
Способ размножения вирусов дизъюнктивный
Размножение вирусов не осуществляется бинарным делением. Еще в 50-х годах прошлого века было установлено, что размножение осуществляется методом репродукции (в переводе с англ. reproduce – делать копию, воспроизводить), то есть путем воспроизведения нуклеиновых кислот, а также синтеза белка с последующим сбором вирионов. Данные процессы происходят в различных частях клетки так называемого хозяина (к примеру, в ядре или цитоплазме). Данный разобщенный метод репродукции вирусов называется дизъюнктивным. Именно на этом мы и остановимся подробнее в нашей статье.

Процесс репродукции
Данный процесс имеет свои особенности репродукции вирусов и отличается последовательной сменой некоторых стадий. Рассмотрим их по отдельности.
Вирусы не могут размножаться в питательной среде, так как они представляют собой строгие внутриклеточные паразиты. Кроме того, в отличие от хламидий или риккетсий, во время репродукции вирусы в клетке хозяина не способны расти и не размножаются методом деления. Все составные части данного вируса включают в себя нуклеиновые кислоты, а также белковые молекулы, которые синтезируются в "хозяйской" клетке раздельно, в различных частях клетки: в цитоплазме и в ядре. Помимо этого, белоксинтезирующие клеточные системы подчиняются одному вирусному геному, а также его НК.

Вирусная репродукция в клетке осуществляется в несколько фаз, которые описаны ниже:
Адсорбция
Под этой стадией репродукции вирусов подразумевается проникновение вирусной частицы в клетки. Адсорбция начинается на клеточной поверхности при помощи взаимодействия клеточных, а также вирусных рецепторов. В переводе с латинского слово "рецепторы" означает "принимающий". Они представляют собой специальные чувствительные образования, которые воспринимают раздражения. Рецепторы - это молекулы либо молекулярные комплексы, расположенные на поверхности клеток, а также способны распознавать химические специфические группировки, молекулы либо другие клетки, связывать их. У наиболее сложных вирионов такие рецепторы располагаются с внешней оболочки в виде шиповидного выроста или ворсинки, у простых вирионов они находятся, как правило, на поверхности капсида.

Механизм адсорбции на поверхности восприимчивой клетки основывается на взаимодействии рецепторов с так называемыми комплементарными рецепторами "хозяйской" клетки. Рецепторы вириона и клетки являются некими специфическими структурами, которые расположены на поверхности.
Аденовирусы и миксовирусы адсорбируются непосредственно на мукопротеиновых рецепторах, а арбовирусы и пикорнавирусы ― на липопротеиновых рецепторах.
У вириона миксовирусов нейраминидаза разрушает мукогфотеиновый рецептор и отщепляет N-ацетилнейраминовые кислоты от олигосахарида, который содержит в себе галактозу и галактозамин. Их взаимодействия на данном этапе обратимы, ведь на них значительно влияет температура, реакция среды и солевые компоненты. Адсорбции вириона препятствуют гепарин и сульфатированные полисахариды, несущие при этом отрицательный заряд, однако их ингибирующее воздействие снимается некоторыми поликарионами (экмолин, ДЭАЭ-декстран, протаминсулъфат), нейтрализующие отрицательный заряд от сульфатированных полисахаридов.
Попадание вириона в "хозяйскую" клетку
Путь внедрения вируса в чувствительную к нему клетку не всегда будет одним и тем же. Многие вирионы способны проникать в клетки методом пиноцитоза, что в переводе с греческого означает "пить", "выпивать". При данном методе пиноцитозная вакуоль будто бы втягивает вирион непосредственно внутрь клетки. Остальные вирионы могут проникать в клетку напрямую сквозь ее оболочку.

Контакт фермента нейраминидаза с клеточными мукопротеидами способствует попаданию вирионов в клетку среди миксовирусов. Результаты исследований последних лет доказывают, что ДНК и РНК вирионов от внешней оболочки не отделяются, т. е. вирионы проникают целиком в чувствительные клетки путем пиноцитоза или виропексиса. На настоящий момент это подтверждено в отношении вируса оспы, осповакцины, а также других вирусов, выбирающих средой обитания организм животных. Если говорить о фагах, они заражают нуклеиновой кислотой клетки. Механизм заражения основывается на том, что те вирионы, которые содержатся в вакуолях клеток, гидролизуются ферментами (липаз, протеаз), в процессе чего от оболочки фага освобождается ДНК и попадает в клетку.
Для проведения эксперимента выполнялось заражение клетки с помощью нуклеиновой кислоты, которая была выделена от некоторых вирусов, и вызывается один полный цикл репродукции вирионов. Однако в естественных условиях инфицирования при помощи такой кислоты не происходит.
Дезинтеграция
Следующий этап репродукции вирусов – дезинтеграция, которая представляет собой освобождение НК от капсида и внешней оболочки. После попадания вириона в клетки, капсид переживает некоторые изменения, приобретая чувствительность к клеточному протеазу, затем он разрушается, параллельно освобождая НК. У отдельных бактериофагов в клетки попадает свободная НК. Фитопатогенный вирус проникает через повреждение в клеточной стенке, а затем он адсорбируется на внутреннем клеточном рецепторе с одновременным высвобождением НК.
Репликация РНК и синтез вирусного белка
Следующим этапом репродукции вирусов является синтез вирусоспецифичного белка, который происходит с участием так называемых информационных РНК (у отдельных вирусов они находятся в составе вирионов, а у некоторых синтезируются только в зараженных клетках непосредственно на матрице вирионной ДНК или РНК). Происходит репликация вирусной НК.

Существует 2 вида РНК-полимераз. К таковым относятся: РНК-полимераза I, которая катализирует формирование репликативной формы непосредственно на матрице плюс-цепи, а также РНК-полимераза II, которая принимает участие в синтезе однонитчатой вирусной РНК на матрице репликативного типа. Синтез нуклеиновых кислот у мелких вирусов происходит в цитоплазме. Что касается вируса гриппа, то в ядре синтезируется внутренний белок и РНК. РНК выделяется затем из ядра и проникает в цитоплазму, в которой совместно с рибосомами начинает синтезировать вирусный белок.
После попадания вирионов в клетки, в них подавляется синтез нуклеиновой кислоты, а также клеточных белков. При репродукции вирусов, ДНК содержащих, на матрице в ядре синтезируется еще и-РНК, которая несет в себе информацию для синтеза белка. Механизм синтеза вирусного белка осуществляется на уровне клеточной рибосомы, а источником построения будет аминокислотный фонд. Активизация аминокислот осуществляется ферментами, при помощи и-РНК переносятся непосредственно в рибосомы (полисомы), в которых они располагаются уже в синтезированной молекуле белков.
Таким образом, в зараженных клетках синтез нуклеиновых кислот и белков вириона осуществляется в составе репликативно-транскриптивного сложного комплекса, который регулируется некой системой механизма.

Морфогенез вириона
Образование вирионов может произойти только в случае строго упорядоченного соединения структурных вирусных полипептидов, а также их НК. А это обеспечивается так называемой самосборкой молекул белка около НК.
Формирование вириона
Формирование вириона происходит с участием некоторых структурных компонентов, входящих в состав клетки. Вирусы герпеса, полиомиелита и осповакцины образуются в цитоплазме, а аденовирусы ― в ядре. Синтез вирусной РНК, а также формирование нуклеокапсида происходит непосредственно в ядре, а гемагглютинин формируется в цитоплазме. После этого нуклеокапсид перебирается из ядра в цитоплазму, в которой осуществляется образование оболочки вириона. Нуклеокапсид покрывается снаружи вирусными белками, а в состав вириона при этом включаются гемагглютинины и нейраминидазы. Именно таким образом происходит образование потомства, например, вируса гриппа.
Высвобождение вириона из "хозяйской" клетки
Из "хозяйской" клетки частицы вируса выделяются одновременно (во время разрушения клеток) либо постепенно (без каких-либо разрушений клеток).
Именно в таком виде и происходит репродукция вирусов. Вирионы высвобождаются из клеток, как правило, двумя способами.
Первый метод
Первый способ подразумевает следующее: после абсолютного созревания вирионов непосредственно внутри клетки они округляются, там образуются вакуоли, а затем разрушается и клеточная оболочка. По завершению этих процессов вирионы выходят все одновременно и полностью из клеток (пикорнавирусы). Данный способ принято называть литическим.

Второй метод
Второй способ подразумевает процесс освобождения вирионов по мере их созревания в течение 2―6 часов на цитоплазматической мембране (миксовирусы и арбовирусы). Выделению из клетки миксовирусов способствует нейраминидазы, разрушающие клеточную оболочку. Во время этого способа 75-90 % вирионов выходят спонтанно в культуральную среду, а клетки постепенно погибают.
Сайт СТУДОПЕДИЯ проводит ОПРОС! Прими участие :) - нам важно ваше мнение.
Репродукция вирусов. Взаимодействие с хозяеном. Культивирование.
Вирусы не размножаются бинарным делением. В 50-х годах ХХ в. было установлено, что размножение вирусов происходит путем репродукции (англ. reproduce – воспроизводить, делать копию), т.е. путем воспроизведения их нуклеиновых кислот и синтеза белков с последующей сборкой вирионов. Эти процессы происходят в разных частях клетки хозяина (например, в ядре и цитоплазме). Такой разобщенный способ репродукции получил название дизъюнктивного.Репродукция вирусов характеризуется последовательной сменой отдельных стадий:
1) Адсорбция. Проникновение вирусной частицы в клетку начинается с ее адсорбции на клеточной поверхности благодаря взаимодействию клеточных и вирусных рецепторов. Рецепторы (лат. receptor– принимающий) – чувствительные специальные образования, воспринимающие раздражения, это молекулы или молекулярные комплексы на поверхности клеток, способные распознавать специфические химические группировки, молекулы или другие клетки и связывать их. У сложных вирионов рецепторы располагаются на внешней оболочке в виде шиповидных выростов или ворсинок, у простых вирионов – на поверхности капсида.
2) Проникновение вириона в клетку хозяина. Пути внедрения вирусов в чувствительные к ним клетки неодинаковы. Многие вирионы могут проникать в клетку путем пиноцитоза (греч. pino – пить, выпивать), когда образующаяся пиноцитарная вакуоль втягивает вирион внутрь клетки. Другие вирионы могут попадать в клетку прямым путем через ее оболочку.
3) Дезинтеграция (или "раздевание") вириона – освобождение НК от внешней оболочки и капсида. После проникновения вириона в клетку капсид претерпевает изменения, приобретает чувствительность к клеточным протеазам, разрушается, освобождая НК. У некоторых бактериофагов в клетку проникает свободная НК. Фитопатогенные вирусы проникают через повреждения в клеточной стенке, после чего адсорбируются на внутренних клеточных рецепторах и высвобождается НК.
4) Синтез вирусных белков и репликация НК. Синтез вирусоспецифичных белков происходит с участием информационных РНК (у одних вирусов они входят в состав вирионов, а у других синтезируются в зараженных клетках на матрице вирионной РНК или ДНК). Происходит репликация вирусных НК.
5) Сборка, или морфогенез вириона. Формирование вирионов возможно только при условии строго упорядоченного соединения вирусных структурных полипептидов и их НК, что обеспечивается самосборкой белковых молекул вокруг НК
6) Выход вириона из клетки хозяина. Из клетки вирусные частицы выходят одновременно (при разрушении клеток) или постепенно (без разрушения клеток).
Вирусы не способны размножаться на питательных средах - это строгие внутриклеточные паразиты. Более того, в отличие от риккетсий и хламидий, вирусы в клетке хозяина не растут и не размножаются путем деления. Составные части вируса - нуклеиновые кислоты и белковые молекулы синтезируются в клетке хозяина раздельно, в разных частях клетки - в ядре и в цитоплазме. При этом клеточные белоксинтезирующие системы подчиняются вирусному геному, его НК.
Репродукция вируса в клетке происходит в несколько фаз:
· Первая фаза - адсорбция вируса на поверхности клетки, чувствительной к данному вирусу.
· Вторая фаза - проникновение вируса в клетку хозяина путем виропексиса.
Адсорбция вирионов на клетке. Механизм адсорбции вириона на восприимчивой клетке основан на взаимодействии его рецепторов с комплементарными рецепторами клетки. Рецепторы клетки и вириона являются специфическими структурами, расположенными на их поверхности. Миксовирусы и аденовирусы адсорбируются на мукопротеиновых рецепторах, а пикорнавирусы и арбовирусы ― на липопротеиновых рецепторах. Нейраминидаза у вириона миксовирусов разрушает мукогфотеиновые рецепторы и отщепляет N-ацетилнейраминовую кислоту от олигосахарида, содержащего галактозамин и галактозу. Их взаимодействия на этом этапе обратимы, так как на них влияют температура, солевые компоненты и реакция среды. Адсорбции вириона на клетке препятствуют сульфатированные полисахариды и гепарин, несущие отрицательный заряд, но их ингибирующее действие снимается поликар-тионами (ДЭАЭ-декстран, экмолин, протамннсулъфат), которые нейтрализуют отрицательный заряд сульфатированных полисахаридов.
Проникновение вириона в клетку. Процесс проникновения вирионов в клетку у миксовирусов осуществляется ферментом нейраминидазой, который вступает в непосредственный контакт с мукопротеидами клетки. Научные факты, накопленные за последние годы, показывают, что РНК и ДНК вирионов не отделяются от внешней их оболочки, т. е. вирионы целиком проникают в чувствительную клетку путем виропексиса или пиноцитоза. Это доказано в отношении вирусов оспы, осповакцины и других вирусов животных. Что касается фагов, то они заражают клетки своей нуклеиновой кислотой. Механизм заражения основан на том, что вирионы, содержащиеся в вакуолях клетки, гидролйзуются ферментами (протеаз, липаз). При этом освобождается ДНК от внешней оболочки фага и проникает в клетку.
В эксперименте заражают клетки нуклеиновой кислотой, выделенной от некоторых вирусов, и вызывают один цикл репродукции "вирионов. Но в естественных условиях передача инфекции с помощью инфекционной кислоты не происходит.
Синтез компонентов ДНК-вирусов. После проникновения вирионов в клетку в ней подавляется синтез нуклеиновых кислот и клеточных белков. В ядре на матрице ДНК-вируса синтезируется и-РНК, несущая информацию для синтеза белков. Механизм синтеза вирусных белков осуществляется на клеточных рибосомах, и источником их построения является аминокислотный фонд клетки. Активизация аминокислот происходит ферментами, с помощью и-РНК переносятся в рибосомы (полисомы), где они располагаются в синтезированной молекуле белка.
Таким образом, в зараженной клетке синтез нуклеиновой кислоты и белков вириона происходит в составе сложного репликатив-но-транскриптивного комплекса, который, по-видимому, регулируется определенной системой контрольного механизма.
Формирование вириона осуществляется с участием структурных компонентов клетки. Вирусы полиомиелита, герпеса и осповакцины формируются в цитоплазме, а аденовирусов ― в 'ядре. Синтез вирусной РНК и образование нуклеокапсида (S-анти-гена) происходит в ядре, а гемагглютцнина (V-антигена) ―в цитоплазме. Затем S-антиген переходит из ядра в цитоплазму, где осуществляется формирование оболочки вириона. S-антиген покрывается вирусными белками, и в состав вириона включаются-гемагглютинины и нейраминидаза. И так происходит формирование потомства вируса гриппа.
Выход вирусов из клетки. Вирионы освобождаются из клеток двумя способами. Первый способ ― после полного созревания вирионов внутри клетки последние округляются, в них образуются вакуоли, разрушается клеточная оболочка; вирионы выходят одновременно и полностью из клетки (рикорнавирусы). Этот способ называется литическим. Второй способ ― вирионы освобождаются по мере созревания их на цитоплазматической мембране в течение 2―6 часов (арбовирусы,и миксовирусы). Освобождению миксовирусов из клетки, по-видимому, способствует нейраминидаза, которая разрушает клеточную оболочку. При этом способе 75― 90% вирионов спонтанно выходят в культуральную среду и клетки погибают постепенно).
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Для вирусов характерен дизъюнктивный (от disjuncus -- разобщенный) способ репродукции-размножения. Потомство вируса возникает в результате сборки нуклеиновых кислот и белковых субъединиц, которые синтезируются раздельно клеткой хозяина. Проникновение вируса в клетку и воспроизведение себе подобных проходит в несколько фаз: проникновение в клетку хозяина, синтез ферментов, необходимых для репликации вирусных нуклеиновых кислот, синтез вирусных частей, сборка и композиция зрелых вирионов, выход зрелых вирионов из клетки.
Фаза I -- адсорбция вириона на поверхности клетки.
Протекает в две стадии: первая -- неспепифическая, когда вирус удерживается на поверхности клетки благодаря возникновению противоположных зарядов между отдельными участками мембраны клеток и вируса. Эта фаза взаимодействия вируса с клеткой обратима, на нее оказывают влияние такие факторы, как рН и солевой состав среды.
Вторая стадия -- специфическая, когда взаимодействуют специфические рецепторы вируса и рецепторы клетки, комплементарные друг другу. По химической природе рецепторы клетки могут быть мукопротеидашг (или мукополисахаридами) и липопротеидами. Разные вирусы фиксируются на разных рецепторах: вирусы гриппа, парагриппа, аденовирусы -- на мукопротеидах, а вирусы клещевого энцефалита, полиомиелита -- на липопротеидах.
Фаза II -- проникновение вируса в клетку. Электроноскопические наблюдения за процессом проникновения вирусов в чувствительные к ним клетки показали, что оно осуществляется посредством механизма, напоминающего пиноцитоз, или, как чаще называют, виропексис. В месте адсорбции вируса клеточная стенка втягивается внутрь клетки, образуется вакуоль, в которой оказывается вирион. Параллельно клеточные ферменты (липазы и протеазы) вызывают депротеинизацию вириона -- растворение белковой оболочки и освобождение нуклеиновой кислоты.
Фаза III -- скрытый период (период эклипса -- исчезновения). В этот период в клетке невозможно определить наличие инфекционного вируса ни химическими, ни электронно-микроскопическими, ни серологическими методами. О сущности этого явления и его механизмов пока известно мало. Предполагается, что в скрытой фазе нуклеиновая кислота вируса проникает в хромосомы клетки и вступает с ними в сложные генетические взаимоотношения.
Фаза IV -- синтез компонентов вириона. В этой фазе вирус и клетка представляют единое целое, вирусная нуклеиновая кислота выполняет генетическую функцию, индуцирует образование ранних белков и изменяет Функцию рибосом. Ранние белки подразделяются на:
а) белки-ингибиторы (репрессоры), подавляющие метаболизм клеток
б) белки-ферменты (полимеразы), обеспечивающие синтез вирусных нуклеиновых кислот.
Синтез нуклеиновых кислот и белков протекает неодновременно и в разных структурных частях клетки. У вирусов, содержащих ДНК или РНК, эти процессы имеют некоторые различия и особенности.
Процесс формирования вирионов начинается спустя определенное время после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность этого периода довольно вариабельна и предопределяется природой вируса -- для РНК-содержащих обычно короче, чем для ДНК-вирусов. Например, продукция полных вирусных частиц осповакцины начинается приблизительно спустя 5--6 ч после инфицирования клеток и продолжается в течение последующих 7--8 ч, т. е. после того как синтез вирусной ДНК уже завершен.
Между нуклеиновой кислотой и соответствующим белковыми субъединицами образуются очень прочные связи, о чем свидетельствуют трудности отделения белка от вирусной нуклеиновой кислоты. Большую прочность вирусной частице придают входящие в ее состав углеводы и особенно липиды.
Формирование вирионов, так же как и синтез компонентов вируса, происходит в разных местах клетки, при участии различных клеточных структур. После завершения процесса формирования образуется зрелая дочерняя вирусная частица, обладающая всеми свойств вами родительского вириона. Но иногда наблюдается образование так называемых неполных вирусов, которые состоят или только из нуклеиновой кислоты, или из белка, или из вирусных частиц, формирование которых остановилось в какой-то промежуточной стадии.
Фаза VI -- выход зрелых вирионов из клетки. Существуют два основных механизма выхода зрелых вирионов из клетки: 1) выход вириона с помощью почкования. В этом случае наружная оболочка вириона происходит из клеточной мембраны, она содержит как материал клетки хозяина, так и вирусный материал; 2) выход зрелых вирионов из клетки через бреши в мембране. Эти вирусы не имеют наружной оболочки. При таком механизме выхода вирусов клетка, как правило, погибает и в среде появляется большое количество вирусных частиц.
Особенности репродукции вирусов
1. Периоды осуществления продуктивной вирусной инфекции
2. Репликация вируса
1. Продуктивная вирусная инфекция осуществляется в 3 периода:
· начальный период включает стадии адсорбции вируса на клетке, проникновения в клетку, дезинтеграции (депротеинизации) или "раздевания" вируса. Вирусная нуклеиновая кислота была доставлена в соответствующие клеточные структуры и под действием лизосомальных ферментов клетки освобождается от защитных белковых оболочек. В итоге формируется уникальная биологическая структура: инфицированная клетка содержит 2 генома (собственный и вирусный) и 1 синтетический аппарат (клеточный);
· после этого начинается вторая группа процессов репродукции вируса, включающая средний и заключительный периоды, во время которых происходят репрессия клеточного и экспрессия вирусного генома. Репрессию клеточного генома обеспечивают низкомолекулярные регуляторные белки типа гистонов, синтезируемые в любой клетке. При вирусной инфекции этот процесс усиливается, теперь клетка представляет собой структуру, в которой генетический аппарат представлен вирусным геномом, а синтетический аппарат -- синтетическими системами клетки.
2. Дальнейшее течение событий в клетке направлено на репликацию вирусной нуклеиновой кислоты (синтез генетического материала для новых вирионов) и реализацию содержащейся в ней генетической информации (синтез белковых компонентов для новых вирионов). У ДНК-содержащих вирусов, как в прокариотических, так и в эукариотических клетках, репликация вирусной ДНК происходит при участии клеточной ДНК-зависимой ДНК-полимеразы. При этом у однонитевых ДНК-содержащих вирусов сначала образуется комплементарная нить -- так называемая репликативная форма, которая служит матрицей для дочерних молекул ДНК.
3. Реализация генетической информации вируса, содержащейся в ДНК, происходит следующим образом: при участии ДНК-зависимой РНК-полимеразы синтезируются и-РНК, которые поступают на рибосомы клетки, где и синтезируются вирусспецифические белки. У двунитевых ДНК-содержащих вирусов, геном которых транскрибируется в цитоплазме клетки хозяина, это собственный геномный белок. Вирусы, геномы которых транскрибируются в ядре клетки, используют содержащуюся там клеточную ДНК-зависимую РНК-полимеразу.
У РНК-содержащих вирусов процессы репликации их генома, транскрипции и трансляции генетической информации осуществляются иными путями. Репликация вирусных РНК, как минус-, так и плюс-нитей, осуществляется через репликативную форму РНК (комплементарную исходной), синтез которой обеспечивает РНК-зависимая РНК-полимераза -- это геномный белок, который есть у всех РНК-содержащих вирусов. Репликативная форма РНК минус-нитевых вирусов (плюс-нить) служит не только матрицей для синтеза дочерних молекул вирусной РНК (минус-нитей), но и выполняет функции и-РНК, т. е. идет на рибосомы и обеспечивает синтез вирусных белков (трансляцию).
У плюс-нитевых РНК-содержащих вирусов функцию трансляции выполняют ее копии, синтез которых осуществляется через репликативную форму (минус-нить) при участии вирусных РНК-зависимых РНК-полимераз.
У некоторых РНК-содержащих вирусов (реовирусы) имеется совершенно уникальный механизм транскрипции. Он обеспечивается специфическим вирусным ферментом -- ревертазой (обратной транскриптазой) и называется обратной транскрипцией. Суть ее состоит в том, что вначале на матрице вирусной РНК при участии обратной транскрипции образуется транскрипт, представляющий собой одну нить ДНК. На нем с помощью клеточной ДНК-зависимой ДНК-полимеразы синтезируется ,вторая нить и формируется двунитевой ДНК-транскрипт. С него обычным путем через образование и-РНК происходит реализация информации вирусного генома.
Результатом описанных процессов репликации, транскрипции и трансляции является образование дочерних молекул вирусной нуклеиновой кислоты и вирусных белков, закодированных в геноме вируса.
После этого наступает третий, заключительный период взаимодействия вируса и клетки. Из структурных компонентов (нуклеиновых кислот и белков) на мембранах цитоплазматического ретикулума клетки собираются новые вирионы. Клетка, геном которой был репрессирован (подавлен), обычно гибнет. Вновь сформировавшиеся вирионы пассивно (в результате гибели клетки) или активно (путем почкования) покидают клетку и оказываются в окружающей ее среде.
Таким образом, синтез вирусных нуклеиновых кислот и белков и сборка новых вирионов происходят в определенной последовательности (разобщены во времени) и в разных структурах клетки (разобщен в пространстве), в связи с чем способ репродукции вирусов и был назван дизъюнктивным (разобщенным). При абортивной вирусной инфекции процесс взаимодействия вируса с клеткой по тем или иным причинам прерывается до того, как произошло подавление клеточного генома. Очевидно, что в этом случае генетическая информация вируса реализована не будет и репродукции вируса не происходит, а клетка сохраняет свои функции неизменными. вирион клетка вирус
При латентной вирусной инфекции в клетке одновременно функционируют оба генома, а при вирусиндуцированных трансформациях вирусный геном становится частью клеточного, функционирует и наследуется вместе с ним.
2. Вирусология. Под редакцией Филдса Б., Найта Д., тт. 1-3, М., 1989.
3. Госманов Р.Г., Колычев Н.М. Ветеринарная вирусология. М.: КолосС. - 2003.
4. Белоусова Р.В., Преображенская Э.А., Третьякова И.В. Ветеринарная вирусология: Учебник для вузов (под ред. Белоусовой Р.В.). - М.: КолосС. - 2007
Для вирусов характерен дизъюнктивный (от disjuncus — разобщенный) способ репродукции-размножения. Потомство вируса возникает в результате сборки нуклеиновых кислот и белковых субъединиц, которые синтезируются раздельно клеткой хозяина.
Проникновение вируса в клетку и воспроизведение себе подобных проходит в несколько фаз:
1.проникновение в клетку хозяина,
2.синтез ферментов, необходимых для репликации вирусных нуклеиновых кислот,
3.синтез вирусных частей,
4.сборка и композиция зрелых вирионов,
5.выход зрелых вирионов из клетки.
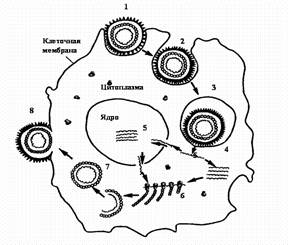
Стадии репродукции вирусов.
1 - адсорбция вириона на клетке; 2 - проникновение вириона в клетку путем виропексиса;
3 - вирус внутри вакуоли клетки; 4 - `раздевание вириона вируса; 5 - репликация вирусной нуклеиновой кислоты; 6 - синтез вирусных белков на рибосомах клетки; 7 - формирование вириона; 8 - выход вириона из клетки путем почкования.
Фаза I — адсорбция вириона на поверхности клетки.
Протекает в две стадии: первая — неспецифическая, когда вирус удерживается на поверхности клетки при помощи электростатических сил, т. е. благодаря возникновению противоположных зарядов между отдельными участками мембраны клеток и вируса. Эта фаза взаимодействия вируса с клеткой обратима, на нее оказывают влияние такие факторы, как рН и солевой состав среды.
Вторая стадия — специфическая, когда взаимодействуют специфические рецепторы вируса и рецепторы клетки, комплементарные друг другу. По химической природе рецепторы клетки могут быть мукопротеидами (или мукополисахаридами) и липопротеидами. Разные вирусы фиксируются на разных рецепторах: вирусы гриппа, парагриппа, аденовирусы — на мукопротеидах, а вирусы клещевого энцефалита, полиомиелита — на липопротеидах.
Фаза II — проникновение вируса в клетку. Электроноскопические наблюдения за процессом проникновения вирусов в чувствительные к ним клетки показали, что оно осуществляется посредством механизма, напоминающего пиноцитоз, или, как чаще называют, виропексис. В месте адсорбции вируса клеточная стенка втягивается внутрь клетки, образуется вакуоль, в которой оказывается вирион. Параллельно клеточные ферменты (липазы и протеазы) вызывают депротеинизацию вириона — растворение белковой оболочки и освобождение нуклеиновой кислоты.
Фаза III — скрытый период (период эклипса — исчезновения). В этот период в клетке невозможно определить наличие инфекционного вируса ни химическими, ни электронно-микроскопическими, ни серологическими методами. О сущности этого явления и его механизмов пока известно мало. Предполагается, что в скрытой фазе нуклеиновая кислота вируса проникает в хромосомы клетки и вступает с ними в сложные генетические взаимоотношения.
Фаза IV — синтез компонентов вириона. В этой фазе вирус и клетка представляют единое целое, вирусная нуклеиновая кислота выполняет генетическую функцию, индуцирует образование ранних белков и изменяет функцию рибосом. Ранние белки подразделяются на:
а) белки-ингибиторы (репрессоры), подавляющие метаболизм клеток
б) белки-ферменты (полимеразы), обеспечивающие синтез вирусных нуклеиновых кислот.
Синтез нуклеиновых кислот и белков протекает неодновременно и в разных структурных частях клетки. У вирусов, содержащих ДНК или РНК, эти процессы имеют некоторые различия и особенности.
Процесс формирования вирионов начинается спустя определенное время после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность этого периода довольно вариабельна и предопределяется природой вируса — для РНК-содержащих обычно короче, чем для ДНК-вирусов. Например, продукция полных вирусных частиц осповакцины начинается приблизительно спустя 5—6 ч после инфицирования клеток и продолжается в течение последующих 7—8 ч, т. е. после того как синтез вирусной ДНК уже завершен.
Между нуклеиновой кислотой и соответствующим белковыми субъединицами образуются очень прочные связи, о чем свидетельствуют трудности отделения белка от вирусной нуклеиновой кислоты. Большую прочность вирусной частице придают входящие в ее состав углеводы и особенно липиды.
Формирование вирионов, так же как и синтез компонентов вируса, происходит в разных местах клетки, при участии различных клеточных структур. После завершения процесса формирования образуется зрелая дочерняя вирусная частица, обладающая всеми свойствами родительского вириона. Но иногда наблюдается образование так называемых неполных вирусов, которые состоят или только из нуклеиновой кислоты, или из белка, или из вирусных частиц, формирование которых остановилось в какой-то промежуточной стадии.
Фаза VI — выход зрелых вирионов из клетки. Существуют два основных механизма выхода зрелых вирионов из клетки:
1) выход вириона с помощью почкования. В этом случае наружная оболочка вириона происходит из клеточной мембраны, она содержит как материал клетки хозяина, так и вирусный материал;
2) выход зрелых вирионов из клетки через бреши в мембране. Эти вирусы не имеют наружной оболочки. При таком механизме выхода вирусов клетка, как правило, погибает и в среде появляется большое количество вирусных частиц.
Причиной гибели зараженной клетки могут быть три механизма:
2.защитная реакция клетки, запускающая генетическую программу ее гибели (апоптоз);
3. иммунная система организма, уничтожающая зараженную клетку.
Кроме продуктивного типа взаимодействия вируса и клетки возможно интегративное сосуществование или вирогения. Вирогения характеризуется интеграцией (встраиванием) нуклеиновой кислоты вируса в геном клетки, а также репликацией и функционированием вирусного генома как составной части генома клетки. Для интеграции с клеточным геномом необходимо возникновение кольцевой формы двунитевой ДНК вируса. Встроенная в состав хромосомы клетки вирусная ДНК называется провирусом. Провирус реплицируется в составе хромосомы и переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. Под влиянием некоторых физических или химических факторов провирус может переходить в автономное состояние с развитием продуктивного типа взаимодействия с клеткой. Дополнительная генетическая информация провируса при вирогении сообщает клетке новые свойства, что может быть причиной развития опухолей, аутоиммунных и хронических заболеваний. На способности вирусов к интеграции с геномом клетки основаны персистенция (от лат. persisto - постоянно пребывать, оставаться) вирусов в организме и развитие персистентных вирусных инфекций. Например, вирус гепатита В способен вызывать персистирующие поражения с развитием хронического гепатита и часто опухолей печени.
Читайте также: