
Размножению вирусов инфекций патогенной
В основе патогенеза вирусных инфекций лежит взаимодействие генома вируса с генетическим аппаратом чувствительной клетки. Исключая поражения, вызываемые вирусами, распространяющимися по нервной ткани, патогенез вирусных инфекций сопровождает вирусемия (виремия)— циркуляция возбудителя в крови. В кровоток возбудитель проникает прямым путем или из лимфатической системы. Многие вирусы (например, ВИЧ, вирусы гриппа, кори, герпеса) поражают иммунокомпетентные клетки, что чаще проявляется в нарушении функций и уменьшении числа Т-хелперов, увеличении содержания и активации Т-супрессоров или В-клеток. Некоторые возбудители образуют внутриядерные или цитоплазматические тельца включений (например, тельца Бабеша
Нёгри). имеющие диагностическое значение.
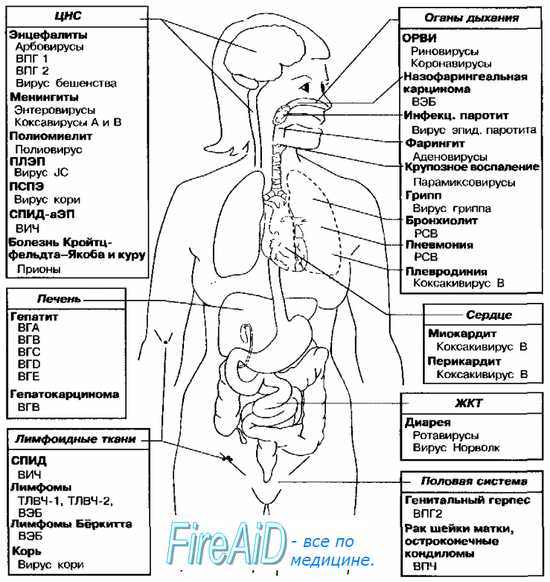
Рис. 5-9. Органы-мишени наиболее распространённых вирусных инфекций человека. ВЭБ — вирус Эпстайна-Барр, ВГА — вирус гепатита А, ВГВ — вирус гепатита В, ВГС — вирус гепатита С, BID — вирус гепатита D, ВГЕ — вирус гепатита Е, ВПЧ — вирус папилломы человека, ВПГ — вирус простого герпеса, ТЛВЧ — Т-пимфотропный вирус человека, ВИЧ — вирус иммунодефицита человека, ПЛЭП — прогрессирующая лейкоэнцефалопатия, РСВ — респираторно-синцитиальный вирус, ПСПЭ — подострый склерозирующий панэнцефалит, ОРВИ — острая респираторная вирусная инфекция, СПИД-аЭП — СПИД-ассоциированная энцефалопатия.
Проникновение вируса в организм. Основные входные ворота для возбудителей вирусных инфекции человека — дыхательные пути и ЖКТ, реже — кожные покровы. В некоторых случаях развиваются локальные поражения, но чаще в месте проникновения не возникает каких-либо проявлений или они носят стёртый характер, а возбудитель мигрирует в чувствительные ткани. Распространение возбудителя в организме может носить локальный или системный характер.
Локальные поражения вирусами типичны для возбудителей респираторных и кишечных инфекций, а также для некоторых кожных заболеваний. Продолжительность инкубационного периода большинства подобных инфекций составляет 2-3 сут. Первичную репликацию часто сопровождает вирусемия. Она обычно протекает бессимптомно или по типу продромальных явлений, но может возникать и на фоне выраженной клинической картины, не вызывая развития дополнительной симптоматики. Для подобных заболеваний характерно повторное заражение, так как циркулирующие AT не проявляют протективный эффект, а секреторный иммуноглобулин А (IgA) оказывает лишь кратковременное нейтрализующее действие на слизистой оболочке. Системные поражения. Из места проникновения возбудители попадают в кровоток, вызывая вирусемию, и постепенно фиксируются в чувствительных тканях. Первичное распространение обычно вызывает продромальные явления. Поскольку вирусемия предшествует поражению чувствительных тканей, то продолжительность инкубационного и продромального периодов подобных инфекций могут увеличиваться до 2-3 нед. Вирусемия при системных инфекциях обычно носит двухэтапный характер. Первый этап заканчивается поглощением циркулирующих вирусов клетками ретикулоэндотелиальной системы.
В дальнейшем возможно несколько вариантов:
• полная элиминация возбудителя (абортивная инфекция);
• размножение вирусов в фагоцитах с последующим выходом и развитием выраженной вторичной вирусемии, сопровождающейся появлением характерных клинических признаков заболевания (например, энцефалитов);
• некоторые вирусы (например, вирус гепатита В, пикорна- и тогавирусы) слабо поглощаются фагоцитами и могут циркулировать в крови в свободном состоянии, а возбудители колорадской клещевой лихорадки и лихорадки долины Рифт внедряются в эритроциты.
Основные органы-мишени наиболее распространённых вирусных инфекций представлены на рис. 5-9. Многие из указанных на рисунке возбудителей могут поражать, кроме названных, и другие ткани (так, полиовирусы способны вызывать поражения ЖКТ, а вирус эпидемического паротита обладает тропностью к эпителию извитых канальцев яичек).
Патогенные свойства вирусов складываются из следующих компонентов:
1.способности вируса проникать в организм и адсорбироваться на клеточных мембранах,
2.проникать в чувствительные к ним клетки;
3.способности этих клеток депротеинизировать вирусный геном и делать его функционально активным;
4.пермиссивности клеток или возможности этих клеток обеспечить транскрипцию и репликацию генетического материала, полноценную сборку вирионов;
5.возможности воспроизведения в клетках нескольких циклов репродукции вирусов, цитопатического действия вируса;
6.способности вирусов распространяться на новые клетки, расположенные рядом с пораженными; распространения вирусов за пределы первичного очага поражения по всему организму;
7.способности вызывать местные и общие патологические цроцессы, лежащие в основе клинических проявлений вызываемых ими заболеваний;
8.способности вйируса к переходу в новый организм и обеспечение его эстафетной передачи.
Все эти свойства необходимы, но в то же время сами по себе они могут быть недостаточными для патогенного действия вируса. Некоторые из этих свойств обусловлены клетками, в которых они размножаются, что получило название хозяйского ограничения клеткой.
Многие вирусы проникают в организм непосредственно через слизистые оболочки, которые служат входными воротами инфекции и защищены целым рядом неспецифических факторов резистентности, поэтому вирусы должны быть устойчивы к действию данных неблагоприятных факторов, что детерминируется генами вирусов. Например, кишечные вирусы обычно устойчивы к кислым значениям рН, детергентному действию солей желчных кислот и к разрушающему их действию протеолитических ферментов.
Способность вирусов адсорбироваться на мембранах чувствительных к вирусам клеток является специфическим процессом для вирусов. Этот процесс протекает при участии прикрепительных белков (антирецепторов) у вирусов и чувствительных к ним клеточных рецепторов. Простые вирусы содержат прикрепительные белки в составе капсида, а сложноустроенные вирусы — в составе супер-капсида. Такие сложные вирусы, как вирус осповакцины и вирус простого герпеса, могут иметь прикрепительные белки нескольких видов. Способность вирусов адаптироваться к новому хозяину обусловлена изменением первичной структуры в области участка прикрепительного белка, узнающего клеточный рецептор. Эти участки консервативны по своему строению и расположены в углублениях-каньонах, которые чрезвычайно малы по своим размерам, благодаря чему недоступны для активных центров антител, реагирующих лишь с окружающими эти углубления гипервариабельными участками, что позволяет вирусам избежать иммунологического пресса. Мутации в генах, кодирующих антирецепторы, иногда приводят к полной потере способности вирусов взаимодействовать с клеточными рецепторами.
В отличие от парамиксовирусов, у вирусов гриппа белком слияния является гемагглютинин, обуславливающий также адсорбцию вирусов к клетке. Однако функции прикрепления и слияния разделены между разными его участками большой (НА1) и малой (НА2) субъединицами соответственно. Важным фактором патогенности у вирусов гриппа является нейраминидаза, которая, удаляя остатки сиаловой кислоты с вирусного гемагглютинина, делает его доступным для протеолитического расщепления, необходимого для проявления инфекционности вирусов.
Очевидно, что сходный по функции с белками слияния сложных вирусов белок существует в составе капсида простых вирусов, и один из поверхностно расположенных белков капсида вызывает дестабилизацию клеточной мембраны, что способствует проникновению модифицированного капсида из эндоцитарной вакуоли в цитоплазму.
Взаимодействие вируса и клетки — это всегда взаимодействие вирусного и клеточного генома. В результате адсорбции вируса, его проникновения в клетку и раздевания происходит освобождение генетического материала вирусов, который становится функционально активным, так как освобождается от внешних защитных оболочек, препятствующих его экспрессии. Степень активности генома обусловлена разной степенью депротеинизации у вирусов разных семейств. Депротеинизация
клетки приспособлен для трансляции только моноцистронных мРНК, так как он не распознает внутренних участков инициации в мРНК. В результате вирусы вынуждены синтезировать либо отдельные мРНК для каждого гена, либо мРНК, включающие несколько генов и кодирующие большой полипротеин, который затем разрезается на индивидуальные белки. Транскрипция вирусного генома строго регулируется на протяжении инфекционного процесса многочисленными вирусо-специфическими и клеточными факторами. Со степенью транскрипции нередко связан характер инфекции, ее тип (от продуктивной до абортивной инфекции).
Важную роль в регуляции процессов транскрипции играют гены усилители и трансактиваторы. Они расположены в специальной области генома вирусов и содержат гены, усиливающие и активирующие экспрессию структурных генов. Усилители— это генетические элементы, усиливающие транскрипцию. Структура вирусных усилителей не отличается от структуры клеточных. Факторы транскрипции, связывающиеся с промотором и усилителем, выполняют одну и ту же функцию и могут представлять собой как клеточные, так и вирусные белки. Усилители, контролирующие уровень экспрессии генов, обнаружены у паповавирусов, гепаднавиру-сов, герпесвирусов, ретровирусов и ряда других вирусов.
Белки трансактиваторыне обладают специфичностью действия. Они связываются с регуляторными областями генов и одновременно активируют усиленную транскрипцию всех генов, в том числе и других вирусов, что сопровождается взрывной продукцией вирусных частиц, а также включают экспрессию бактериальных генов и клеточных онкогенов. Они действуют не только на стадии транскрипции, но и на посттранскрипционном уровне. Взаимодействие вирусных и клеточных трансактиваторов может приводить к переходу латентной инфекции в литическую, а также к онкогенной трансформации зараженных клеток. Как и усилители, трансактиваторы содержат две важные для их функции области. Одна из них определяет транспорт и связывание белка с мишенью, а другая пред-
Усилители и трансактиваторы являются необходимым атрибутом вирусов как генетических паразитов, конкурирующих с клеточным геномом. Неравные шансы небольших по размерам вирусов на победу уравновешиваются возникшими в ходе эволюции генетическими элементами, позволяющими гораздо меньшей по величине молекуле вирусного генома успешно завершить экспрессию своих генов и создать вирусное потомство. При этом вирусы широко используют механизмы клеточного происхождения, которые теперь обращены против клетки хозяина.
Важную роль в формировании патогенное -ти сложных вирусов, помимо посттрансляционной модификации вирусных белков,.играет синтез М-белка (матриксного белка), участвующего в сборке вирусной частицы. Включение М-белка в плазматическую мембрану является лимитирующим событием, определяющим возможность почкования вирусных частиц. Синтез М-белка жестко регулируется как ви-русоспецифическими, так и клеточными механизмами. Количество М-белка в зараженных клетках во многом определяет особенности репродукции вируса в данной клеточной системе. Аберрантный синтез М-белка и его нарушенный внутриклеточный транспорт служат одной из частых причин абортивных и персис-тентных вирусных инфекций. Экспрессия гена М значительно варьирует в клетках разного происхождения.
Патогенность вирусов обусловлена также их белковыми продуктами, блокирующими апоптоз клетки и изменяющими защитные реакции в макроорганизме, подавляя продукцию цитокинов, что способствует репродукции вирусов и их распространению по макроорганизму. Например, вирусы натуральной оспы образуют TNF-связывающий белок, белки, подавляющие созревание антигенов МНС 1 класса и аналог рецепторов у-интерферона. Вирус иммунодефицита человека, наоборот, усиливает продукцию цитокинов пораженными им клетками, что ведет к усилению воспалительной реакции и развитию нейротоксического действия. Как и другие микробы, вирусы, благодаря наличию внешней липидсодержащей оболочке, образованной из мембраны клетки хозяина, вариабельности структуры поверхностных антигенов, интеграции в геном клетки, гибели Т-лимфоцитов и т. д., обладают способностью уходить от воздействия иммунной системы макроорганизма.
Заражение восприимчивых клеток вовсе не означает, что в клетках неизбежно будет происходить размножение вируса, так как восприимчивость не идентична пермиссивнос-ти клеточной системы. Это одна из главных концепций в вирусологии. Многие стадии взаимодействия вируса с клеткой имеют не столько вирусоспецифическую, сколько опосредованную клеткой природу (эндоцитоз, де-протеинизация, синтез вирусоспецифических белков и т.д.). Клетка принимает активное участие в формировании патогенных вирусов лишь в пермиссивной клеточной системе, содержащей весь набор необходимых факторов, используемых вирусами на разных стадиях инфекционного процесса, а репликативный цикл завершается и приводит к образованию инфекционного потомства, что не будет происходить в полупермиссивных и непермис-сивных клеточных системах (хозяинная или хозяйская рестрикция).
Тканевой тропизм определяется не только наличием на клетках рецепторов, но и возможностью осуществления в клетках ви-русоспецифических синтезов. В зависимости от пермиссивности клеточной системы инфекция восприимчивых клеток может быть продуктивной, ограниченной и абортивной.Продуктивная инфекция происходит в пер-миссивных клетках и характеризуется полным циклом репродукции, который заканчивается формированием инфекционного потомства. Пермиссивность клеточной системы обуславливает и многократную цикличность размножения в ней вирусов.
Абортивной называется инфекция, которая не завершается образованием инфекционных вирусных частиц или при которой они образуются в гораздо меньшем количестве, чем при продуктивной инфекции. Абортивная инфекция может наступить в силу двух обстоятельств. Во-первых, несмотря на восприимчивость к заражению, клетки могут оказаться непермиссивными, так как в них могут экспрессироватся не все, а лишь некоторые гены вирусов. В основе механизмов генетически обусловленной непермиссивнос-ти клеток лежит либо отсутствие клеточных факторов, необходимых для репродукции, либо наличие факторов, нарушающих процессы репродукции вирусов. Во-вторых, абортивная инфекция может быть результатом заражения как пермиссивных, так и непермиссив-ных клеток дефектными вирусами,у которых отсутствует полный набор вирусных генов, необходимых для репродукции. Дефектные вирусы представляют собой крайнюю форму паразитизма, так как они используют генные продукты, образованные другими, часто не родственными им, не гомологичными вирусами. Примером таких вирусов являются аденоассоциированные вирусы и вирус гепатита D, помощником которого служит вирус гепатита В. Абортивную инфекцию вызывают также дефектные интерферирующие вирусные
частицы,которые тоже лишены части генетического материала. В отличие от дефектных вирусов, в ходе репликации они интерферируют с гомологичными инфекционными вирусами, в связи с чем их назвали дефектными интерферирующими вирусными частицами (ДИ-частицами). Образование ДИ-частиц играет важную роль в ослаблении летального действия полноценных вирусов в силу интерференции и предрасполагает некоторые клетки к формированию в них длительной персистентной инфекции.
Наконец, клетки могут быть только временно пермиссивными, вследствие чего вирус либо сохраняется в клетках до момента, когда они становятся пермиссивными, либо в любой данный момент вирусное потомство образуется только в немногих клетках популяции. Этот вид инфекции одними исследователями был определен как рестриктив-ный (restrictive), другими — как ограниченный (restringent). В ряде случаев цитолитические вирусы могут только лишь изменять функциональную активность клеток, не вызывая их морфологических повреждений (изменять синтез гормонов, холестерина и т. д.), или вызывать опухолевую трансформацию клеток. Дополнительным следствием как ограниченной, так и абортивной инфекции является сохранение в клетке вирусного генома.
Если геном вируса реплицируется независимо от клеточного генома, такая инфекция называется автономной. Если вирусный геном интегрирует в состав генома клетки и реплицируется вместе с ним, то такая инфекция называется интегративной (вирогения). Интегрировать может как полный геном, так и часть его. Например, при гепатите В возможна интеграция полного генома, при аденовирусной или герпесвирусной инфекциях обычно интегрирует часть генома, при заражении онковирусами может интегрировать как полный геном, так и часть его. Вирусные последовательности, входящие в состав генома клетки, называются провирусом или про-вирусной ДНК. Интеграционный тип инфекционного процесса возможен при заражении адено-, папиллома-, герпесвирусами, вирусом гепатита В и обязателен для ретровирусов, имеющих фермент — обратную транскриптазу. Возникшая интеграция может явиться причиной ряда хронических и автоиммунных заболеваний.
По исходу взаимодействия с клеткойинфекция может быть
Патогены представляют собой микроскопические организмы, которые вызывают или могут вызывать заболевание. Различные виды возбудителей инфекций включают бактерии, вирусы, протисты (амеба, плазмодий и т. д.), грибы, паразитические черви (плоские черви и круглые черви ) и прионы. Хотя эти патогены вызывают множество заболеваний, начиная от незначительных до опасных для жизни, важно отметить, что не все бактерии являются патогенными.
Фактически, наш организм содержит тысячи видов бактерий, грибов и простейших, которые являются неотъемлемой частью микробиома (микрофлоры) человека. Эти микроорганизмы полезны и важны для правильной работы биологических процессов, таких как пищеварение и функция иммунной системы. Они вызывают проблемы только в редких случаях, когда ослабевают функции иммунной системы. Напротив, действительно патогенные организмы имеют одну цель: выжить и размножаться любой ценой. Возбудители инфекций специально приспособлены для заражения живых организмов в обход иммунной системы хозяина. Они распространяются внутри тела и покидают его для заражения другого хозяина.
Как передаются патогены?
Патогены могут передаваться либо прямым, либо косвенным путем. Прямая передача включает распространение патогенов путем прямого контакта тела с телом. Такой тип передачи может происходить от матери к ребенку, как показано на примере ВИЧ, вируса Зика и сифилиса. Другие типы прямого контакта, через которые могут распространяться патогены, включают касание (метициллин-резистентный стафилококк), поцелуи (простой герпес) и сексуальный контакт (папилломавирусы человека).
Патогены способны также распространяться путем косвенной передачи, которая включает контакт с поверхностью или веществом, которые заражены вредоносными микроорганизмами, а также контакт и передача через животное или укс насекомых. Типы косвенной передачи включают:
- Воздушно-капельный (как правило, чиханьем, кашлем, смехом и т. д.). Вредоносные микроорганизмы остаются подвешенным в воздухе и вдыхается или контактирует с дыхательными мембранами другого человека.
- Капельки - патогены, содержащиеся в каплях жидкости организма (слюна, кровь и т. д.) контактируют с другим человеком или загрязняют поверхность. Капли слюны чаще всего распространяются через чиханье или кашель.
- Еда - передача инфекции происходит путем употребления зараженной и неправильно обработанной пищи.
- Вода - патоген распространяется через потребление или контакт с загрязненной водой.
- Животные - патоген распространяется от животных к людям. К примеру, при укусе насекомых или контакте людей с дикими или домашними животни.
Несмотря на то, что невозможно полностью предотвратить передачу патогенов, лучший способ минимизировать вероятность инфекционных заболеванием - поддерживать правильную гигиену. Не забывайте мыть руки после использования туалета, обрабатывайте сырые продукты и различные поверхности, которые подвергаются воздействию микробов, а также своевременно убирайте экскременты домашних питомцев.
Типы патогенов
Патогены очень разнообразны и состоят из прокариотических и эукариотических организмов. Наиболее распространенными патогенами являются бактерии и вирусы. Хотя оба способны вызвать инфекционное заболевание, бактерии и вирусы отличаются друг от друга. Бактерии являются прокариотическими клетками, которые вызывают заболевание, продуцируя токсины. Вирусы представляют собой частицы нуклеиновой кислоты (ДНК или РНК), заключенные внутри оболочки белка или капсида. Они вызывают заболевание, захватывая клеточную систему своего хозяина, чтобы создать многочисленные копии вируса. Эта активность уничтожает клетку-хозяина в процессе. Эукариотические возбудители инфекций включают грибы, протистов и паразитических червей.
Прионы - это уникальный тип патогена, который является белком, а не живым организмом. Белки прионов имеют те же аминокислотные последовательности, что и обычные белки, но сложены в неправильную форму. Эта измененная форма делает прионные белки заразными, поскольку они влияют на другие нормальные белки, заставляя спонтанно принимать инфекционную форму. Прионы обычно влияют на центральную нервную систему. Они, как правило, собираются вместе в тканях мозга, что приводит к ухудшению состояния мозга. Прионы вызывают смертельное нейродегенеративное заболевание Крейтцфельдта-Якоба у людей, а также губчатую энцефалопатию у крупного рогатого скота.
Бактерии ответственны за ряд инфекций, которые варьируются от бессимптомных до внезапных и интенсивных. Заболевания, вызванные патогенными бактериями, обычно являются результатом производства токсинов. Эндотоксины служат компонентами стенки бактериальных клеток, высвобождающиеся после смерти или ухудшения состояния бактерий. Эти токсины вызывают различные симптомы, включая лихорадку, изменения артериального давления, озноб, септический шок, повреждение органов и даже смерть.
Экзотоксины продуцируются бактериями и высвобождаются в окружающую среду. Три типа экзотоксинов включают цитотоксины, нейротоксины и энтеротоксины. Цитотоксины повреждают или уничтожают определенные типы клеток организма. Бактерии Streptococcus pyogenes продуцируют цитотоксины, называемые эритротоксинами, которые разрушают клетки крови, повреждают капилляры и вызывают симптомы, связанные с некротическим фасциитом.
Нейротоксины являются ядовитыми веществами, которые действуют на нервную систему и мозг. Бактерии Clostridium botulinum высвобождают нейротоксин, вызывающий мышечный паралич. Энтеротоксины влияют на клетки кишечника, провоцируя сильную рвоту и диарею. Бактериальные виды, которые производят энтеротоксины, включают Bacillus, Clostridium, Escherichia, Staphylococcus и Vibrio.
- Clostridium botulinum: отравление ботулизмом, затрудненное дыхание, паралич;
- Streptococcus pneumoniae: пневмония, ангина, менингит;
Вирусы - это уникальные патогены, поскольку они не являются клетками, а сегментами ДНК или РНК, заключенными внутри капсида (белковая оболочка). Они вызывают заболевание, заражая клетки и заставляя клеточные структуры вырабатывать больше вирусов быстрыми темпами. Вирусы противодействуют или предотвращают обнаружение иммунной системой и энергично размножают внутри клетки-хозяина. Эти микроскопические вредоносные частички инфицируют не только клетки животных и растений, но также заражают бактерии и археи.
Вирусные инфекции у людей варьируются по тяжести от умеренных до смертельно опасных (Эбола). Они часто мигрируют и заражают определенные ткани или органы в организме. Вирус гриппа имеет родство с тканям дыхательной системы, что приводит к симптомам, затрудняющим дыхание. Вирус бешенства обычно заражает ткани центральной нервной системы, а различные вирусы гепатита локализируются в печени. Некоторые вирусы также связаны с развитием определенных типов рака. Вирусы папилломы человека связаны с раком шейки матки, гепатит В и С провоцируют возникновения рака печени, а вирус Эпштейна-Барра связан с лимфомой Беркитта.
- Вирус Эбола: геморрагическая лихорадка Эбола;
- Вирус иммунодефицита человека (ВИЧ): пневмония, фарингит, менингит;
- Вирус гриппа: грипп, вирусная пневмония;
- Норовирус: вирусный гастроэнтерит (желудочный грипп);
- Вирус ветряной оспы: ветряная оспа (ветрянка);
- Вирус Зика: вирусная болезнь Зика, микроцефалия (у младенцев).
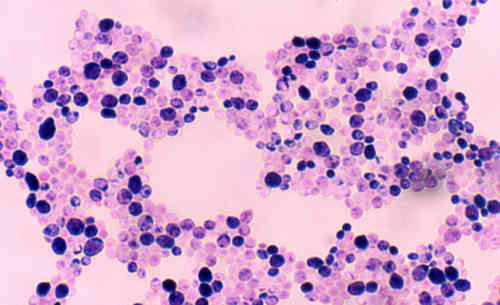
Грибы - эукариотические организмы, которые включают дрожжи и плесень. Заболевание, вызванное грибами, редко встречается у людей и обычно является результатом повреждения физического барьера (кожи, слизистой оболочки и т.д.) или нарушений в работе иммунной системы. Патогенные грибы часто вызывают заболевание, переходя от одной формы роста к другой. То есть, одноклеточные дрожжи демонстрируют обратимый рост от дрожжеподобной до плесневой формы, тогда как плесень переходит к дрожжеподобному росту.
Дрожжи Candida albicans меняют морфологию, переключаясь с округлого возрастающего роста клеток на плетевидный (нитевидный) удлиненный клеточный рост, основанный на ряде факторов. Эти факторы включают изменения температуры тела, рН и наличия определенных гормонов. C. albicans вызывает вагинальные дрожжевые инфекции. Подобным же образом гриб Histoplasma capsulatum существует как нитчатая плесень в естественной среде обитания почвы, но при попадании в организм переключается на почкообразный рост дрожжей. Импульсом для этого изменения является повышенная температура в легких по сравнению с температурой почвы. H. capsulatum вызывает тип инфекции легких, называемый гистоплазмозом, который может развиться в легочные заболевания.
- Aspergillus spp.: бронхиальная астма, аспергиллез легких;
- Candida albicans: оральный кандидоз, вагинальные дрожжевые инфекции;
- Epidermophyton spp.: атлетическая стопа, стригущий лишай;
- Histoplasma capsulatum: гистоплазмоз, пневмония;
- Trichophyton spp.: заболевания кожи, волос и ногтей.
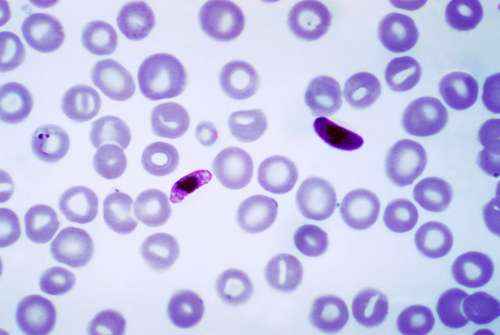
Простейшие - крошечные одноклеточные организмы из царства протисты. Это очень разнообразное царство включает в себя такие организмы, как водоросли, амебы, эвглена, плазмодий и слизевики. Большинство протистов, которые вызывают заболевание у людей, являются простейшими. Они делают это путем паразитного питания и размножения за счет хозяина. Паразитарные простейшие обычно передаются людям через зараженную почву, пищу или воду. Они также могут передаваться животными и через укусы насекомых.
Амеба Naegleria fowleri, обычно встречающаяся в почвенных и пресноводных местах обитания, также называется мозговой амебой, так как вызывает заболевание, называемое первичным амебным менингоэнцефалитом (ПАМ). Эта редкая инфекция обычно возникает, когда люди купаются в зараженной воде. Амеба мигрирует из носа в мозг, где повреждает ткани мозга.
- Giardia lamblia: лямблиоз (диарейные заболевания);
- Entamoeba histolytica: амебная дизентерия, амебный абсцесс печени;
- Plasmodium spp.: малярия;
- Trypanosoma brucei: африканская сонная болезнь;
- Trichomonas vaginalis: трихомониаз (инфекция, передающаяся половым путем);
- Toxoplasma gondii: токсоплазмоз, биполярное расстройство, депрессия, болезни глаз.

Эти паразиты заражают ряд различных организмов, включая растения, насекомых и животных. Паразитические черви, также называемые гельминтами, включают нематоды (круглые черви) и платигельминт (плоские черви). Анкилостомы, острицы, нитевидные черви, власоглав и трихинеллы являются типами паразитических круглых червей. Паразитические плоские черви включают ленточных червей и трематод. У людей большинство этих червей заражают кишечник, но иногда распространяются на другие участки тела. Кишечные паразиты прикрепляются к стенкам пищеварительного тракта и питаются за счет хозяина. Они производят тысячи яиц, которые вылупляются либо внутри, либо снаружи тела.
Паразитические черви распространяются через контакт с загрязненной пищей и водой. Они также могут передаваться от животных и насекомых людям. Не все паразитические черви заражают пищеварительный тракт. Плоские черви - шистосомы могут проникать в органы тела (печень, селезенка, легкие), вызывая потерю крови, обструкцию толстой кишки, увеличение селезенки или чрезмерное накопление жидкости в брюшной полости. После того, как самки откладывают яйца, некоторое количество яиц выходит из организма с мочой или фекалиями. Шистосомы передаются при контакте с зараженной личинками водой. Они попадают в организм, проникая через кожу.

Вадим Израилевич Агол — член-корреспондент РАН и РАМН, доктор биологических наук, главный научный сотрудник Института полиомиелита и вирусных энцефалитов им. М. П. Чумакова, заведующий отделом Научно-исследовательского института физико-химической биологии им. А. Н. Белозерского МГУ. Область научных интересов — молекулярная биология и генетика вирусов.
Считается, что вирусы (от лат. virus — яд) — нечто противное, приносящее одни неприятности. Но это грубая ошибка. Вирусы — ключевые созидатели живой природы и двигатели ее эволюции.
Каковы главные защитные механизмы зараженной клетки? Это компоненты врожденного иммунитета: деградация РНК (вирусных, а также клеточных), угнетение синтеза белков (как вирусных, так и клеточных), самоликвидация (апоптоз и другие виды программируемой гибели) и, наконец, воспаление. Собственно, многие вирусы так и обнаружили свое существование — из-за вызываемого ими воспаления (энцефалита, воспаления легких и т. д.). Клетка борется с вирусом, нарушая собственные обмен веществ и / или структуру, и ее защитные механизмы, как правило, самоповреждающие. Можно сказать, что человек, умерший от полиомиелита (а умирает менее 1%), сам убил себя, борясь с инфекцией.
У растений в качестве противовирусного механизма очень важную роль играет РНК-интерференция. Из вирусной РНК образуется двуцепочечная (важный фактор, по которому клетка узнает о наличии вируса). При участии компонентов системы РНК-интерференции — фермента Dicer, который разрезает эту двуцепочечную РНК на фрагменты длиной 21–25 пар нуклеотидов, а затем РНК-белкового комплекса RISC — в конце концов образуются одноцепочечные короткие фрагменты РНК. Гибридизуясь с вирусной РНК, они вызывают либо ее деградацию, либо угнетение ее трансляции. Такой защитный механизм эффективен, но может повреждать саму клетку, что хорошо видно на примере вироидов. Это патогены растений, короткие (несколько сотен нуклеотидов) молекулы кольцевой одноцепочечной РНК, не покрытые белковой оболочкой. Вироиды не кодируют белки, но могут вызывать тяжелые симптомы в зараженном растении. Это происходит потому, что клетка защищается. Образующаяся вироидная двуцепочечная РНК подвергается действию всех компонентов системы РНК-интерференции, в результате образуются фрагменты одноцепочечной РНК, которые гибридизуются уже не с вирусной РНК, а с клеточной. Это приводит к ее деградации и развитию симптомов заболевания. Однако многие вирусы растений кодируют разнообразные белки, препятствующие РНК-интерференции (viral suppressors of RNA silencing — VSR). Они либо угнетают распознавание и расщепление вирусных РНК, либо подавляют формирование и функционирование комплекса RISC. Поэтому эти VSR-белки могут нарушать механизмы физиологически важной (не связанной с вирусами) РНК-интерференции, вызывая патологические симптомы.


Схема полипротеина (белка-предшественника) пикорнавирусов

Модель вируса полиомиелита. Фото: virology.wisc.edu

Модель менговируса (штамма вируса энцефаломиокардита). Фото: virology.wisc.edu

Эффект одновременного выключения клеточных защитных и вирусных противозащитных механизмов. Зараженные менговирусом дикого типа, клетки HeLa (а, незараженные) быстро гибнут от некроза (б, 4 ч после заражения). Клетки, зараженные частично разоруженным вирусом, у которого инактивирован лидерный белок, живут чуть дольше и гибнут уже от апоптоза (в, 8 ч после заражения). Когда же частично разоружены и вирус (инактивирован его лидерный белок), и клетки (у них выключен апоптоз добавлением химического соединения, которое угнетает каспазы), даже через вдвое больший промежуток времени клетки чувствуют себя значительно лучше, чем те, которые не были разоружены (г, д, 8 и 16 ч после заражения). Большинство клеток на панелях г и д морфологически больше похожи на клетки на панели а, чем на б и в. Сканирующая электронная микроскопия. Фото С. И. Галкиной
Однако длительная коэволюция хозяина и вируса должна приводить к снижению патогенности последнего (взаимовыгодному обоюдному разоружению). Классический пример — вирус миксомы / фибромы. В середине XIX в. в Австралию завезли европейских кроликов, которые быстро размножились и стали серьезной угрозой для сельского хозяйства. Через 100 лет для контроля их популяции стали использовать патогенный вирус фибромы / миксомы (из семейства поксвирусов, к которому относится и вирус оспы). Разные кролики по-разному реагируют на этот вирус. У бразильских кроликов через три недели после заражения он вызывает доброкачественную опухоль — фиброму (локализованный узелок на коже). Но у европейских кроликов, чувствительных к этому вирусу, уже через 10 дней после заражения развивается генерализованное смертельное заболевание.
Мой рассказ далеко не исчерпывает тему: о природе патогенности вирусов известно значительно больше. Многое из того, что мы сейчас знаем, удалось изучить в самые последние годы, и есть все основания ожидать новых сюрпризов. Можно и нужно винить вирусы за тяжелые болезни и необходимо бороться с ними, но мы должны быть благодарны вирусам за существование и разнообразие живой природы, и в том числе — за существование человека.
Автор благодарен коллегам по научной кооперации — сотрудникам Института полиомиелита и вирусных энцефалитов им. М. П. Чумакова РАМН, Московского государственного университета им. М. В. Ломоносова, Института белка РАН (Пущино Московской области), Университета Базеля (Швейцария), Университета штата Висконсин (США), Университета Неймегена им. Радбода (Нидерланды).
Читайте также: