
Обратная транскрипция у ретровирусов
При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза.
Каждый вирион (полноценная вирусная частица, состоящая из нуклеиновой кислоты и белковой оболочки) ретровирусов содержит две идентичные цепи РНК размером от 8000 до 10 000 нуклеотидов. Области 5'- и 3'-концов обеих цепей модифицированы, как у всех эукариотических мРНК (5'-кэпы, З'-полиадениловые хвосты). Вирусные РНК имеют 5 структурных элементов: 1) прямые повторы на 5'- и З'-концах РНК (R); 2) последовательность из 80 -- 120 нуклеотидов, находящуюся около 5 концевого повтора (U5); 3) последовательность из 170 -- 1200 нуклеотидов около З'-концевого повтора (U3); 4) последовательность из 15 20 нуклеотидов (Р), в пределах которой клеточная тРНК комплементарно взаимодействует с ретровирусной РНК, что создает праймер для синтеза первой цепи ДНК; 5) сегмент Pu, находящийся непосредственно перед повтором U3 и являющийся сайтом для праймирования второй цепи ДНК -- такой сегмент одинаков у РНК всех ретровирусов определенного типа.
Этапы обратной транскрипции:
5. К моменту завершения синтеза первой цепи ДНК большая часть вирусной РНК разрушается РНКазой Н.
6. Синтез З'-конца второй цепи ДНК.
7. Удаление тРНК и оставшегося участка вирусной (+) РНК РНКазой Н.
8. Второй прыжок, в результате которого новосинтезированная вторая цепь ДНК комплементарно взаимодействует с тРНК-связывающей последовательностью первой цепи.
9. Удлинение З'-концов каждой цепи, образование дуплекса ДНК.
Вся последовательность реакций протекает без явного участия ферментов репликации клетки-хозяина (топоизомеразы, хеликазы, праймазы, ДНК-связывающего белка, лигазы и т.д.). При этом следует отметить, что молекулы вирусных ДНК длиннее молекул вирусных РНК, которые послужили матрицей для обратной транскрипции. Действительно, к 5'-концу (+) цепи вирусной ДНК добавилась последовательность U3, а к 3 -концу этой цепи -- последовательность U5. В результате на концах молекулы вирус специфической ДНК появился длинный (несколько сотен нуклеотидов) концевой повтор (ДКП или LTR.), имеющий структуру U3U5.

Рис.2. Схема обратной транскрипции ретровирусной РНК с образованием двуцепочечной ДНК (Darnell J., et.al. Molecular Cell Biology. - N. Y.: Scientific Amer. Books, 1986. - P. 1052)
Синтез ДНК на РНК-матрице in vitro и ревертаза используется в генетической инженерии для синтеза генов и их фрагментов, а также целенаправленного синтеза на матричных РНК комплементарных молекул ДНК (кДНК) для расшифровки первичной структуры РНК и белков.

Рис. 3. Схема получения кДНК с использованием ревертазы вируса и трех дополнительных ферментов: поли (А)-полимеразы, фрагмента Кленова ДНК-полимеразы I и нуклеазы S1. (Цитировано по)
Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А) - содержащих мРНК используют олигo (dT)-праймер, а для молекул РНК, не имеющих З'-поли (А) концов, -- химически синтезированные олигонуклеотиды, комплементарные З'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. сoli (остающаяся часть молекулы, которая сохраняет присущие ей каталитические активности).
Уже одно это открытие формально поставило РНК в центр основного постулата молекулярной генетики, так как показало, что поток генетической информации распространяется от РНК не в одном, а в двух направлениях: не только к белку, но и к ДНК. Все более глубокое проникновение в механизмы основных молекулярно-генетических процессов (репликацию, транскрипцию и трансляцию) способствовало возникновению понятия о неканонических функциях РНК, осознанию полифункциональности рибонуклеиновых кислот.
После проникновения происходит обратная транскрипция. Синтезированная ДНК – копия генома встраивается в геном клетки и использует клеточный механизм транскрипции

Параретровирусы
В капсиде в качестве генома содержится ДНК. ДНК сразу идет в ядро.Встраивается в геном клетки-хозяина с помощью интегразы.
ДНК - вирусы
Особенности жизненного цикла:
Ø Репликация и транскрипция происходит в ядре клетки (искл.: поксовирусы, асфавирусы).
Ø Для репликации вирусной ДНК необходима Ori – короткая нуклеотидная последовательность в вирусном геноме, к которым специфично присоединяются клеточные и вирусные факторы.
Ø Проблема недорепликации 5’-концов.

1)
2) Схема Кернса

3) Схема катящегося кольца
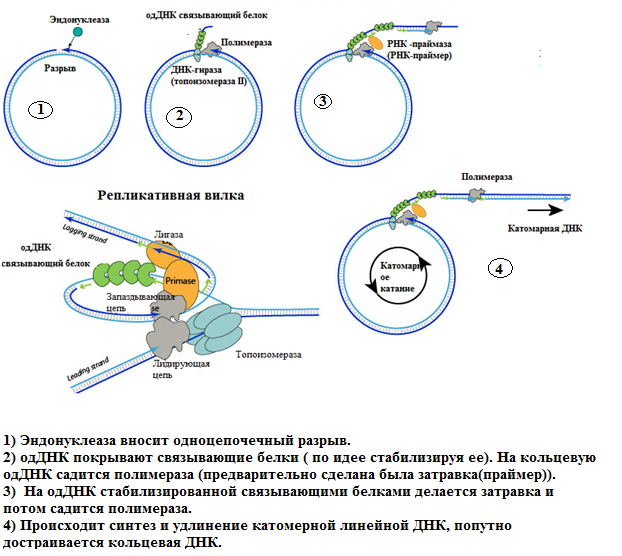
4) Самозатравочный механизм
Геном представлен линейной онДНК, оба конца которой имеют самокомплементарные (инвертированные повторы) последовательности, формирующие шпилечные структуры. Репликация вирусной ДНК осуществляется по самозатравочному механизму и осуществляется полимеразным комплексом из числа ранних вирусных белков: сначала эндонуклеаза вносит разрыв в цепи ДНК вблизи инвертированногоповтора с одного или двух концов (см. рис. 1.2.20, Б), после чего происходит расплетение концевой шпильки и синтез комплементарной цепи с использованием двунитевого участка в качестве затравки (см. рис. 1.2.20, В,Г), затем – отжиг концевых инвертированных повторов, которые используются как затравки для последующего синтеза комплементарных цепей ДНК (см. рис. 1.2.20, Д), в результате чего формируется либо пара геномов, либо конкатемер удвоенного вирусного генома (см. рис. 1.2.20, Е), и в последнем случае необходимо внесение двух эндонуклеазных разрывов цепи (см. рис. 1.2.20, Ж).

5) Аденовирусная репликация с использованием белков затравки


3. Тактические приёмы, используемые вирусами при экспрессии геномов. Сдвиг рамки считывания, протечка при сканировании, внутренняя инициация трансляции, рибосомальный пропуск, рибосомальное шунтирование.
Сдвиг рамки считывания – тип мутации в последовательности ДНК, для которого характерна вставка или делеция нуклеотидов, в количестве не кратном трем. В результате происходит сдвиг рамки считывания при транскрипции мРНК. Следует отличать от однонуклеотидного полиморфизма, в котором происходит замена одного нуклеотида на другой.
Внутренняя инициация трансляции (IRES) – регуляторная структура мРНК, задействованная в КЭП-независимом механизме инициации трансляции, при котором рибосома связывается с мРНК в области этой структуры в 5'-НТО недалеко от сайта инициации трансляции.
Рибосомальный пропуск – если после глицина идет пролин, происходит разрыв полипептида, при том что синтез полипептида продолжается. Вследствие этого, будут нарушены функции белка (или они изменятся) или же он станет нефункциональным.

Дата добавления: 2018-06-01 ; просмотров: 604 ;
Современная биология поражает уникальностью и масштабностью своих открытий. На сегодняшний день эта наука изучает большинство процессов, которые скрыты от нашего глаза. Этим примечательна молекулярная биология – одно из перспективных направлений, которое помогает разгадать сложнейшие тайны живой материи.
Что такое обратная транскрипция
Обратная транскрипция (сокращенно ОТ) – это специфический процесс, характерный для большинства РНК-содержащих вирусов. Главной его особенностью является синтез двухцепочечной молекулы ДНК на базе матричной РНК.
ОТ не характерна для бактерий или эукариотических организмов. Главный фермент – ревертаза – играет ключевую роль в синтезе двухцепочечной ДНК.
История открытия
Идея о том, что молекула рибонуклеиновой кислоты может стать матрицей для синтеза ДНК, считалась абсурдной вплоть до 1970 года. Тогда Балтимор и Темин, работавшие отдельно друг от друга, почти одновременно открыли новый фермент. Они назвали его РНК-зависимая-ДНК-полимераза, или обратная транскриптаза.
Открытие данного энзима безоговорочно подтвердило существование организмов, способных к обратной транскрипции. В 1975 году оба ученых получили Нобелевскую премию. Через некоторое время Энгельгардт предложил альтернативное название обратной транскриптазы – ревертаза.

Почему ОТ противоречит центральной догме молекулярной биологии
Центральная догма – это принципиальная схема последовательного синтеза белка в любой живой клетке. Такая схема строится из трех компонентов: ДНК, РНК и белок.
Согласно центральной догме, РНК может синтезироваться исключительно на матрице ДНК, а уж затем РНК участвует в построении первичной структуры белка.
Эта догма была официально принята в научном обществе раньше, чем произошло открытие обратной транскрипции. Неудивительно, что идея об обратном синтезе ДНК из РНК долго отвергалась учеными. Лишь в 1970 году вместе с открытием ревертазы была поставлена точка в этом вопросе, что отразилось и в принципиальной схеме синтеза белка.
Ревертаза ретровирусов птиц
Процесс обратной транскрипции не обходится без участия РНК-зависимой-ДНК-полимеразы. Максимально изучена на сегодняшний день ревертаза ретровируса птиц.
Всего около 40 молекул этого белка можно обнаружить в одном вирионе этого семейства вирусов. Протеин состоит из двух субъединиц, которые находятся в равном количестве и выполняют три важнейшие функции ревертазы:
1) Синтез молекулы ДНК как на матрице одноцепочечной/двухцепочечной РНК, так и на основе дезоксирибонуклеиновых кислот.
2) Активация РНКазы Н, главная роль которой заключается в расщеплении молекулы РНК в комплексе РНК-ДНК.
3) Разрушение участков молекул ДНК для встраивания в геном эукариот.
Механизм ОТ
Этапы обратной транскрипции могут варьироваться в зависимости от семейства вирусов, т.е. от вида их нуклеиновых кислот.
Рассмотрим сначала те вирусы, которые используют ревертазу. Здесь процесс ОТ делится на 3 этапа:
Такой способ размножения вирионов характерен для некоторых онкогенных вирусов и вируса иммунодефицита человека (ВИЧ).
Стоит отметить, что для синтеза любой нуклеиновой кислоты на матрице РНК нужна затравка, или праймер. Праймер представляет собой короткую последовательность нуклеотидов, комплементарную 3’-концу молекулы РНК (матрицы) и играющую важную роль в инициации синтеза.
Когда готовые двухцепочечные молекулы ДНК вирусного происхождения встраиваются в геном эукариот, запускается обычный механизм синтеза белков вириона. В итоге “захваченная” вирусом клетка становится фабрикой по производству вирионов, где в больших количествах образуются необходимые молекулы протеинов и РНК.
Повторение таких циклов приводит как к репликации генома вируса, так и к образованию мРНК, способных к синтезу белка в условиях зараженной эукариотической клетки.
Биологическое значение обратной транскрипции
Процесс ОТ имеет первостепенное значение в жизненном цикле многих вирусов (в первую очередь ретровирусов, таких как ВИЧ). РНК вириона, атаковавшего клетку эукариот, становится матрицей для синтеза первой цепочки ДНК, на которой не трудно достроить и вторую цепь.
Полученная двухцепочечная ДНК вируса встраивается в геном эукариот, что приводит к активации процессов синтеза белков вириона и появлению большого количество его копий внутри зараженной клетки. В этом и заключается основная миссия ревертазы и ОТ в целом для вируса.
У эукариот также может встречаться обратная транскрипция в контексте ретротранспозонов – мобильных генетических элементов, способных самостоятельно транспортироваться из одного участка генома в другой. Такие элементы, как считают ученые, стали причиной эволюции живых организмов.
Ретротранспозон представляет собой участок ДНК эукариот, который кодирует несколько белков. Один из них, ревертаза, принимает непосредственное участие в делокализации такого ретротраспорозона.
Использование ОТ в науке
С того момента, как удалось выделить ревертазу в чистом виде, процесс обратной транскрипции взяли на вооружение ученые-биологи. Изучение механизма ОТ и сейчас помогает прочитать последовательности важнейших белков человека.
Дело в том, что геном эукариот, в том числе и нас, содержит неинформативные участки под названием интроны. Когда с такой ДНК считывается последовательность нуклеотидов и формируется одноцепочечная РНК, последняя лишается интронов и кодирует исключительно белок. Если с помощью ревертазы на матрице РНК синтезировать ДНК, ее легко потом секвенировать и узнать порядок нуклеотидов.
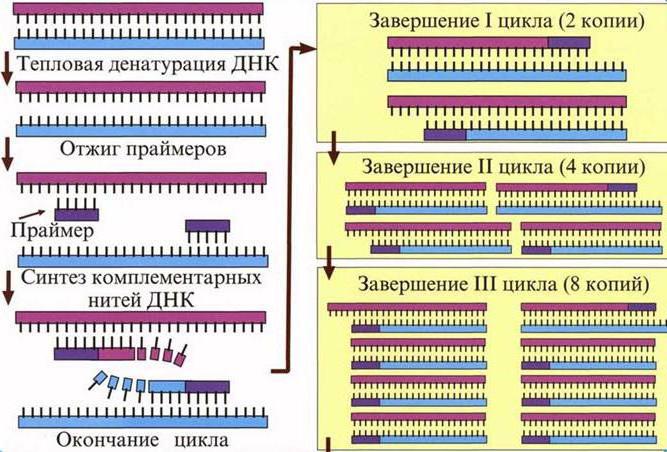
Нуклеиновая кислота, которая была образована с помощью обратной транскриптазы, называется кДНК. Она часто используется в полимеразной цепной реакции (ПЦР), чтобы искусственно увеличить число копий полученной копии кДНК. Данный метод используется не только в науке, но и в медицине: лаборанты определяют сходство такой ДНК с геномами различных бактерий или вирусов из общей библиотеки. Синтез векторов и внедрение их в бактерии – одно из перспективных направлений биологии. Если с помощью ОТ формировать ДНК человека и других организмов без интронов, такие молекулы легко внедряются в геном бактерий. Так последние становятся фабриками по производству необходимых человеку веществ (например, ферментов).
Вирионы сферической формы размером 80 — 100 нм, покрыты внешней липопротеиновой оболочкой, имеющей ворсинки длиной 8 — 10 нм. Внутри икосаэдрального капсида находится спиральный РНП. Наружная оболочка, капсидная мембрана и нуклеоид на разрезе вириона расположены концентрически. Чувствительны к эфиру, термолабильны, относительно резистентны к УФ-лучам. Характерной чертой семейства является наличие в составе вириона РНК-зависимой ДНК- полимеразы, иначе называемой обратной транскриптазой. Это и послужило основой для названия семейства (от лат. retro — обратный). Вирионы имеют 6 структурных белков, из них 4 внутренних (капсидных) негликолизированных и 2 гликопротеина оболочки.
Основными структурными генами кодирующими трансляцию белков, из которых в последующем строится вирус, являются gag (group — specificantigens), pol (polymerase), env (envelope). К регуляторным генам относятся: tat (трансактиватор всех вирусных белков), rev (регулятор экспрессии вирионных белков), vif (вирионный инфекционный фактор), vpr (функции остаются неясными), nef (негативный фактор экспрессии), vpx (функции неизвестны)
Капсидные белки несут группоспецифические межвидовые антигены и являются основой для разделения вирусов на роды и подроды. Гликопротеиды являются типоспецифическими антигенами, участвуют в реакции нейтрализации. Геном ретровирусов представлен однонитчатой РНК с молекулярной массой 7 мегадальтон и состоит из двух копий, каждая из которых является полноценным геномом и содержит одинаковую генетическую информацию, однако неизвестно, обе ли они функциональны. Нуклеиновая кислота онковирусов имеет гомологию с клеточной ДНК своего вида хозяина. Вирионная РНК неинфекционна. Вирусная РНК транскрибируется в ковалентно связанную двунитчатую ДНК, которая интегрируется с клеточной ДНК в виде ДНК-провируса. Провирус, экстрагированный из клетки, обладает инфекционностью. Многие вирусы этого семейства вызывают неопластические процессы, главным образом лейкемии и саркомы ряда видов животных. Нормальные клетки некоторых видов животных содержат интегрированные копии соответствующих видов онковирусов. Они могут никак не проявляться или активируются некоторыми физическими и химическими факторами, а возможно, и при инфекции другими онковирусами. Часто встречаются дефектные вирусы, размножающиеся с помощью вируса-помощника. Передаются вертикально и горизонтально. Ретровирусы, наши сожители, враги и помощники [3]
Retroviridae
Семейство Retroviridae включает три подсемейства:
- Oncovirinae (онковирусы), важнейший представитель которого — T-лимфотропный вирус человека типа 1 ;
- Lentivirinae (лентивирусы), к которому относится ВИЧ ; и
- Spumavirinae (спумавирусы, или пенящие вирусы).
Ретровирус раздваивает функции своего генетического материала: инфекционную функцию, то есть функцию самораспространения, выполняет вирусная РНК, а функцию экспрессии вирусных генов и синтеза молекул РНК, которые затем перенесут генетическую информацию в другие клетки, выполняет вирусная ДНК. Попадая внутрь клетки в ходе вирусной инфекции, ретровирусная РНК превращается в ДНК путем хорошо теперь известного процесса обратной транскрипции. Эта ДНК встраивается в геномную ДНК и с этого момента становится неотъемлемой частью генома клетки. А вирус становится провирусом. Провирус для животной клетки это то же самое, что профаг для бактериальной. Кстати, идеи лизогении, по-видимому, и привели Говарда Темина, который вместе с Дэвидом Балтимором открыл обратную транскрипцию, к идее провирусного состояния ретровирусов. По информационному содержанию ДНК-вариант генома ретровируса отличается от РНК-варианта только тем, что ДНК содержит не короткие концевые повторы, а длинные концевые повторы, LTR .
Особенности трансляции РНК ретровирусов
Находясь в составе геномной ДНК, вирусные гены транскрибируются под контролем LTR
LTR, long terminal repeats. Последовательности LTR включают в себя последовательности STR. Возникновение LTR очень важно для экспрессии вирусных генов. Они содержат вирусные регуляторные транскрипционные элементы: промотор, энхансер, и другие. Например, некоторые вирусы содержат элементы, определяющие зависимость вирусной транскрипции от наличия определенных гормонов. LTR и являются теми регуляторными сигналами, которые вирус использует для эксплуатации клеточной транскрипционной машины в своих целях.
Продуктом транскрипции является полноразмерная вирусная РНК. Она должна транслироваться. И здесь вирусу необходимо решить такую проблему: нужно синтезировать много белков, а РНК одна. И в клетках эукариот РНК моноцистронны, то есть предназначены для синтеза только одного белка. Синтез белка в большинстве случаев начинается с ближайшего к кэп-сайту инициирующего кодона [4] .
Если просмотреть открытую рамку считывания от этого ближайшего инициирующего кодона, то мы увидим, что если бы вирус пользовался традиционными способами экспрессии, то он смог бы синтезировать только полипептид GAG. А дальше идет стоп- кодон. Как быть с POL и ENV? Кроме того, эти полипептиды очень длинны, а в вирусе содержатся гораздо более короткие. Проблема решается несколькими способами.
Во-первых, с помощью сплайсинга эта одна РНК превращается в нашем упрощенном варианте еще в одну, более короткую. При этом последовательности, кодирующие ENV полипептид, оказываются рядом с инициирующим кодоном, ближайшим к кэп-сайту, и начинают транслироваться.
Во-вторых, разными для разных ретровирусов способами они ухитряются обойти стоп — кодон после открытой рамки считывания GAG и синтезировать сплавленный полипетид GAG-POL, который содержит последовательности обоих групп белков.
В-третьих, полученные длинные полипептиды подвергаются процессингу и разрезаются на множество белков, которые и функционируют либо в роли регуляторных, как, например, обратная транскриптаза, либо в роли структурных, как, например, белки оболочки зрелых вирусов.
Иными словами, ретровирусы используют гибкую тоталитарную систему для весьма тонкой регуляции синтеза большого разнообразия белков под контролем одного промотора.
Род Deltaretrovirus
Т-лимфотропный вирус человека — это человеческий РНК ретровирус, который вызывает такие злокачественные новообразования лимфоидной и кроветворной тканей, как Т-клеточный лейкоз и Т-клеточную лимфому у взрослых. Возможно вовлечение этого возбудителя в определенные демиелинизирующие болезни, включая тропический спастический парапарез. Взрослый Т-лимфотропный вирус (ВТЛВ) — это штамм вируса-возбудителя указанной болезни, который поражает преимущественно взрослых. Близкородственный вирус — это вирус лейкоза крупного рогатого скота. В настоящее время известно о четырех серотипах этого вида вируса.
См. также
Ссылки
- Цикл размножения ретровирусов (Flash-анимация)
- NCBI retrovirus book online
Литература
- Супотницкий М.В.Эволюционная патология. К вопросу о месте ВИЧ-инфекции и ВИЧ/СПИД-пандемии среди других инфекционных, эпидемических и пандемических процессов. — Москва: Вузовская книга, 2009. — 400 с. — ISBN 978-5-9502-0378-7
Примечания
Wikimedia Foundation . 2010 .
РЕТРОВИРУСЫ — РЕТРОВИРУСЫ, представители семейства ретровирусов (Retroviridale), чей генетический код, в отличие от других живых организмов, содержит РНК (рибонуклеиновую кислоту), а не обычную ДНК (дезоксирибонуклеиновую кислоту). Для размножения ретровирусы… … Научно-технический энциклопедический словарь
ретровирусы — семейство РНК–содержащих вирусов. Диаметр вирусных частиц 80 100 нм. Капсид икосаэдрический, заключен в липопротеидную оболочку. Содержит несколько фрагментов одноцепочечной линейной РНК (мол. масса 10–12 млн Да), обратную транскрипта–зу.… … Словарь микробиологии
РЕТРОВИРУСЫ — (от лат. retro назад и вирусы) семейство РНК содержащих вирусов. Обнаружены у всех позвоночных (в т. ч. человека) и некоторых беспозвоночных. Особенность ретровирусов наличие в жизненном цикле обратной транскрипции (синтез ДНК на матрице РНК),… … Большой Энциклопедический словарь
РЕТРОВИРУСЫ — (от лат. retro обратно, назад и вирусы), семейство РНК содержащих вирусов. Диам. вирусных частиц 80 100 нм. Капсид икосаэдрический, заключён в липопротеидную оболочку. Содержат неск. фрагментов одноцепочечной линейной РНК (общая мол. м. 10 12 000 … Биологический энциклопедический словарь
ретровирусы — лейковирусы Группа РНК содержащих вирусов диаметром 70 120 нм, капсид заключен в липопротеиновую оболочку, каждая частица включает по 2 идентичные молекулы РНК и связанные с ними молекулы обратной транскриптазы; многие Р. опухолеродны (вирус… … Справочник технического переводчика
Ретровирусы — * рэтравірусы * retroviruses класс однонитчатых РНКовых вирусов, инфицирующих клетки эукариот, в которых РНК геном транскрибируется в ДНК копию (кДНК) с помощью РНК зависимой ДНК полимеразы (обратной транскриптазы, см.). Эта двунитчатая кДНК… … Генетика. Энциклопедический словарь
РЕТРОВИРУСЫ — вирусы с необычным способом репликации генетического материала. Для цикла репродукции этого большого семейства вирусов характерен обратный поток генетической информации: вместо обычной транскрипции (т.е. переписывания) дезоксирибонуклеиновой… … Энциклопедия Кольера
ретровирусы — (от лат. retro назад и вирусы), семейство РНК содержащих вирусов. Обнаружены у всех позвоночных (в том числе человека) и некоторых беспозвоночных. Особенность ретровирусов наличие в жизненном цикле обратной транскрипции (синтез ДНК на матрице… … Энциклопедический словарь
ретровирусы — (Retraviridae; син. вирусы лейкозов) семейство вирусов, вирионы которых содержат нуклеокапсид и имеют наружную оболочку; геном представлен однонитчатой молекулой рибонуклеиновой кислоты; в вирионе Р. содержится ревертаза (обратная транскриптаза); … Большой медицинский словарь
Ретрови́русы (лат. Retroviridae) — семейство РНК-содержащих вирусов, заражающих преимущественно позвоночных. Наиболее известный и активно изучаемый представитель — вирус иммунодефицита человека.
После инфицирования клетки ретровирусом в цитоплазме начинается синтез вирусного ДНК-генома с использованием вирионной РНК в качестве матрицы. Все ретровирусы используют для репликации своего генома механизм обратной транскрипции: вирусный фермент обратная транскриптаза (или ревертаза) синтезирует одну нить ДНК на матрице вирусной РНК, а затем уже на матрице синтезированной нити ДНК достраивает вторую, комплементарную ей нить. Образуется двунитевая молекула ДНК, которая, проникнув через ядерную оболочку, интегрируется в хромосомную ДНК клетки и далее служит матрицей для синтеза молекул вирусных РНК. Эти РНК выходят из клеточного ядра и в цитоплазме клетки упаковываются в вирусные частицы, способные инфицировать новые клетки.
По одной из гипотез, ретровирусы могли произойти от ретротранспозонов — подвижных участков генома эукариот.
Идея обратной транскрипции вначале была очень непопулярна, так как противоречила центральной догме молекулярной биологии, которая предполагала, что ДНК транскрибируется в РНК и далее транслируется в белки.
Однако в 1970 году Темин и Балтимор независимо друг от друга открыли фермент, названный обратной транскриптазой (ревертазой), и возможность обратной транскрипции была окончательно подтверждена. В 1975 году Темину и Балтимору была присуждена Нобелевская премия в области физиологии и медицины.
ВИЧ — вирус иммунодефицита человека, вызывающий заболевание — ВИЧ-инфекцию, последняя стадия которой известна как синдром приобретённого иммунодефицита (СПИД) — в отличие от врождённого иммунодефицита.
Попадая в организм человека, ВИЧ заражает CD4+ лимфоциты, макрофаги и некоторые другие типы клеток. Проникнув в клетки указанных типов, вирус начинает активно в них размножаться. Это в конечном счёте приводит к разрушению и гибели зараженных клеток. Присутствие ВИЧ со временем вызывает нарушение иммунной системы из-за избирательного уничтожения им иммунокомпетентных клеток и подавления их субпопуляции. Вышедшие из клетки вирусы внедряются в новые, и цикл повторяется. Постепенно число CD4+ лимфоцитов снижается настолько, что организм уже не может противостоять возбудителям оппортунистических инфекций, которые не опасны или мало опасны для здоровых людей с нормально функционирующей иммунной системой.
Для диагностики поражения слизистой оболочки рта у ВИЧ-инфицированных больных принята рабочая классификация, утверждённая в Лондоне, в сентябре 1992 года. Все поражения разделены на 3 группы:
ü 1 группа — поражения, чётко связанные с ВИЧ-инфекцией. В эту группу включены следующие нозологические формы:
- кандидозы (эритематозный, псевдомембранозный, гиперпластический, атрофический);
- волосистая лейкоплакия;
- маргинальный гингивит;
- язвенно-некротический гингивит;
- деструктивный пародонтит;
- саркома Капоши;
- неходжкинская лимфома.
ü 2 группа — поражения, менее чётко связанные с ВИЧ-инфекцией:
- бактериальные инфекции;
- болезни слюнных желёз;
- вирусные инфекции;
- тромбоцитопеническая пурпура.
ü 3 группа — поражения, которые могут быть при ВИЧ-инфекции, но не связанные с нею.
Наибольший интерес вызывают и наиболее часто встречаются поражения, относящиеся к 1 группе. В России при постановке диагноза ВИЧ-инфекция проводится дотестовое и послетестовое консультирование пациента, разъяснение основных фактов о заболевании. Пациенту предлагается встать на учет в территориальный центр профилактики и борьбы со СПИД для бесплатного диспансерного наблюдения врачом-инфекционистом. Приблизительно раз в полгода рекомендуется сдавать анализы (на иммунный статус и на вирусную нагрузку) для контроля состояния здоровья. В случае значительного ухудшения этих показателей рекомендуется прием антиретровирусных препаратов (терапия бесплатна, доступна практически во всех регионах).
Читайте также: