репликация) и ее перемещение в другой участок ДНК, увеличение числа копий МГЭ
28 Мобильные элементы генома прокариот IS– элементы – (insertion sequences) - простые инсерционные последовательности размером от 800 до 1000 п.н., содержат ген транспозазы и концевые IR-элементы (инвертированные повторы) Транспозоны (Tn) – сложные МГЭ, размером более 2000 п.н., содержат гены транспозазы и резистентности к антибиотикам
30 Мобильные элементы генома эукариот транспозоны - редко встречаются в геноме эукариот. ретротранспозоны - транспозиция происходит при образовании РНК - копии с последующим синтезом копии ДНК с помощью фермента обратной транскриптазы (RT). Большая часть мобильных элементов генома эукариот – это ретротранспозоны
31 Мобильные элементы генома человека Ретротранспозоны - к ним относятся последовательности SINE (Alu) и LINE (L1) Эндогенные ретровирусы (HERV) транспозоны (редко, например, транспозон mariner)
32 Геномы вирусов могут быть представлены разными формами молекул ДНК или РНК: Однонитевая кольцевая ДНК (фаг М13) Однонитевая линейная ДНК (парвовирусы) Двунитевая кольцевая ДНК (фаг λ, вирусы папилломы человека) Двунитевая линейная ДНК (аденовирусы, герпесвирусы) Однонитевая РНК (ретровирусы, вирус полиомиелита) Двунитевая РНК (реовирусы)
Прокариоты
Все живые организмы, имеющие клеточное строение, делятся на две большие группы: прокариоты и эукариоты (Рис. 1).

Рис. 1. Разделение живых организмов (Источник)
Древнейшие на Земле организмы, не имеющие клеточного ядра, появившиеся около четырех миллиардов лет тому назад, называются прокариотами, то есть доядерными. В настоящее время они тоже распространены, обитают в воде, почве, воздухе, на покровах животных и растений, а также внутри них. Прокариоты освоили экстремальные места обитания (Рис. 2): горячие источники (они выживают и живут при температуре 70 0 и выше), моря и соленые озера (галобактерии живут при солености около 30 %).

Рис. 2. Места обитания прокариот (Источник)
Форма бактерий чрезвычайно разнообразна: шаровидная, палочковидная и изогнутая (Рис. 3).

Рис. 3. Формы бактерий (Источник)
Размеры клеток большинства прокариот – от 0,2 до 10 микрометров, встречаются и карлики (нанобактерии и микоплазмы), размер которых – от 0,05 до 0,1 микрометра. Кроме этого, существуют и гиганты (макромонусы) с размерами до 10 микрометров. Средний размер клетки бактерии – около 1 микрометра. Размеры прокариот меньше размеров эукариот.
Строение прокариотической клетки
По сравнению с эукариотической, клетка прокариот выглядит гораздо проще (Рис. 4).

Рис. 4. Клетка прокариот и эукариот (Источник)
У прокариот нет ядра, единственная кольцевая молекула ДНК, находящаяся в клетках прокариот и условно называемая бактериальной хромосомой, находится в центре клетки, однако эта молекула ДНК не имеет оболочки и располагается непосредственно в цитоплазме.
Рассмотрим строение прокариотической клетки (Рис. 5).

Рис. 5. Строение прокариотической клетки (Источник)
Снаружи клетки прокариот, так же как и эукариотические клетки, покрыты плазматической мембраной. Строение мембран у двух этих групп организмов одинаковое. Клеточная мембрана прокариот образует многочисленные впячивания внутрь клетки – мезосомы. На них располагаются ферменты, обеспечивающие реакции обмена веществ в прокариотической клетке. Поверх плазматической мембраны клетки прокариот покрыты оболочкой, состоящей из углеводов, напоминающей клеточную стенку растительных клеток. Однако эта стенка образована не клетчаткой, как у растений, а другими полисахаридами – пектином и муреином. В цитоплазме прокариотических клеток нет мембранных органоидов: митохондрий, пластидов, ЭПС, комплекса Гольджи, лизосом. Их функции выполняют складки и впячивания наружной мембраны – мезосомы. В цитоплазме прокариот беспорядочно располагаются мелкие рибосомы. Цитоскелета в прокариотических клетках тоже нет, но иногда встречаются жгутики, которые способствуют передвижению бактерий. На поверхности бактериальной клетки находятся пили – белковые нити, с помощью которых бактерии присоединяются к субстрату или поверхности. Половые пили служат для обмена генетического материала между различными бактериями.
Фотосинтезирующие бактерии – цианобактерии, имеют в клетках фотосинтезирующие мембраны или тилакоиды, в которых содержатся пигменты, участвующие в процессе фотосинтеза (Рис. 6), такие как хлорофилл.

Рис. 6. Цианобактерия (Источник)
На тилакоидах содержатся пигменты, являющиеся вспомогательными при процессе фотосинтеза – фикобилины: аллофикоцианин, фикоэритрин и фикоцианин. Фикобилины образуют прочные соединения с белками (фикобилинпротеиды). Связь между фикобилинами и белками разрушается только кислотой.
В клетках прокариот откладываются и запасные питательные вещества, отложение или запас происходит в результате избытка питательных веществ, а потребление при недостатке питательных веществ. К запасным питательным веществам относятся полисахариды (крахмал, гликоген, гранулеза), липиды (гранулы или капли жира), полифосфаты (источник фосфора и энергии).
Большинство эукариот являются аэробами, то есть используют в энергетическом обмене кислород воздуха. Напротив, многие прокариоты являются анаэробами, и кислород для них вреден. Некоторые бактерии, называемые азотфиксирующими, способны усваивать азот воздуха, чего эукариоты делать не могут. Те виды прокариот, которые получают энергию благодаря фотосинтезу, содержат особую разновидность хлорофилла, который может располагаться на мезосомах.
В неблагоприятных условиях (холод, жара, засуха) многие бактерии образуют споры. При спорообразовании вокруг бактериальной хромосомы образуется особая плотная оболочка, а остальное содержимое клетки отмирает. Спора может десятилетиями находиться в неактивном состоянии, а в благоприятных условиях из нее снова прорастает активная бактерия (Рис. 7).

Рис. 7. Схема образования спор у бактерий (Источник)
Размножение прокариот
Чаще всего прокариоты размножаются бесполым путем: ДНК удваивается, и далее клетка делится в поперечной плоскости пополам (Рис. 8). В благоприятных условиях бактерии способны делиться каждые 20 минут; при этом потомство от одной клетки через трое суток теоретически имело бы массу 7500 тонн! К счастью, таких условий в принципе быть не может.

Рис. 8. Размножение прокариот (Источник)
Половое размножение у прокариот наблюдается гораздо реже, чем бесполое, однако оно очень важно, так как при обмене генетической информацией бактерии передают друг другу устойчивость к неблагоприятным воздействиям (например, к лекарствам). При половом процессе бактерии могут обмениваться как участками бактериальной хромосомы, так и особыми маленькими кольцевыми двуцепочечными молекулами ДНК – плазмидами. Обмен может происходить через цитоплазматический мостик между двумя бактериями или с помощью вирусов, усваивающих участки ДНК одной бактерии и переносящих их в другие бактериальные клетки, которые они заражают.
Заключение
Мы рассмотрели прокариотическую клетку, которая организована достаточно просто по сравнению с эукариотической клеткой, основным отличием которой является отсутствие оформленного ядра, кольцевая молекула ДНК располагается в цитоплазме свободно и не окружена ядерной оболочкой. В прокариотической клетке нет мембранных органелл, которые свойственны эукариотическим клеткам.
Список литературы
- Беляев Д.К. Общая биология. Базовый уровень. – 11 издание, стереотипное. – М.: Просвещение, 2012.
- Пасечник В.В., Каменский А.А., Криксунов Е.А. Общая биология, 10-11 класс. – М.: Дрофа, 2005.
- Агафонова И.Б., Захарова Е.Т., Сивоглазов В.И. Биология 10-11 класс. Общая биология. Базовый уровень. – 6-е изд., доп. – Дрофа, 2010.
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
Домашнее задание
- Каково основное различие между прокариотическими и эукариотическими клетками?
- Что такое бактериальная хромосома?
- Как происходит половое размножение прокариот?
Если вы нашли ошибку или неработающую ссылку, пожалуйста, сообщите нам – сделайте свой вклад в развитие проекта. 

ОРГАНИЗАЦИЯ ГЕНОМА ПРОКАРИОТ
Геном прокариот может состоять из одной или нескольких крупных молекул ДНК, называемых хромосомами, и небольших молекул ДНК – плазмид. В хромосомах представлены практически все гены, необходимые для жизнедеятельности бактерии. Плазмиды же несут гены, необязательные для бактерии, без них клетка может обойтись, хотя в некоторых условиях они способствуют ее
Хромосомы и плазмиды могут представлять собой как кольцевые, так и линейные двухцепочечные молекулы ДНК. Геном бактерий может состоять из одной или нескольких хромосом и плазмид . Хромосома(ы) в бактериальной клетке пред-ставлена(ы) в виде одной копии, т.е. бактерии гаплоидны. Плазмиды же могут присутствовать в клетке как в виде одной копии, так и в нескольких.
Хромосома уложена в компактную структуру – нуклеоид, который имеет овальную или сходную с ней форму. Его структура поддерживается ДНК-связывающими гистоноподобными белками и молекулами РНК. С нуклеоидом также ассоциированы молекулы РНК-полимеразы и ДНК-топоизомеразы I. По периферии нуклеоида располагаются петли хромосомной ДНК, которые находятся в транскрипции в активном состоянии. При подавлении транскрипции эти петли втягиваются внутрь. Нуклеоид не является стабильным образованием и во время различных фаз роста бактериальных клеток изменяет свою форму. Изменение его пространстве ой организации сопряжено с изменением транскрипционной активностью определенных генов бактерий.
В состав хромосомы могут входить геномы умеренных фагов. Включение их геномов в клеточный может происходить после заражения фагами бактерий. При этом одни фаговые геномы интегрируют в строго определенные участки хромосомы, другие – в участки различной локализации.
Размер геномов прокариот колеблется от нескольких сотен тысяч до десятка миллионов пар нуклетидов.
Геномы прокариот отличаются друг от друга по содержание ГЦ-пар, их доля в их составе колеблется от 23 до 72 %. Интересно, что в ДНК бактерий, обитающих при высоких температурах, содержание этих нуклеотидов повышено. Их преобладание над АТ-парами обуславливает более высокую температуру плавления ДНК, что является жизненно необходимым фактором для таких бактерий. Нужно
отметить, что в белках термофильных бактерий повышено также и содержание полярных аминокислот, что делает их более устойчивыми к денатурации при повышенных температурах. В составе белков хеликобактерий (обитающих в кислой среде) больше аминокислотных остатков аргинина и лизина. Остатки этих аминокислот способны связывать ионы водорода, тем самым, оказывая влияние на кислотность среды, и способствуя выживанию бактерий в сложных экологических условиях.
О числе генов в геноме судят по наличию в их составе открытых рамок считывания (ОРС). ОРС представляет собой полинуклеотидную последовательность, потенциально способную кодировать полипептид. О существовании ОРС на тех или иных участках ДНК судят на основании расшифрованной первичной структуры ДНК.
Основным критерием принадлежности участка полинуклеотидной цепи к ОРС служит отсутствие стоп-кодонов на достаточно протяженном участке после стартового кодона. В то же время наличие ОРС является недостаточным условием для утверждения о наличии на данном участке ДНК гена.
Гены, прокариот, как правило, имеют оперонную организацию. В одном опероне обычно представлены гены, ответственные за осуществление одного и того же метаболического процесса.
ОРГАНИЗАЦИЯ ГЕНОМА ЭУКАРИОТ
Хранителем генетической информации у эукариот так же как и у прокариот, является двухцепочечная молекула ДНК. Основная часть генетической информации у них сосредоточена в клеточном ядре в составе хромосом, значительно меньшая часть представлена в составе ДНК митохондрий, хлоропластов и других пластид.
Геномная ДНК эукариот представляет собой совокупность ДНК гаплоидного набора хромосом и внехромосомной ДНК, представленные в клетке зародышевой линии. Общее содержание ДНК, приходящиеся на один гаплоидный набор, носит название величина С. Ее выражают в пг ДНК, дальтонах или в парах нуклеотидов (1 пг = 6,1 • 10 11 Да = 0,965 • 10 п.н.). Значение величины С, как правило, возрастает с увеличением организации живых ор- ганизмов . Однако, у некоторых родственных видов величины С могут значительно отличаться, в то время как морфоло-гия и физиология этих видов отличаются друг от друга несущественно.
Каково же значение негенной ДНК? Существуют несколько гипотез, объясняющих ее роль. Одной из таковой является предположение, что некодирующие последовательности генома эукариот способствуют защите генов от химических мутагенов.
Ядерная ДНК эукариот состоит из уникальных и повторяющихся последовательностей. Повторяющаяся ДНК в свою очередь может быть разделена на две фракции: умеренно повторяющаяся и часто повторяющаяся ДНК:
К часто повторяющейся принадлежит ДНК, представленная в геноме более 105 копий. К этой фракции относится сателлитная ДНК.
Содержание сателлитной ДНК составляет в геноме эукариот от 5 до 50 % от всей ДНК. Эта ДНК преимуществе о обнаруживается в центромерных и теломерных рай- онах хромосом, где она выполняет структурные функции. Сателитная ДНК состоит из тандемных повторов длиной от 1 до 20 и более п.н. Благодаря простоте организации и многочисленным копиям эта ДНК обладает способностью к быстрой ренатурации.
В геноме эукариот различают микросателлиты, минисателлиты и макросателлиты. Микросателлиты образованы многократно повторяющимися мономерными звеньями (1 – 4 п.н) и имеют размер до нескольких сотен пар нуклеотидов . Они разбросаны по геному, их длина и общее количество копий коррелирует с размером генома. Количество копий микросателлитов в геноме может достигать десятков и сотен тысяч.
Макросателлиты обладают в сравнении с микросателлитами и минисателлитами большим размером повторяющегося звена до 1000 и более пар нуклеотидов. Они обнаружены в геномах птиц, кошек и человека.
Умеренно повторяющиеся последовательности в геноме представлены до 104 копий. К ним относятся генные семейства и МГЭ.Генные семейства образуют гены, обладающие гомологичной (или идентичной) нуклеотидной последовательностью, и выполняющие одну и ту же или сходные функции. Они могут быть организованы в виде кластеров или же разбросаны по геному. Существование генов в большом числе копий обеспечивает повышенное образование продуктов их экспрессии. Семейства образуют гены гистонов, рРНК, тРНК и др. МГЭ эукариот составляют в среднем около 10 – 30 % генома. Они могут концентрироваться в определенных участках хромосомы или быть рассеянными по геному
К уникальной ДНК относятся неповторяющиеся нуклеотидные последовательности. Ее содержание у различных видов варьирует от 15 до 98 %. Доля уникальных последовательностей ДНК у низших эукариот значительно выше, чем у высших. К уникальной ДНК относятся как кодирующие, так и не кодирующие последовательности. При этом большая часть уникальной ДНК не несет функции кодирования. К некодирующей уникальной ДНК относятся интроны, к кодирующей – экзоны.
Строение м-РНК
Информационная (матричная) РНК (иРНК, мРНК; messenger RNA, mRNA): однонитевая РНК, содержащая информацию об аминокислотных последовательностях белка.
мРНК содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах . На долю м-РНК приходится примерно 0,5-1% от общего содержания РНК клетки.
Зрелая мРНК эукариот наряду с основной последовательностью нуклеотидов, в которой закодирована информация о последовательности аминокислот в соответствующем белке, содержит целый ряд некодирующих последовательностей, необходимых для ее трансляции рибосомами. Часть этих последовательностей, такие как кэп-группа и 3'-концевая поли(А), не кодируются непосредственно генами, а добавляются ко- и посттранскрипционно, другие имеют генное происхождение. Эти последовательности часто содержат регуляторные сигналы, обеспечивающие определенный уровень трансляции мРНК рибосомами.
Участок мРНК, расположенный между кэп-группой и первым инициирующим кодоном основнойоткрытой рамки считывания (ОРС) , которая и несет информацию о последовательности аминокислот в белке, получил название 5'-концевой нетранслируемой области (5'UTR - 5' untranslated region), илилидерной последовательности . Сегмент мРНК, расположенный между последним терминирующим кодоном основной ОРС и началом поли(А)- последовательности , называют 3'-концевой нетранслируемой областью (трейлером) - (3'UTR) . Первое название не совсем удачно: последовательности 5'UTR, как правило, способны образовывать сложные вторичные структуры типа "стебель-петля" и содержать короткие ОРС (uORF - upstream open reading frame) , которые оказывают сильное влияние на эффективность трансляции мРНК.
Помимо этого, 5'UTR могут включать в себя регуляторные последовательности, распознаваемые транс-действующими белковыми факторами. Последовательности 5'UTR обеспечивают регулируемую трансляцию мРНК (и координированную экспрессию соответствующих генов) в онтогенезе многоклеточных организмов.
3'UTR и поли(А)-последовательность оказывают влияние на состояние рибосом после терминации синтеза полипептидных цепей. Кроме того, 3'-концевая поли(А)-последовательность участвует в инициации трансляции.
31)ПРОЦЕССИНГ РНК
Процессингу подвергаются различные виды РНК: иРНК, рРНК, тРНК и др. Наиболее ярко постранскрипционные модификации РНК выражены у эукариот. У прокариот процессинг РНК необходим при образовании зрелых молекул рРНК и тРНК.
ПРОЦЕССИНГ иРНК
В процессе созревания иРНК эукариот происходит образование на 5’-конце кэпа, удаление интронов, синтез на 3’-конце полиА-последовательности. В отличие от эукариот иРНК только в единичных случаях подвержены процессингу. Эукариоты Созревание иРНК происходит в ядре эукариотических клеток. Этот процесс начинается, как правило, уже в ходе транскрипции. К 5’-концу иРНК присоединяется кэп. На 3’-конце иРНК по окончании транскрипции образуется полиА-последовательность, сигналом для полиаденилирования служит последовательность AAUAAA, расположенная за 11 – 30 нуклеотидов до сайта полиаденилирования. иРНК эукариот также подвергаются сплайсингу. Зрелая иРНК транс-портируется из ядра в цитоплазму клетки, где она служит матрицей для синтеза белка Рис.5.18. Процессинг иРНК эукариот
Следует отметить, что некоторые иРНК экариот не имеют полиА-последовательностей, и что некоторые предшественники иРНК не содержат интронов. Естественно, что они неподвергаются сплайсингу. Процессинг иРНК прокариот иРНК прокариот, как правило, процессингу не подвергается, потому что процессы транскрипции итрансляции у них сопряжены. Еще до завершения транскрипции с иРНК, синтезируемой РНКполимеразой, взаимодействуют рибосомы, которые и начинают син-тез полипептидных цепей Некоторые полицистронные иРНК прокариот могут расщеп-ляться с образованием индивидуальных иРНК. В одних случаях такое расщепление необходимо для успешной трансляции, в других не является обязательным. В некоторых случаях 3’-конец иРНК прокариот подвергается посттранскрипционному полиаденилированию, размер поли А-последовательностей составляет 14 – 60 нуклеотидов. Некоторые пре-иРНК прокариот содержат интроны.
ПРОЦЕССИНГ тРНК
Почти все тРНК синтезируются в виде предшественников – более длинных молекул (пре-тРНК). В результате процессинга происходит удаление нуклеотидных последовательностей с флангов пре-тРНК. С 5’-конца фрагмент нуклеотидной цепи отщепляет фермент, называемой РНК-азой. РНКазой являетсярибонуклеопротеином,каталитическую функцию в котором осуществляет РНК-компонент, белок же выполняет структурную роль. В бактериальной РНКазе есть участок, комплементарный ЦЦА участку тРНК. Эукариотическая РНКаза P узнает другие элементы предшественника тРНК. С 3’-конца пре-тРНК действует экзонуклеаза, укорачивающая РНК постепенно, удаляя по одному нуклеотиду. На заключительных ста-диях созревания тРНК к 3’-концу полинуклеотидилтрансфераза присоединяет последовательность ЦЦА (рис. 5.20). Общая схема процессинга тРНК. У прокариот ЦЦA-последовательность может быть закодиро-вана в генах тРНК, тогда в созревании пре-тРНК, считанных с таких генов, полинуклеотидилтрансфераза может не участвовать. Однако в некоторых случаях ЦЦА-последовательность может быть удалена экзонуклеазой в процессе созревания тРНК, тогда для ее восстановления необходимо участие нуклеотидилтрансферазы. У эукариот ЦЦA-последовательность не кодируется в генах тРНК, она добавляется посттранскрипционно. У прокариот первичный транскрипт может содержать несколько последовательностей тРНК, их процессинг включает вырезание
индивидуальных молекул тРНК. В процессе созревания тРНК также происходит модификация азотистых оснований – в результате которой образуются минорные основания: псевдоуридин, дигидроуридин, тимидин, 7-метил-гуанозин, инозин и др. Сплайсинг пре-тРНК В процессе сплайсинга нуклеаза вырезает интрон, а лигаза обеспечивает сшивание двух фрагментов тРНК за счет образования фосфодиэфирной связи, в результате обра-зуется ковалентно замкнутая молекула тРНК (рис. 5.22). ПРОЦЕССИНГ рРНК Поцессинг необходим для созревания как прокариотических рРНК, так и эукариотических.
Образование рРНК у прокариот происходит в результате процессинга первичного транскрипта (30 S), содержащего в своем соста-ве нуклеотидные последовательности всех трех 16S, 23S и 5S рРНК, а также тРНК. При созревании из первичного транскрипта происходит вычленение индивидуальных рРНК и тРНК
Большие рРНК млекопитающих 18S, 5,8S и 28S синтезируются в составе единого первичного транскрипта, обозначаемого 45S-РНК. 45S-РНК во время транскрипции метилируется. Метилированию подвергаются главным образом остатки рибозы. Из 45S-РНК вырезаются индивидуальные рРНК, которые затем при участии 5S рРНК формируют рибосомы (рис. 5.24). Рис. 5.24. Процессинг рРНК млекопитающих У некоторых примитивных эукариот в составе первичного транскрипта нуклеотидные последовательности, соответствующие 26S РНК, содержат интрон. В результате процессинга из первичный транскрипта не только вырезаются индивидуальные рРНК, но и уда-ляется интрон посредством аутосплайсинга (рис. 5.25). Существование различных видов РНК в клетках живых су-ществ обуславливает протекание важнейших биологических процес-сов, связанных, прежде всего, с реализацией генетической информа-ции закодированной в ДНК. Для приобретения функциональной ак-тивности РНК в большинстве случаев должна подвергнуться слож-нейшим посттранскрипционным модификациям. Только иРНК прокариот для проявления своих биологических свойств, как правило, не нуждается в процессинге.

Несмотря на существование очевидных преград и трудностей, которые подчас встают на пути развития и внедрения продуктов генной инженерии (ГИ), XXI век уже невозможно представить без плодов этой важной и многообразной технологии в арсенале современного биолога. Наиболее часто используемым организмом в ГИ являются бактерии.
Что такое ГИ и зачем она нам нужна? Почему бактерии так популярны у генных инженеров? В каком виде проще всего внести нужный ген в бактерию? С какими трудностями можно столкнуться, работая с этими организмами? Что произошло раньше: создание первой генноинженерной бактерии или открытие структуры ДНК и генома? Об этом и многом другом читайте под катом.
В данном пункте приведено краткое описание так называемой Центральной догмы молекулярной биологии. Если вы обладаете базовыми знаниями в молекулярной биологии, то смело переходите к пункту 1.
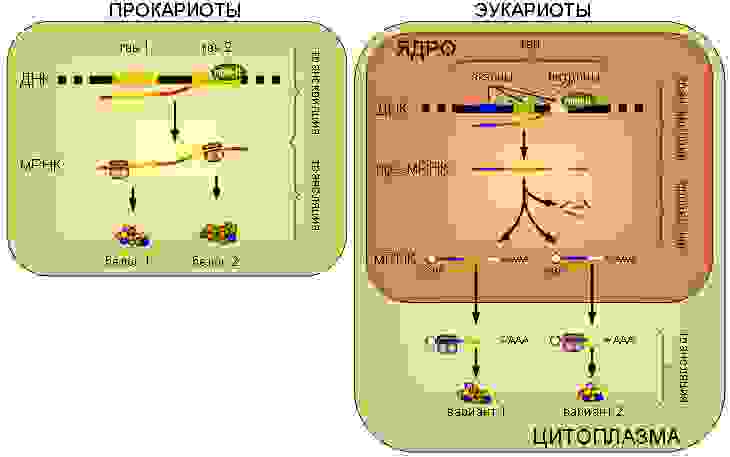
Центральная догма молекулярной биологии в одной картинке
Итак, начнём. Вся информация о всех стадиях развития и свойствах любого организма, будь то прокариоты (бактерии), археи или эукариоты (все остальные одно- и многоклеточные), закодирована в геномной ДНК, которая представляет собой комплекс двух комплементарных друг другу полинуклиотидных цепей, образующих двойную спираль (комплементарные нуклеотиды ДНК: A-T и G-C). Хромосомы эукариот представляют собой линейные двухцепочечные молекулы ДНК, а хромосомы прокариот закольцованы. Зачастую гены составляют лишь небольшую часть всего генома (у человека — около 1,5%).
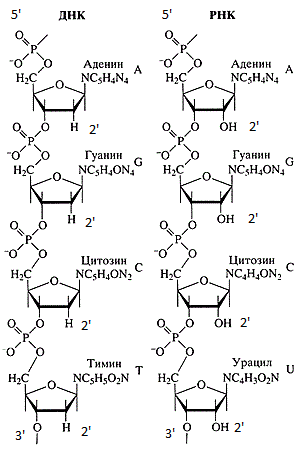
Цепочки ДНК и РНК.
Последовательности ДНК и РНК всегда записывают от 5'-конца к 3'-концу. На то есть ряд причин:
- Синтез новых цепочек ДНК и РНК начинается с 5'-конца (ДНК-полимеразы (ферменты, синтезирующие комплементарную цепь ДНК на матрице ДНК или РНК) и РНК-полимеразы (ферменты, синтезирующие комплементарную цепь РНК на матрице ДНК или РНК) идут по матрице в направлении 3' -> 5', стало быть новая цепь синтезируется в направлении 5' -> 3');
- Рибосома читает кодоны, передвигаясь по мРНК в направлении 5' -> 3';
- Последовательность аминокислот записана в кодирующей цепи ДНК в направлении 5' -> 3' (значащая часть мРНК представляет собой точную копию участка кодирующей цепи ДНК с заменой тимина на урацил и с гидроксильной группой (-OH) вместо водорода в положении 2', разумеется);
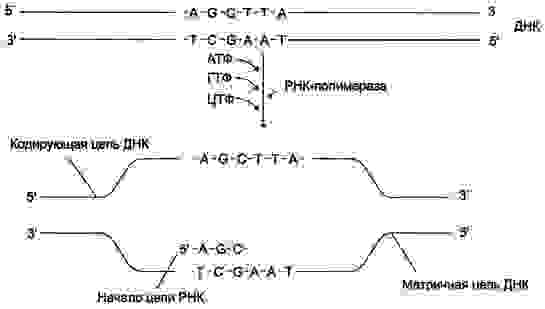
Ген — это участок геномной ДНК, задающий последовательность нуклеотидов молекулы РНК:
Рибосомные РНК — незаменимые компоненты рибосомы. Основной функцией рРНК является обеспечение процесса трансляции: она участвует в считывании информации с мРНК при помощи адапторных молекул тРНК и катализе образования пептидных связей между присоединёнными к тРНК аминокислотами и растущей цепью белка.
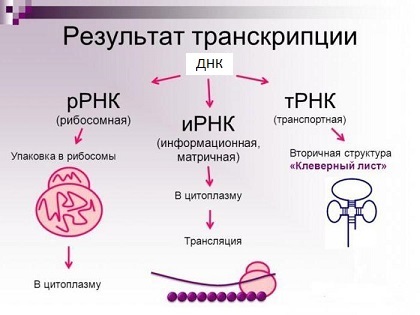
Основные типы молекул РНК (на самом деле их куда больше).
В случае с генами, кодирующими белок процесс расшифровки генетической информации выглядит так:

Прокариотический транскприпционный комплекс. Указанные на рисунке буквы являются общепринятыми обозначениями соответствующих субъединиц. σ70 — сигма фактор генов домашнего хозяйства E. coli
Бактерия в генной инженерии — это потенциальный исходный материал для создания:
Интересным является тот факт, что первые успешные опыты в области генной инженерии бактерий произошли задолго до эпохальной работы Уотсона и Крика. Более того, на основании этих опытов был доказан сам факт того, что информация содержится именно в ДНК, после чего учёные могли не тратить своё время на гипотезы об РНК и белке.
Таким образом, было доказано, что носителем информации о признаках является именно ДНК. Кроме того, было наглядно показано, что возможно самопроизвольное проникновение инородной молекулы ДНК в бактериальную клетку.
Почему бактерии так популярны при очевидных недостатках (например, отсутствие эукариотических посттрансляционных модификаций)? Всё просто. Они неприхотливы в работе, просты в использовании и не требуют дорогих питательных сред.
Современная генная инженерия бактерий в основном представляет собой внедрение плазмидного вектора (модифицированной бактериальной плазмиды, содержащей целевой ген и набор других необходимых элементов, о которых речь пойдёт ниже). Изменение хромосомы бактерии менее типично, но эта процедура также не является чем-то диковинным: например, ген РНК-полимеразы бактериофага Т7 был введёт в хромосому кишечной палочки с помощью вектора на основе профага λ в процессе создания одного из популярных в лабораторной плактике штаммов. Причин, по которым зачастую исследователь выберет внедрение гена в составе плазмидного вектора три:
Кроме гена и промотора основными элементами плазмидного вектора являются:
-
ori — область начала репликации плазмиды. Нужна для поддержания постоянного количества плазмиды и её наследования дочерними клетками;
Другим примером может служить, использование гена β-глюкуронидазы (GUS). Данный фермент превращает определённые соединения в окрашенные или флуоресцирующие, что может быть обнаружено визуально по окрашенности колонии. Естественно, данные соединения нужно добавлять в питательную селективную среду. Ещё один пример — использование гена зелёного флуоресцентного белка (GFP) (хотя использование GUS и GFP более типично для работ с клетками растений и животных);
участок, отвечающий за контроль копийности (мало плазмид в клетке — плохо, много — тоже плохо);
Что ж, вектор у нас на руках. Как в него встроить ген? И вообще, где этот ген взять?
Допустим, что мы знаем последовательность нуклеотидов нужного нам гена. Тогда поступают следующим образом:
- Химически синтезировать короткие одноцепочечные полинуклеотиды таким образом, чтобы они частично перекрывались;
- Объединить эти фрагменты с помощью ПЦР.
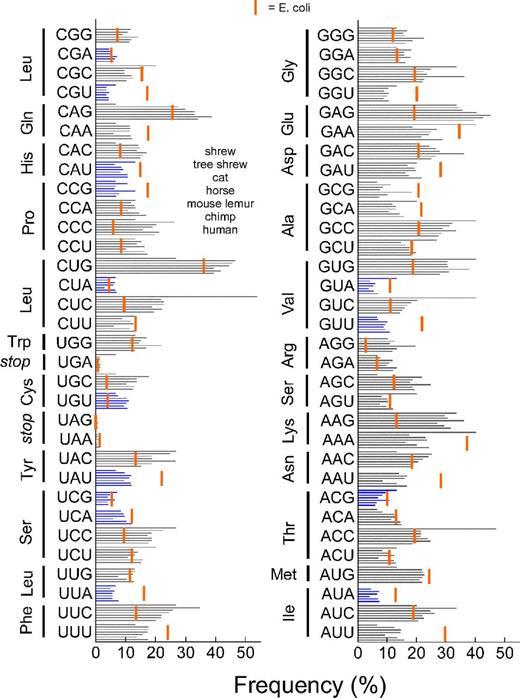
На рисунке синими полосками показаны частоты кодонов у землеройки, тупайи, домашней кошки, лошади, мышиного лемура, шимпанзе и человека. Красной полосой отмечена частота кодонов у кишечной палочки E. coli.
-
Система на основе регуляторных элементов лактозного оперона E. coli (lac-оперона) и сильного промотора.
Дело в том, что у кишечной палочки есть свои правила питания. Во-первых, существует механизм подавления активности lac-оперона, который включён только тогда, когда в клетку не поступает лактоза. Это логично: зачем тратить силы на синтез того, что не пригодится? Но как только лактоза начинает поступать в клетку в достаточном количестве этот механизм выключается.
Однако, есть второй механизм подавления активности lac-оперона. Если в среде есть глюкоза, то клетка питается исключительно глюкозой, так как она активирует второй механизм ингибирования транскрипции lac-оперона. Таким образом, lac-оперон активен только тогда, когда в окружающем клетку пространстве есть только лактоза. Минусом лактозного оперона является крайне слабый промотор, поэтому в штаммах продуцентах он заменён на сильный. Сильные промоторы часто получают из патогенов. Наиболее широко используемые в генной инженерии прокариот сильные промоторы выделены из бактериальных вирусов — бактериофагов. Например, широко используется промотор фага Т7.
К слову, некоторые сильные промоторы для генной инженерии растений также выделены из вирусов, например, это промотор вируса мозаики цветной капусты.
Как было указано выше, у E. coli нет РНК-полимеразы, которая бы узнавала промоторы бактериофагов, поэтому предварительно в продуцент встраивают ген РНК-полимеразы соответствующего бактериофага.
Можно поступить и по-другому: вообще не добавлять в питательную среду глюкозу и лактозу, а потом, когда культура дорастёт до нужной плотности, добавить то, что клетка примет за лактозу, но не сможет метаболизировать или разрушить. Сейчас в качестве такого индуктора используют ИПТГ.
Система на основе регуляторного механизма промотора pL бактериофага λ.
Данный промотор инактивируется белком-репрессором cI. При этом была обнаружена термочувствительная форма этого белка, названная cI857: данный фактор транскрипции сохраняет функциональность при температуре около 30⁰С и теряет её при 42⁰С. Поэтому при использовании такой системы сначала бактериальную культуру растят до нужной плотности при 30⁰С, а затем поднимают температуру до 42⁰С, тем самым запуская синтез целевого белка.
Ну что же, вектор разработан. Дальше дело за малым — найти подходящий метод для его внесения в бактериальную клетку. Но это уже совсем другая история.
Читайте также: