
Одноцепочечные рнк содержащие вирусы

Вирусы — это микроскопические патогены, заражающие клетки живых организмов для самовоспроизводства. Они состоят из одного вида нуклеиновой кислоты (или ДНК или РНК, но не обе вместе), которая защищена оболочкой, содержащей белки, липиды, углеводы или их комбинацию. Размер типичного вируса варьируется от 15 до 350 нм, поэтому его можно увидеть только с помощью электронного микроскопа.
В 1892 году русский ученый Д.И. Ивановский впервые доказал существование ранее неизвестного типа возбудителя болезней, это был вирус мозаичной болезни табака. А в 1898 году Фридрих Лоффлер и Пол Фрош нашли доказательства того, что причиной ящура у домашнего скота была инфекционная частица, которая меньше, чем любая бактерия. Это были первые шаги к изучению природы вирусов, генетических образований, которые лежат где-то в серой зоне между живыми и неживыми состояниями материи. На текущий момент описано около 6 тыс. вирусов, но их существует несколько миллионов.
Строение вирусов
Вне клеток-хозяев вирусы существуют в виде белковой оболочки (капсида), иногда заключенного в белково-липидную мембрану. Капсид обволакивает собой либо ДНК, либо РНК, которая кодирует элементы вируса. Находясь в такой форме вне клетки, вирус метаболически инертен и называется вирионом.
Простая структура, отсутствие органелл и собственного метаболизма позволяет некоторым вирусам кристаллизоваться, т.е. они могут вести себя подобно химическим веществам. С появлением электронных микроскопов было установлено, что их кристаллы состоят из тесно прижатых друг к другу нескольких сотен миллиардов частиц. В одном кристалле вируса полиомиелита столько частиц, что ими можно заразить не по одному разу всех жителей Земли.
Формы вирусов
Вирусы встречаются в трех основных формах. Они бывают:
- Сферическими (кубическими или полигидральными). Вирусы герпеса, типулы, полиомы и т.д.
- Спиральными (цилиндрическими или стержнеобразными). Вирусы табачной мозаики, гриппа, эпидемического паротита и др.
- Сложными. Например, бактериофаги.
Проникновение вирусов в клетку-хозяина
Капсид в основном защищает нуклеиновую кислоту от действия клеточного нуклеазного фермента. Но некоторые белки капсида способствуют связыванию вируса с поверхностью клеток-хозяев, и работают, как ключики, вставляемые в нужные замочки. Другие поверхностные белки действуют как ферменты, они растворяют поверхностный слой клетки-хозяина и таким образом помогают проникновению нуклеиновой кислоты вируса в клетку-хозяина.
Жизненный цикл вирусов сильно отличается у разных видов, но существует шесть основных этапов жизненного цикла вирусов:
Присоединение к клетке-хозяину представляет собой специфическое связывание между вирусными капсидными белками и рецепторами на клеточной поверхности. Эта специфика определяет хозяина вируса.
Проникновение следует за прикреплением: вирионы проникают в клетку-хозяина через рецептор-опосредованный эндоцитоз или слияние мембран. Это часто называют вирусной записью.
Проникновение вирусов в клетку достигается за счет:
Размножение вирусов
После того, как вирусный геном освобождается от капсида, начинается его транскрипция или трансляция. Именно эта стадия вирусной репликации сильно различается между ДНК- и РНК-вирусами и вирусами с противоположной полярностью нуклеиновой кислоты. Этот процесс завершается синтезом новых вирусных белков и генома (точных копий внедрённых).
Механизм репликации зависит от вирусного генома.
- ДНК-вирусы обычно используют белки и ферменты клетки-хозяина для получения дополнительной ДНК, она транскрибируется в РНК-мессенджер (мРНК), которая затем используется для управления синтезом белка.
- РНК-вирусы обычно используют ядро РНК в качестве матрицы для синтеза вирусной геномной РНК и мРНК. Вирусная мРНК направляет клетку-хозяина на синтез вирусных ферментов и капсидных белков и сборку новых вирионов. Конечно, есть исключения из этого шаблона. Если клетка-хозяин не обеспечивает ферменты, необходимые для репликации вируса, вирусные гены предоставляют информацию для прямого синтеза отсутствующих белков.
Чтобы преобразовать РНК в ДНК, вирусы должны содержать гены, которые кодируют вирус-специфический фермент обратной транскриптазы. Она транскрибирует матрицу РНК в ДНК. Обратная транскрипция никогда не происходит в неинфицированных клетках. Необходимый фермент, обратная транскриптаза, происходит только от экспрессии вирусных генов в инфицированных клетках.
Вироиды
Вироиды заражают только растения. Одни вызывают экономически важные заболевания сельскохозяйственных культур, в то время как другие являются доброкачественными. Двумя примерами экономически важных вироидов являются кокосный cadang-cadang (он вызывает массовую гибель кокосовых пальм) и вироид рубцовой кожицы яблок, который безнадежно портит товарный вид яблок.
30 известных вироидов были классифицированы в две семьи.
- Члены семейства Pospiviroidae, названные по имени вироида клубневого веретена картофеля, имеют палочковидную вторичную структуру с небольшими одноцепочечными областями, имеет центральную консервативную область, и реплицируются в ядре клетки.
- Avsunviroidae, названный в честь вироида авокадо, имеет как палочковидную, так и разветвленную области, но не имеет центральной консервативной области и реплицируется в хлоропластах растительной клетки.
В отличие от вирусов, которые являются паразитами механизма трансляции хозяина, вироиды являются паразитами клеточных транскрипционных белков.
Бактериофаги

Существуют тысячи разновидностей фагов, каждый из которых может заразить только один тип или несколько близких типов бактерий или архей. Фаги классифицируются по ряду семейств вирусов; например:
Как и все вирусы, фаги являются простыми организмами, которые состоят из ядра генетического материала (нуклеиновой кислоты), окруженного капсидом белка. Нуклеиновая кислота может представлять собой либо ДНК, либо РНК, и может быть двухцепочечной или одноцепочечной.
Существует три основных структурных формы фага:
- Икосаэдрическая (20-сторонняя) головка с хвостом
- Икосаэдрическая головка без хвоста
- Нитевидная форма
Во время заражения фаг прикрепляется к бактерии и вставляет в нее свой генетический материал. После этого фаг обычно следует одному из двух жизненных циклов: литическому (вирулентному) или лизогенному (умеренному).
Литические, или вирулентные, фаги захватывают механизм клетки, чтобы скопировать компоненты фага. Затем они разрушают или лизируют клетку, высвобождая новые частицы фага.
Лизогенные, или умеренные, фаги включают свою нуклеиновую кислоту в хромосому клетки-хозяина и реплицируются с ней как единое целое, не разрушая клетку. При определенных условиях лизогенные фаги могут индуцироваться в соответствии с литическим циклом.
Существуют и другие жизненные циклы, в т.ч. псевдолизогенез и хроническая инфекция. При псевдолизогении бактериофаг проникает в клетку, но не использует механизм репликации клеток и не интегрируется в геном хозяина, просто как бы прячется внутри бактерии, не нанося ей никакого вреда. Псевдолизогенез возникает, когда клетка-хозяин сталкивается с неблагоприятными условиями роста и, по-видимому, играет важную роль в выживании фага, обеспечивая сохранение генома фага до тех пор, пока условия роста хозяина снова не станут благоприятными.
При хронической инфекции новые фаговые частицы образуются непрерывно и длительно, но без явного уничтожения клеток.
Вскоре после открытия фаги начали использовать для лечения бактериальных заболеваний человека, таких как бубонная чума и холера. Но фаговая терапия тогда не была успешной, и после открытия антибиотиков в 1940-х годах она была практически заброшена. Однако с появлением устойчивых к антибиотикам бактерий терапевтическому потенциалу фагов уделяется все больше внимания.
Наше время с антибиотиками заканчивается. В 2016 году женщина в штате Невада умерла от бактериальной инфекции, вызванной Klebsiella pneumoniae, которая была устойчивой ко всем известным антибиотикам. Бактерии, устойчивые к колистину, антибиотику последней инстанции, были обнаружены на свинофермах в Китае. В настоящее время бактерии приспосабливаются к антибиотикам быстрее, чем когда-либо.
Покажите ножницы которыми вирусы разрезают молекулу РНК что бы встроиться для мутации.Может что нибудь придумаете другое.К примеру деление цепочка аминокислот получив энергию из вне как одноименные заряды распадается на две. К каждой соединятся только те какие были ранее (другие проскочат мимо),казалось бы копии,но внутренняя энергия разная(уменьшается увеличивается) поэтому распад и создание. Вся химия углерода на этом построена 1000 орган соединений создает у других хим элементов этого свойства нет. Иммунная система делает накладку(интерференция)с помощью энергии интерферонов пытаясь разрушить цепочку РНК вируса.Надо помочь организму но не вакциной(вирус быстро мутирует)
Экспрессия и репликация вирусного генома
Требования и ограничения
В ходе эволюции вирусов сложилось несколько стратегий, обеспечивающих: а) организацию вирусных генов и их кодирующую функцию, б) экспрессию вирусных генов, в) репликацию вирусных геномов и г) сборку и созревание вирусного потомства.
Прежде чем мы рассмотрим каждое из этих положений в деталях, стоит напомнить, что ключевым моментом в репликации вирусов является использование для синтеза вирусных белков хозяйских структур, синтезирующих белки клетки. Независимо от размеров, состава и организации своего генома вирус должен предоставить белоксинтезирующему аппарату эукариоти-ческой клетки информационную РНК, которую клетка должна распознать и транслировать. В этом отношении клетка навязывает вирусу два ограничения.
Во-первых, клетка синтезирует в ядре свою собственную мРНК путем транскрипции своей ДНК и последующего постранскрипционного процессинга транскрипта. Поэтому в клетке: а) ни в ядре, ни в цитоплазме нет фер* ментов, необходимых для транскрипции мРНК с вирусного РНК-генома, и б) в цитоплазме нет ферментов, способных транскрибировать вирусную ДНК. В связи с этим клеточную транскриптазу для синтеза вирусных мРНК могут использовать только вирусы, содержащие ДНК и способные проникать в ядро. Все другие вирусы вынуждены создавать собственные ферменты для синтеза мРНК.
Для удобства сначала лучше обсудить РНК-содержащие вирусы, фокусируя внимание прежде всего на функции геномной РНК.
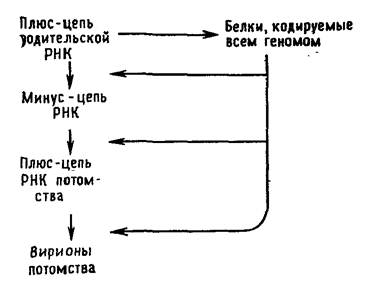
Вирусы с одноцепочечной РНК можно подразделить на три группы. В первую группу входят пикорнавирусы и тогавирусы. Их геномы выполняют две функции (рис. 5.2 и 5.3). Во-первых, они функционируют как мРНК. Вирусы, геном которых может служить в качестве мРНК, принято называть вирусами с позитивным геномом. РНК пикорнавирусов после проникновения в клетку связывается с рибосомами и полностью транслируется. Затем продукт этой трансляции — полипротеин — расщепляется. Во-вторых, геномные РНК выполняют функцию матрицы для синтеза на ней комплементарных минус-цепей при участии полимеразы, появившейся в результате расщепления полипротеина. Образуется двуспиральная репликативная форма. На ее минус-цепях синтезируются новые плюс-цепи, которые могут использоваться в качестве а) мРНК, б) матриц для синтеза новых минус-цепей и в) составной части вирусных частиц потомства.
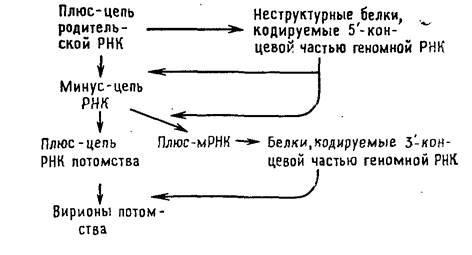
Тогавирусы и ряд других вирусов с позитивным геномом отличаются от пикорнавирусов в одном отношении: для трансляции в первом цикле синтеза белков доступна только часть их геномной РНК. Вероятной функцией образующихся при этом белков является способность транскрибировать геномную РНК. Они осуществляют синтез минус-цепи, которая в свою очередь служит матрицей для синтеза двух различных по размеру классов молекул плюс-РНК. В клетках, зараженных тогавирусами, РНК первого класса представлены небольшими молекулами мРНК, фланкирующими участок геномной РНК, который не транслировался в первом цикле. Образующиеся на них полипротеины расщепляются на белки, которые играют структурную роль в вирионах. Плюс-РНК второго класса состоит из полноразмерных цепей, которые упаковываются в вирионы.
РНК-содержащие вирусы с негативным геномом: ортомиксовирусы, парамиксовирусы, буньявирусы, аренавирусы и рабдовирусы.
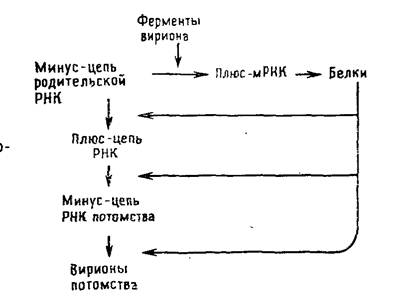
Их геномная РНК выполняет две матричные функции: во-первых, для транскрипции и, во-вторых, для репликации. В связи с тем что для синтеза мРНК должен транскрибироваться вирусный геном, а в клетках соответствующие ферменты отсутствуют, все вирусы с негативным геномом содержат в вирионе кроме вирусного генома транскриптазу. Транскрипция вирусного генома — первое событие после проникновения вируса в клетку, в результате которого накапливаются функционально активные моноцистронные мРНК [позитивные, или плюс-цепи], кодирующие один белок. Репликацию начинают новосинтезированные вирусные белки, катализирующие образование полной плюс-цепи, которая служит матрицей для синтеза геномной минус-РНК (рис. 5.4).
Размножение ортомиксовирусов и парамиксовирусов.
Ортомиксовирусы (вирусы гриппа А; В;С) генм представлен не одной а набором односпиральных РНК (грипп птиц и человека 8-7фрагментов). В структуре вириона:
– РНК зависимая РНК полимераза (синтез +нитей,
– две разновидности РНК полимераз
РНК транскриптаза (синтез +нитей в начале инфекции,
РНК репликаза ( – нитей в конце инфекции)
Ретровирусы входят в третью группу РНК-содержащих вируссов.
Характерно, что геномы ретровирусов монолитны, но имеют диплоидную структуру, и обе цепи либо частично соединены водородными связями друг с другом, либо спарены неизвестным до настоящего времени образом. Единственная известная функция геномной РНК — матричная функция для синтеза вирусной ДНК. Поскольку эукариотические клетки не имеют для этого соответствующих ферментов, вирион кроме генома содержит еще и РНК-зависимую ДНК-полимеразу (обратную транскриптазу 1), а также смесь тРНК хозяина, одна из которых служит в качестве затравки. В цикле репродукции можно выделить следующие ключевые ступени: а) связывание комплекса тРНК —обратная транскриптаза с геномной РНК; б) синтез ДНК-копии, комплементарной по отношению к РНК, с переходом полимеразы с одной молекулы РНК-матрицы на другую, что приводит к образованию кольцевой одноцепочечной молекулы ДНК, связанной водородными связями с линейной геномной РНК; в) расщепление геномной РНК нуклеазой, атакующей только РНК в ДНК—РНК-гибридах (рибонуклеазои Н, также содержащейся в вирионе), и г) синтез комплементарной копии вирусной ДНК. Затем кольцевая двухцепочечная ДНК перемещается в ядро, где интегрирует с геномом хозяина, но последующая экспрессия вирусных генов не обязательна. Если экспрессия происходит, то интегрированная.вирусная ДНК транскрибируется транскриптазой клетки-хозяина. Продуктами транскрипции являются молекулы РНК, как равные по длине молекуле генома, так и более короткие мРНК-транскрипты нескольких соседних генов, которые транслируются с образованием полипротеинов. Полипротеины затем расщепляются на отдельные вирусные белки. В состав вирионов включаются только транскрипты, содержащие весь геном.
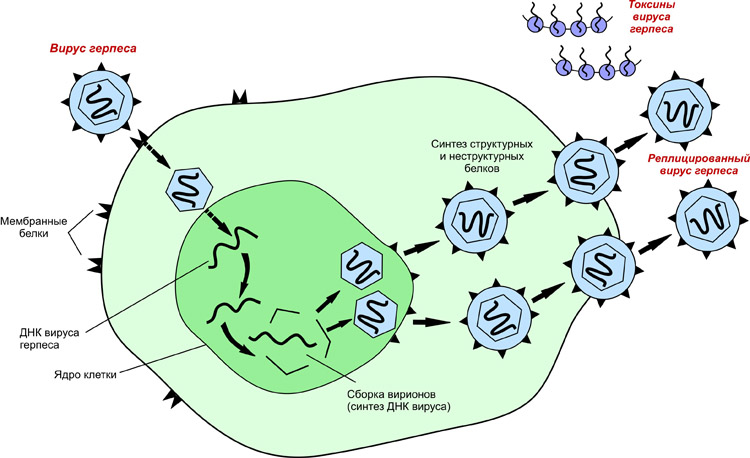
Вирусы, содержащие двуцепочечную ДНК для репликации попадают в ядро клетки, так как им требуется клеточная ДНК-полимераза. Также репликация ДНК этих вирусов сильно зависит от стадии клеточного цикла. В некоторых случаях вирус может вызывать деления клетки, что может приводить к раковому перерождению. Примерами таких вирусов являются Herpesviridae, Adenoviridae и Papovaviridae.
У представителей семейства Poxvirus геномная ДНК реплицируется не в ядре.
Вирусы семейств Circoviridae и Parvoviridae реплицируют геномную ДНК в ядре и в ходе репликации образуют интермедиат — двуцепочечную ДНК.
Как и большинство РНК-вирусов, представители класса III реплицируют геномную РНК в цитоплазме и используют полимеразы хозяина в меньшей степени, чем ДНК-вирусы. Класс III включает в себя два крупных семейства Reoviridae и Birnaviridae. Репликация моноцистронная, геном сегментирован, каждый ген кодирует один белок.
Классы IV и V включают вирусы двух типов, репликация которых не зависит от стадии клеточного цикла. Наряду с вирусами, содержащими двуцепочечную ДНК, эти вирусы наиболее изучены.
Непосредственно на (+) геномной РНК вирусов IV класса может идти синтез белка на рибосомах клетки хозяина. Вирусы классифицируют на две группы, в зависимости от особенностей РНК:
- у вирусов с полицистронной мРНК трансляция приводит к образованию полипротеина, который затем разрезается на зрелые белки. С одной цепи РНК может синтезироваться несколько разных белков, что снижает длину генов.
- вирусы со сложной транскрипцией содержат субгеномные мРНК, синтез белка идет со сдвигом рамки считывания, также используется протеолитический процессинг полипротеинов. Эти механизмы обеспечивают синтез разных белков с одной цепи РНК.
Примеры вирусов данного класса включают представителей семейств Astroviridae, Caliciviridae, Coronaviridae, Flaviviridae, Picornaviridae, Arteriviridae и Togaviridae.
Геномные РНК вирусов класса V не могут быть транслированы на рибосомах клетки хозяина, предварительно требуется транскрипция вирусными РНК-полимеразами в (+)РНК. Вирусы пятого класса классификации по Балтимору классифицируют на две группы:
- вирусы, содержащие несегментированный геном, на первом этапе репликации происходит транскрипция (−)РНК вирусной РНК-зависимой РНК-полимеразной в моноцистронную мРНК, и далее синтезируются дополнительные копии (+)РНК, служащие матрицами для синтеза геномных (−)РНК. Репликация геномных РНК таких вирусов осуществляется в цитоплазме.
- вирусы с сегментированными геномами, репликация геномных РНК которых происходит в клеточном ядре, вирусная РНК-зависимая РНК-полимераза синтезирует моноцистронные мРНК с каждого сегмента генома. Наибольшим отличием данной группы вирусов от другой группы пятого класса состоит в том, что репликация осуществляется в двух местах.
Представители данного класса входят в состав семейств: Arenaviridae, Orthomyxoviridae, Paramyxoviridae, Bunyaviridae, Filoviridae и Rhabdoviridae.
Наиболее хорошо изученным семейством данного класса вирусов, являются ретровирусы. Вирусы класса VI используют фермент обратную транскриптазу для превращения (+)РНК в ДНК. Вместо использования РНК в качестве матрицы для синтеза белков, вирусы этого класса используют матрицу ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина. Наиболее хорошо изученным представителем данной группы вирусов является ВИЧ.
Небольшая группа вирусов, в состав которой входит вирус гепатита В, представитель семейства Hepadnaviridae, имеют двуцепочечную геномную ДНК, которая ковалентно замкнута в форме кольца и является матрицей для синтеза мРНК вируса, а также субгеномных РНК. Субгеномная РНК служит матрицей для синтеза ДНК-генома ферментом обратной транскриптазой вируса.
Одни вирусы способны интегрироваться в геном клетки-мишени и таким образом оставаться во всех дочерних клетках, которые будут в будущем получены после ее деления. К таким вирусам относятся гаммаретровирусы и лентивирусы. Другие делать этого не умеют (например, адено- и аденоассоциированные вирусы). Но для производства белков и репликации (размножения) все они используют клетку и ее синтетический аппарат.
Как это работает?
Для того чтобы вирус мог проникнуть в клетку, белки его оболочки должны связаться с мембранными белками клетки-мишени. Важно отметить, что проникает вирус только в те клетки, которые могут в дальнейшем помочь его репликации. Вирус ВИЧ живет в клетках иммунной системы, вирус гепатита С — в клетках печени. Есть особые вирусы, которые поражают только растения или даже только бактерии.
Содержащие неактивный вирус и оставшиеся в живых клетки иногда сохраняют возможность нормального функционирования. В этом случае клетки могут быть заражены, но вирус проявит себя спустя длительный период времени. Так устроен герпес.
В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы. И тут стоит остановиться на классификации.
Типы вирусов. Коротко о главном
Современная типология вирусов содержит 7 классов и была предложена Дэвидом Балтимором еще в 1971 году. С тех пор, впрочем, она была уточнена и расширена, в том числе советскими учеными. И выглядит в настоящее время таким образом:
- Класс I
Вирусы, содержащие двухцепочечную ДНК
Описание
Для репликации вирусу необходимо попасть в ядро клетки-мишени и воспользоваться ее ДНК-полимеразой. Иногда вирус вызывает незапланированное деление самое клетки, то есть становится онкогенным. Эти вирусы хорошо изучены.
Пример: Вирус герпеса, адено- и папилломавирусы
Вирусы, содержащие одноцепочечную ДНК
Описание
Попадая в ядро клетки, вирусы образуют двухцепочечную ДНК, после чего реплицируются так же, как вирусы класса I.
Пример: Парво- и цирковирусы
Вирусы, в которых РНК способна к репликации (редупликации)
Описание
Вирусы этого класса могут размножаться в цитоплазме клетки, им не нужна молекула ДНК. Каждый ген, находящийся в РНК вируса, кодирует только один вирусный белок.
Пример: Бирна- и реовирусы
Вирусы, содержащие одноцепочечную (+) РНК
Описание
Из геномной (+) РНК на рибосомах хозяина создаются вирусные белки. В одном фрагменте РНК могут быть закодированы разные белки, что увеличивает сложность вируса без удлинения генов.
Пример: Пикорнавирусы (полиомиелит, гепатит А) и коронавирусы
Вирусы, содержащие одноцепочечную (–) РНК
Описание
(–) РНК этих вирусов предварительно должна быть транскрибирована в (+) РНК вирусными РНК-полимеразами, после чего может начаться синтез вирусных белков. Вирусы этого класса делятся еще на две группы, в зависимости от их генома и места его репликации (цитоплазма или ядро).
Пример: Филовирусы, аренавирусы (геморрагическая лихорадка Ласса), ортомиксовирусы (вирусы гриппа) и так далее.
Вирусы, содержащие одноцепочечную (+) РНК, реплицирующиеся через стадию ДНК
Описание
Такие вирусы используют фермент обратную транскриптазу для превращения (+) РНК в ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина.
Пример: Ретровирусы (в том числе ВИЧ)
Вирусы, содержащие двухцепочечную ДНК, реплицирующиеся через стадию одноцепочечной РНК
Описание
Молекула ДНК замкнута в кольцо и является матрицей для синтеза мРНК и дополнительных молекул РНК, которые используются при репликации вирусного генома обратными транскриптазами.
Пример: Колимовирусы (вызывают инфекции растений) и гепаднавирусы (например, гепатит В)
Вакцинация и лечение
Как правило, организмы умеют бороться с паразитирующими на них вирусами. На примере млекопитающих и человека мы обычно говорим о главном инструменте — врожденном иммунитете.
Впрочем, наиболее эффективен этот вид защиты в отношении бактериальных инфекций и не может обеспечить продолжительную и надежную защиту, особенно от инфекций вирусных.
Именно поэтому огромное значение имеет приобретенный иммунитет, в результате которого клетки иммунной системы обучаются вырабатывать специфические к вирусу антитела, способные уничтожать как саму вирусную частицу, так и зараженные ею клетки.
Еще одна врожденная система борьбы с вирусными инфекциями — внутриклеточная. Как правило, клетка способна распознать чужеродную РНК в своей цитоплазме, куда ее сперва и доставляют многие вирусы, и имеет специальные комплексы для ее деградации. Но часть вирусов научились обходить и эту ловушку. К примеру, ротавирусы, которые даже внутри клетки сохраняют капсид с геномной РНК.
Миссия: уничтожить
Основная сложность в лечении вирусных заболеваний заключается в том, что они используют естественные функции клеток-мишеней для своего размножения, поэтому ученым зачастую оказывается не так-то просто придумать препарат, который будет токсичен для вируса и безопасен для самой клетки. Если такой безопасности достичь не удастся, лекарство будет иметь слишком много побочных эффектов, повреждающих сам организм, что окажется нецелесообразно для использования.
По принципу действия противовирусные препараты подразделяются на две группы: стимулирующие иммунную систему атаковать вирусы (например, за счет индукции синтеза белков-интерферонов) и атакующие вирусы напрямую. Препараты второй группы различаются по этапу жизненного цикла вируса, на котором они активны: это препараты, препятствующие проникновению вируса в клетку, препятствующие размножению вируса внутри клетки и препятствующие выходу копий вируса из клетки.
Чтобы помешать проникновению вируса, препарат должен заблокировать рецептор на клетке, с которым связывается вирусная частица. Так работает, например, ибализумаб — зарегистрированный в США новый препарат против ВИЧ, о котором мы недавно писали.
Еще один класс противовирусных препаратов блокирует ферменты, необходимые для создания и модификаций белков вируса. Такие лекарства называют протеазными ингибиторами.
Вместо заключения: а могут ли вирусы приносить пользу?
Безусловно, да. Несмотря на то, что вирусы ассоциируются у большинства людей с однозначным вредом, они могут приносить и пользу — если речь идет о так называемых вирусных векторах и терапевтических подходах на их основе.
Исследователи давно научились помещать в белковую оболочку вируса интересующие их нуклеиновые кислоты, чтобы доставлять нужный ген в клетки, а также убирать те гены, которые делают вирус опасным для организма.
Это позволило сделать возможной генную терапию, помогающую бороться с заболеваниями, вызванными известными генетическими мутациями. Создание вирусных векторов — достаточно непростая задача, к тому же ограниченная свойствами самих вирусных частиц: количеством помещающейся генетической информации, местом ее вставки, стабильностью. Кроме того, вирусный вектор, используемый в медицине, не должен вызывать иммунного ответа или критично влиять на жизнедеятельность клетки. Тем не менее эти сложности решаются, поэтому уже одобрен ряд вполне успешных и безопасных генных терапий. А в качестве основы для вирусных векторов чаще всего используются ретро-, ленти-, адено- и аденоассоциированные вирусы.
РНК - вирус представляет собой вирус , который имеет РНК (рибонуклеиновая кислота) в качестве своего генетического материала . Это нуклеиновой кислоты , как правило , одноцепочечной РНК ( оцРНК ) , но может быть двухцепочечной РНК (дцРНК). Известные заболевания человека , вызываемые вирусами РНК включают Эбола hemorrhoragic лихорадка , SARS , Бешенство , простуду , грипп , гепатит С , лихорадки Западного Нила , полиомиелит и корь .
Международный комитет по таксономии вирусов (ICTV) классифицирует РНК -содержащие вирусы , как те , которые принадлежат к группе III , IV группы или группы V по классификации Балтимор системы классификации вирусов и не рассматривают вирусы с ДНК интермедиатами в их жизненном цикле , как РНК - вирусы. Вирусы с РНК в качестве своего генетического материала , которые также включают в себя промежуточные продукты ДНК в их цикле репликации называются ретровирусы , и включают в себя группу VI классификации Балтимор. Известный ретровирус человека включает в себя ВИЧ-1 и ВИЧ-2 , причина заболевания СПИДА .
Другой термин для РНК -содержащих вирусов , которые явно исключает ретровирусов ribovirus .
содержание
Характеристики
РНК - вирусы могут быть дополнительно классифицированы по смыслу или полярности их РНКА в отрицательную смысле и положительный смысл , или двусмысловые РНК - вирусы. Положительно смысл вирусной РНК аналогична мРНК и , таким образом , могут быть немедленно переведены клеткой - хозяином. Отрицательные чувства вирусной РНК является комплементарной мРНК и , следовательно , должны быть преобразованы в положительной смысловой РНК с помощью РНК-зависимой РНК - полимеразы перед переводом. Таким образом , очищенный РНК вируса позитивно-смысловой может непосредственно вызвать инфекцию , хотя это может быть менее заразным , чем весь вирусной частицы. Очищенная РНК вируса отрицательного смысла не заразна сам по себе , как это должно быть расшифровано в положительном смысловой РНК; каждый из вирионов может быть переписана несколько положительно смысловых РНК. Двусмысловые РНК - вирусы похожи на вирусы РНК отрицательного смысла, за исключением того, что они также переводят гены от положительной цепи.
В (DS) вирусы двунитевой РНК представляют собой разнородную группу вирусов , которые широко варьировать в диапазоне хозяина (человека, животных, растений, грибов и бактерий ), генома номера сегмента ( от одного до двенадцати) и вириона организации ( номер триангуляции , капсида слои, шипы, башни и т.д.). Члены этой группы включают ротавирусы , известный во всем мире как наиболее распространенной причиной гастроэнтерита у детей младшего возраста, а также picobirnaviruses , известные во всем мире , как наиболее часто встречающийся вирус в фекальных образцах как людей , так и животных , с или без признаков диареи. Вирус КЛО является экономически важным патогеном крупного рогатого скота и овец. В последние годы значительный прогресс был достигнут в определении, на атомных и subnanometeric уровнях структуры ряда ключевых вирусных белков и вириона капсид нескольких вирусов дсРНК, выделяя значительные параллели в структуре и репликативные процессы многих эти вирусы.
РНК - вирусы , как правило , имеют очень высокие мутационные цены по сравнению с ДНК - вирусами , так как вирусный РНК - полимераза не хватает корректуру способности ДНК - полимераз . Это одна из причин , почему это трудно сделать эффективные вакцины для профилактики заболеваний , вызываемых вирусами РНК. Ретровирусы также имеют высокую частоту мутаций , даже если их ДНК промежуточные интегрируют в геном хозяина (и, таким образом , при условии размещения ДНК корректуры один раз интегрированной), так как ошибки в процессе обратной транскрипции встроены в обе нити ДНК до интеграции. Некоторые гены РНК вируса являются важными для вирусных циклов репликации и мутация не допускаются. Например, область вируса гепатита С геном , который кодирует основной белок высоко консервативен , так как она содержит структуру РНК , участвующих в внутреннем сайте рибосомного входа .
копирование
РНК-вирусы животных классифицируются по ICTV. Есть три отдельные группы РНК-вирусов, в зависимости от их генома и способа репликации:
- Двухцепочечные РНК-вирусы (III группа) содержат от одного до дюжины различных молекул РНК, каждый из кодирования для одного или нескольких вирусных белков.
- Положительно смысловые оцРНК вирусы (группы IV) имеют их геном непосредственно использованы в качестве мРНК, с принимающими рибосомыперевод его в один белок , который модифицирован путем хозяина и вирусных белков , чтобы сформировать различные белки , необходимые для репликации. Один из них включает в себя РНК-зависимой РНК - полимеразы (РНК - репликазы), который копирует вирусной РНК с образованием двухцепочечной репликативной формы. В свою очередь , это дсРНК управляет формированием новой вирусной РНК.
- Отрицательно смысл оцРНК вирусы (группа V) должны иметь свой геном скопированный с помощью РНК-репликазы с образованием положительной смысловой РНК. Это означает, что вирус должен принести вместе с ним РНК репликазы фермента. Молекула РНК положительной смысл тогда действует как вирусной мРНК, которая транслируется в белки принимающими рибосом.
Ретровирусы (VI группы) имеют одноцепочечной РНК - геном , но, в общем, не считаются РНК -содержащих вирусов , поскольку они используют промежуточные продукты ДНК для репликации. Обратные транскриптазы , вирусный фермент , который происходит от вируса себя после того, как она без покрытия, преобразует вирусной РНК в комплементарные нити ДНК, которая копируется , чтобы произвести двухцепочечную молекулу вирусной ДНК. После того, как эта ДНК интегрирована в геном хозяина с помощью вирусного фермента интегразы , экспрессия генов , кодируемых может привести к образованию новых вирионов.
классификация
Классификация вирусов РНК, оказалось трудной задачей. Это отчасти из-за высокой частоты мутаций эти геномы подвергаются. Классификация основана главным образом, от типа генома (двухцепочечный, или отрицательный положительным однонитевой) и числа генов и организации. В настоящее время существует 5 порядков и 47 семейств РНК-содержащих вирусов, признаваемые. Есть также много нераспределенных видов и родов.
Относящиеся к но отличаются от РНК - вирусов являются вироидов и РНК спутниковые вирусы . Они в настоящее время не классифицируются как РНК - вирусы и описаны на своих страницах.
Изучение нескольких тысяч вирусов РНК показал наличие по крайней мере , пяти основных таксонов: в levivirus и группы родственников; пикорнавирус супергруппа; альфавирус супергруппа плюс флавивирус супергруппа; вирусы дцРНКа; и католита вирусы прядей. Лентивирусов группа , как представляется, базальная для всех остальных РНК -содержащих вирусов. Следующее крупное подразделение лежит между picornasupragroup и остальными вирусами. Вирусы дцРНК , по всей видимости, произошли от + ве РНК предка и минусу РНК -содержащих вирусов из вирусов внутри дцРНК. Ближайший reltion к минусу скрученных РНК вирусов являются Reoviridae .
Это единственная самая большая группа РНК-содержащих вирусов с 30 семей. Делались попытки сгруппировать эти семьи в высших порядках. Эти предложения были основаны на анализе РНК-полимеразы и все еще находятся на рассмотрении. На сегодняшний день предложение, предложенное не было широко принято из-за сомнения по поводу пригодности одного гена, чтобы определить таксономию кладов.
Предложенная классификация положительно прядей РНК-вирусов на основе РНК-зависимой РНК-полимеразы. Три группы были признаны:
- Bymoviruses, comoviruses, nepoviruses, nodaviruses, пикорнавирусы, potyviruses, sobemoviruses и подмножество luteoviruses (свекла вирус западных желтеет и вирус Скручивание листьев картофеля) -The пикорнавирусы, как группы (Picornavirata).
- Carmoviruses, dianthoviruses, флавивирусов, пестивирусов, statoviruses, tombusviruses, одноцепочечные РНК бактериофаги, вирус гепатита С и подмножество luteoviruses (желтой карликовости ячменя вирус) -The flavi, как группы (Flavivirata).
- Альфавирусы, carlaviruses, furoviruses, hordeiviruses, potexviruses, rubiviruses, tobraviruses, tricornaviruses, tymoviruses, яблоко хлоротичного вирус пятна листьев, свекла желтеет вирус и гепатит Е вирус-альфа, как группы (Rubivirata).
Альфа , как супергруппы может быть дополнительно разделен на три клады : рубите-подобный, tobamo-подобный, и tymo-подобные вирусы.
Дополнительная работа определила пять групп положительных одноцепочечной РНК вирусов, содержащих четыре, три, три, три, и один заказ (ы), соответственно. Эти четырнадцать порядков содержат 31 семейства вирусов (в том числе 17 семей вирусов растений) и 48 родов (в том числе 30 родов вирусов растений). Этот анализ предполагает, что альфавирусы и флавивирусы могут быть разделены на два семейство-в Togaviridae и Flaviridae, соответственно, но предполагают, что другие таксономические задания, такие как пестивирусы, вирус гепатита С, rubiviruses, вирус гепатита Е, и arteriviruses, могут быть неправильными. В коронавирус и toroviruses, как представляется, отдельные семьи в различных заказов и не отличаются роды того же семейства, в настоящее время классифицируются. В luteoviruses, как представляется, две семьи, а не один, а яблоко вирус хлоротичное пятнистость листьев, кажется, не быть клостеровирус, но новый род Potexviridae.
Эволюция пикорнавирусах на основе анализа их РНК - полимеразы и геликаз , как представляется , дата дивергенции эукариот . Их предполагаемые предки включают бактериальную группа II ретроэлементы , семейство HtrA протеаз и ДНК бактериофагов .
Partitiviruses связаны и, возможно, произошли от totivirus предка.
Hypoviruses и barnaviruses по-видимому, разделяют родословную с potyvirus и sobemovirus линий соответственно.
Этот анализ также показывает, что вирусы дцРНКа не тесно связаны друг с другом, но вместо того, чтобы принадлежать к четырем дополнительным классам-Birnaviridae, Cystoviridae, Partitiviridae и Reoviridae - и один дополнительному заказу (Totiviridae) одного из классов положительных вирусов оцРНКа в же подтип, как положительно прядей РНК-вирусов.
Одно исследование предположило, что есть два больших клада: один включает в Caliciviridae, Flaviviridae и Пикорнавирусов семьи и второй, который включает в себя Alphatetraviridae, Birnaviridae и Cystoviridae, Nodaviridae и Permutotretraviridae семьи.
Эти вирусы имеют различные типы генома , начиная от одной молекулы РНК до восьми сегментов. Несмотря на их разнообразие кажется , что они , возможно, возникли в членистоногих и диверсифицируют оттуда.
Ряд спутниковых вирусов - вирусы , которые требуют помощи другого вируса , чтобы завершить свой жизненный цикл - также известны. Их таксономии выделялось еще предстоит урегулировать. Следующие четыре рода были предложены для положительных чувственных одноцепочечных РНК спутниковых вирусов , которые заражают растения - Albetovirus , Aumaivirus , Papanivirus и Virtovirus . Семейство - Sarthroviridae , который включает в себя род Macronovirus - был предложен для положительном смысле одноцепочечных РНК спутниковых вирусов, поражающих членистоногих .
Вирусы группы III-дцРНК
Есть двенадцать семей и ряд присвоенных в родах и видов, признанных в этой группе.
Группа IV-позитивно-смысловые вирусы оцРНК
Есть три порядка и 34 семей, признанных в этой группе. Кроме того, существует целый ряд неклассифицированных видов и родов.
Группа V-отрицательные чувства вирусы оцРНКа
С Exeption от вируса гепатита D , эта группа вирусов были помещены в один фила - Negarnaviricota . Этот тип был разделен на две subphyla - Haploviricotina и Polyploviricotina . В подтипа Haploviricotina четыре класса в настоящее время признаются: Chunqiuviricetes , Milneviricetes , Monjiviricetes и Yunchangviricetes . В подтипа Polyploviricotina два класса признаны: Ellioviricetes и Insthoviricetes .
Шесть классов, семь порядков и двадцать четыре семьи в настоящее время признаются в этой группе. Ряд присвоенных в видов и родов еще не классифицированы.
Читайте также: