
Способность к обратной транскрипции характерна для вирусов
1. Сходное строение клеток растений и животных - доказательство
А) их родства
Б) общности происхождения организмов всех царств
В) происхождения растений от животных
Г) усложнения организмов в процессе эволюции
Д) единства органического мира
Е) многообразия организмов
2. Какие функции выполняет комплекс Гольджи?
А) синтезирует органические вещества из неорганических
Б) расщепляет биополимеры до мономеров
В) накапливает белки, липиды, углеводы, синтезируемые в клетке
Г) обеспечивает упаковку и вынос веществ из клетки
Д) окисляет органические вещества до неорганических
Е) участвует в образовании лизосом
3. Установите соответствие между признаком организма и группой, для которой он характерен: 1-прокариоты, 2-вирусы.
А) клеточное строение тела
Б) наличие собственного обмена веществ
В) встраивание собственной ДНК в ДНК клетки хозяина
Г) состоит из нуклеиновой кислоты и белковой оболочки
Д) размножение делением надвое
Е) способность к обратной транскрипции
А1 Б1 В2 Г2 Д1 Е2
4. К автотрофам относят
А) споровые растения
Б) плесневые грибы
В) одноклеточные водоросли
Г) хемотрофные бактерии
Д) вирусы
Е) большинство простейших
5. Установите последовательность процессов, происходящих в ходе мейоза
А) расположение пар гомологичных хромосом в экваториальной плоскости
Б) конъюгация, кроссинговер гомологичных хромосом
В) расположение в плоскости экватора и расхождение сестринских хромосом
Г) образование четырёх гаплоидных ядер
Д) расхождение гомологичных хромосом
5а. Какие признаки характеризуют мейоз?
А) наличие двух следующих одно за другим делений
Б) образование двух клеток с одинаковой наследственной информацией
В) расхождение гомологичных хромосом в разные клетки
Г) образование диплоидных дочерних клеток
Д) отсутствие интерфазы перед первым делением
Е) конъюгация и кроссинговер хромосом
6. Установите соответствие между характеристикой гаметогенеза и его видом: 1-овогенез, 2-сперматогенез
А) образуется одна крупная половая клетка
Б) образуются направительные клетки
В) формируется много мелких гамет
Г) питательные вещества запасаются в одной из четырех клеток
Д) образуются подвижные гаметы
7. Установите последовательность процессов, происходящих в клетке с хромосомами в интерфазе и последующем митозе
А) расположение хромосом в экваториальной плоскости
Б) репликация ДНК и образование двухроматидных хромосом
В) спирализация хромосом
Г) расхождение сестринских хромосом к полюсам клетки
7+. Какие структуры клетки претерпевают наибольшие изменения в процессе митоза?
А) ядро
Б) цитоплазма
В) рибосомы
Г) лизосомы
Д) клеточный центр
Е) хромосомы
8. Установите соответствие между строением органоида и его видом: 1-клеточный центр, 2-рибосома
А) состоит из двух перпендикулярно расположенных цилиндров
Б) состоит из двух субъединиц
В) образован микротрубочками
Г) содержит белки, обеспечивающие движение хромосом
Д) содержит белки и нуклеиновую кислоту
9. Установите последовательность процессов, происходящих при фагоцитозе
А) поступление мономеров в цитоплазму
Б) захват клеточной мембраной питательных веществ
В) гидролиз полимеров до мономеров
Г) образование фагоцитозного пузырька внутри клетки
Д) слияние фагоцитозного пузырька с лизосомой
10. Вирусы, в отличие от бактерий
А) имеют клеточную стенку
Б) адаптируются к среде
В) состоят только из нуклеиновой кислоты и белка
Г) размножаются вегетативно
Д) не имеют собственного обмена веществ
Е) ведут только паразитический образ жизни
12. Основные положения клеточной теории позволяют сделать выводы о
А) влиянии среды на приспособленность
Б) родстве организмов
В) происхождении растений и животных от общего предка
Г) развитии организмов от простого к сложному
Д) сходном строении клеток всех организмов
Е) возможности самозарождения жизни из неживой материи
12+. Основные положения клеточной теории позволяют сделать выводы о
А) биогенной миграции атомов
Б) родстве организмов
В) происхождении растений и животных от общего предка
Г) появлении жизни на Земле около 4,5 млрд. лет назад
Д) сходном строении клеток всех организмов
Е) взаимосвязи живой и неживой природы
12++. Какие положения содержит клеточная теория?
А) Новые клетки образуются в результате деления материнской клетки
Б) В половых клетках содержится гаплоидный набор хромосом
В) Клетки сходны по химическому составу
Г) Клетка – единица развития всех организмов
Д) Клетки тканей всех растений и животных одинаковы по строению
Е) Все клетки содержат молекулы ДНК
13. Сходство клеток грибов и животных состоит в том, что они имеют
А) оболочку из хитиноподобного вещества
Б) гликоген в качестве запасного углевода
В) оформленное ядро
Г) вакуоли с клеточным соком
Д) митохондрии
Е) пластиды
14. Какие функции выполняет в клетке плазматическая мембрана?
А) придает клетке жесткую форму
Б) отграничивает цитоплазму от окружающей среды
В) служит матрицей для синтеза иРНК
Г) обеспечивает поступление в клетку ионов и мелких молекул
Д) обеспечивает передвижение веществ в клетке
Е) участвует в поглощении веществ клеткой
15. Установите последовательность расположения структур в эукариотной клетке растения (начиная снаружи)
А) плазматическая мембрана
Б) клеточная стенка
В) ядро
Г) цитоплазма
Д) хромосомы
16. Установите соответствие между характеристикой клеток животных и их типом: 1-соматические, 2-половые (гаметы)
А) образуют ткани и органы
Б) участвуют в процессе оплодотворения
В) всегда гаплоидны
Г) имеют диплоидный набор хромосом
Д) образуются в процессе мейоза
Е) делятся путем митоза
А1 Б2 В2 Г1 Д2 Е1
17. Чем митохондрии отличаются от лизосом?
А) имеют наружную и внутреннюю мембраны
Б) имеют многочисленные выросты – кристы
В) участвуют в процессах освобождения энергии
Г) в них пировиноградная кислота окисляется до углекислого газа и воды
Д) в них биополимеры расщепляются до мономеров
Е) участвуют в обмене веществ
Современная биология поражает уникальностью и масштабностью своих открытий. На сегодняшний день эта наука изучает большинство процессов, которые скрыты от нашего глаза. Этим примечательна молекулярная биология – одно из перспективных направлений, которое помогает разгадать сложнейшие тайны живой материи.
Что такое обратная транскрипция
Обратная транскрипция (сокращенно ОТ) – это специфический процесс, характерный для большинства РНК-содержащих вирусов. Главной его особенностью является синтез двухцепочечной молекулы ДНК на базе матричной РНК.
ОТ не характерна для бактерий или эукариотических организмов. Главный фермент – ревертаза – играет ключевую роль в синтезе двухцепочечной ДНК.
История открытия
Идея о том, что молекула рибонуклеиновой кислоты может стать матрицей для синтеза ДНК, считалась абсурдной вплоть до 1970 года. Тогда Балтимор и Темин, работавшие отдельно друг от друга, почти одновременно открыли новый фермент. Они назвали его РНК-зависимая-ДНК-полимераза, или обратная транскриптаза.
Открытие данного энзима безоговорочно подтвердило существование организмов, способных к обратной транскрипции. В 1975 году оба ученых получили Нобелевскую премию. Через некоторое время Энгельгардт предложил альтернативное название обратной транскриптазы – ревертаза.

Почему ОТ противоречит центральной догме молекулярной биологии
Центральная догма – это принципиальная схема последовательного синтеза белка в любой живой клетке. Такая схема строится из трех компонентов: ДНК, РНК и белок.
Согласно центральной догме, РНК может синтезироваться исключительно на матрице ДНК, а уж затем РНК участвует в построении первичной структуры белка.
Эта догма была официально принята в научном обществе раньше, чем произошло открытие обратной транскрипции. Неудивительно, что идея об обратном синтезе ДНК из РНК долго отвергалась учеными. Лишь в 1970 году вместе с открытием ревертазы была поставлена точка в этом вопросе, что отразилось и в принципиальной схеме синтеза белка.
Ревертаза ретровирусов птиц
Процесс обратной транскрипции не обходится без участия РНК-зависимой-ДНК-полимеразы. Максимально изучена на сегодняшний день ревертаза ретровируса птиц.
Всего около 40 молекул этого белка можно обнаружить в одном вирионе этого семейства вирусов. Протеин состоит из двух субъединиц, которые находятся в равном количестве и выполняют три важнейшие функции ревертазы:
1) Синтез молекулы ДНК как на матрице одноцепочечной/двухцепочечной РНК, так и на основе дезоксирибонуклеиновых кислот.
2) Активация РНКазы Н, главная роль которой заключается в расщеплении молекулы РНК в комплексе РНК-ДНК.
3) Разрушение участков молекул ДНК для встраивания в геном эукариот.
Механизм ОТ
Этапы обратной транскрипции могут варьироваться в зависимости от семейства вирусов, т.е. от вида их нуклеиновых кислот.
Рассмотрим сначала те вирусы, которые используют ревертазу. Здесь процесс ОТ делится на 3 этапа:
Такой способ размножения вирионов характерен для некоторых онкогенных вирусов и вируса иммунодефицита человека (ВИЧ).
Стоит отметить, что для синтеза любой нуклеиновой кислоты на матрице РНК нужна затравка, или праймер. Праймер представляет собой короткую последовательность нуклеотидов, комплементарную 3’-концу молекулы РНК (матрицы) и играющую важную роль в инициации синтеза.
Когда готовые двухцепочечные молекулы ДНК вирусного происхождения встраиваются в геном эукариот, запускается обычный механизм синтеза белков вириона. В итоге “захваченная” вирусом клетка становится фабрикой по производству вирионов, где в больших количествах образуются необходимые молекулы протеинов и РНК.
Повторение таких циклов приводит как к репликации генома вируса, так и к образованию мРНК, способных к синтезу белка в условиях зараженной эукариотической клетки.
Биологическое значение обратной транскрипции
Процесс ОТ имеет первостепенное значение в жизненном цикле многих вирусов (в первую очередь ретровирусов, таких как ВИЧ). РНК вириона, атаковавшего клетку эукариот, становится матрицей для синтеза первой цепочки ДНК, на которой не трудно достроить и вторую цепь.
Полученная двухцепочечная ДНК вируса встраивается в геном эукариот, что приводит к активации процессов синтеза белков вириона и появлению большого количество его копий внутри зараженной клетки. В этом и заключается основная миссия ревертазы и ОТ в целом для вируса.
У эукариот также может встречаться обратная транскрипция в контексте ретротранспозонов – мобильных генетических элементов, способных самостоятельно транспортироваться из одного участка генома в другой. Такие элементы, как считают ученые, стали причиной эволюции живых организмов.
Ретротранспозон представляет собой участок ДНК эукариот, который кодирует несколько белков. Один из них, ревертаза, принимает непосредственное участие в делокализации такого ретротраспорозона.
Использование ОТ в науке
С того момента, как удалось выделить ревертазу в чистом виде, процесс обратной транскрипции взяли на вооружение ученые-биологи. Изучение механизма ОТ и сейчас помогает прочитать последовательности важнейших белков человека.
Дело в том, что геном эукариот, в том числе и нас, содержит неинформативные участки под названием интроны. Когда с такой ДНК считывается последовательность нуклеотидов и формируется одноцепочечная РНК, последняя лишается интронов и кодирует исключительно белок. Если с помощью ревертазы на матрице РНК синтезировать ДНК, ее легко потом секвенировать и узнать порядок нуклеотидов.
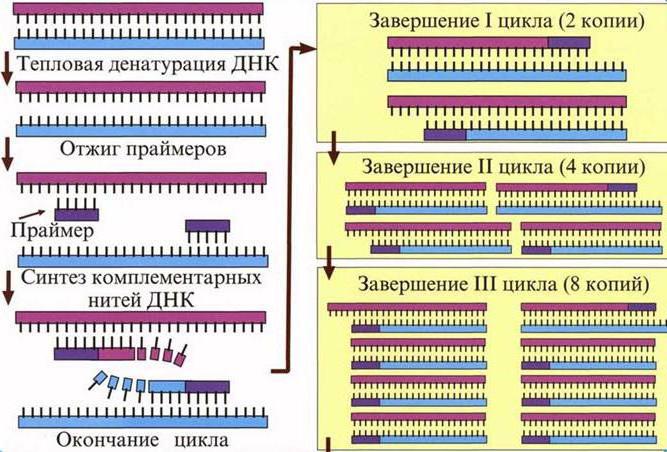
Нуклеиновая кислота, которая была образована с помощью обратной транскриптазы, называется кДНК. Она часто используется в полимеразной цепной реакции (ПЦР), чтобы искусственно увеличить число копий полученной копии кДНК. Данный метод используется не только в науке, но и в медицине: лаборанты определяют сходство такой ДНК с геномами различных бактерий или вирусов из общей библиотеки. Синтез векторов и внедрение их в бактерии – одно из перспективных направлений биологии. Если с помощью ОТ формировать ДНК человека и других организмов без интронов, такие молекулы легко внедряются в геном бактерий. Так последние становятся фабриками по производству необходимых человеку веществ (например, ферментов).
При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза.
Каждый вирион (полноценная вирусная частица, состоящая из нуклеиновой кислоты и белковой оболочки) ретровирусов содержит две идентичные цепи РНК размером от 8000 до 10 000 нуклеотидов. Области 5'- и 3'-концов обеих цепей модифицированы, как у всех эукариотических мРНК (5'-кэпы, З'-полиадениловые хвосты). Вирусные РНК имеют 5 структурных элементов: 1) прямые повторы на 5'- и З'-концах РНК (R); 2) последовательность из 80 -- 120 нуклеотидов, находящуюся около 5 концевого повтора (U5); 3) последовательность из 170 -- 1200 нуклеотидов около З'-концевого повтора (U3); 4) последовательность из 15 20 нуклеотидов (Р), в пределах которой клеточная тРНК комплементарно взаимодействует с ретровирусной РНК, что создает праймер для синтеза первой цепи ДНК; 5) сегмент Pu, находящийся непосредственно перед повтором U3 и являющийся сайтом для праймирования второй цепи ДНК -- такой сегмент одинаков у РНК всех ретровирусов определенного типа.
Этапы обратной транскрипции:
5. К моменту завершения синтеза первой цепи ДНК большая часть вирусной РНК разрушается РНКазой Н.
6. Синтез З'-конца второй цепи ДНК.
7. Удаление тРНК и оставшегося участка вирусной (+) РНК РНКазой Н.
8. Второй прыжок, в результате которого новосинтезированная вторая цепь ДНК комплементарно взаимодействует с тРНК-связывающей последовательностью первой цепи.
9. Удлинение З'-концов каждой цепи, образование дуплекса ДНК.
Вся последовательность реакций протекает без явного участия ферментов репликации клетки-хозяина (топоизомеразы, хеликазы, праймазы, ДНК-связывающего белка, лигазы и т.д.). При этом следует отметить, что молекулы вирусных ДНК длиннее молекул вирусных РНК, которые послужили матрицей для обратной транскрипции. Действительно, к 5'-концу (+) цепи вирусной ДНК добавилась последовательность U3, а к 3 -концу этой цепи -- последовательность U5. В результате на концах молекулы вирус специфической ДНК появился длинный (несколько сотен нуклеотидов) концевой повтор (ДКП или LTR.), имеющий структуру U3U5.

Рис.2. Схема обратной транскрипции ретровирусной РНК с образованием двуцепочечной ДНК (Darnell J., et.al. Molecular Cell Biology. - N. Y.: Scientific Amer. Books, 1986. - P. 1052)
Синтез ДНК на РНК-матрице in vitro и ревертаза используется в генетической инженерии для синтеза генов и их фрагментов, а также целенаправленного синтеза на матричных РНК комплементарных молекул ДНК (кДНК) для расшифровки первичной структуры РНК и белков.

Рис. 3. Схема получения кДНК с использованием ревертазы вируса и трех дополнительных ферментов: поли (А)-полимеразы, фрагмента Кленова ДНК-полимеразы I и нуклеазы S1. (Цитировано по)
Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А) - содержащих мРНК используют олигo (dT)-праймер, а для молекул РНК, не имеющих З'-поли (А) концов, -- химически синтезированные олигонуклеотиды, комплементарные З'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. сoli (остающаяся часть молекулы, которая сохраняет присущие ей каталитические активности).
Уже одно это открытие формально поставило РНК в центр основного постулата молекулярной генетики, так как показало, что поток генетической информации распространяется от РНК не в одном, а в двух направлениях: не только к белку, но и к ДНК. Все более глубокое проникновение в механизмы основных молекулярно-генетических процессов (репликацию, транскрипцию и трансляцию) способствовало возникновению понятия о неканонических функциях РНК, осознанию полифункциональности рибонуклеиновых кислот.
Введение ………………………………………………………………………. 3
1. Значение обратной транскрипции ………………………………………… 4
2. Механизм обратной транскрипции………………………………………. 10
3. Применение обратной транскрипции в молекулярной биологии вирусов………………………………………………………………………….
13
Реферат обратная транскрипция.doc
Обратная транскрипция: значение, механизм и применение в молекулярной биологии вирусов
| Введение ………………………………………………………………………. | 3 |
| 1. Значение обратной транскрипции …………………… …………………… | 4 |
| 2. Механизм обратной транскрипции……………………… ………………. | 10 |
| 3. Применение обратной транскрипции в молекулярной биологии вирусов…………………………………… ……………………………………. | 13 |
| Заключение ……………………………………………………………………. | 18 |
| Список литературы …………………………………………………………… | 21 |
Рис. 1 Превращение РНК в ДНК с помощью транскриптазы и встраивание в геном хозяина
При делении клетки встроенная копия ДНК вирусного генома удваивается и передается дочерним клеткам. Таким образом, наследственный материал вируса оказывается включенным в геном клетки. Вирусная РНК может образоваться позже путем копирования встроенной ДНК. Это приведет к образованию новых инфекционных вирусов.
В 1970 году Howard Temin и David Baltimore независимо друг от друга открыли фермент, названный обратной транскриптазой (ревертазой), и возможность обратной транскрипции была окончательно подтверждена.
В общем виде идею обратной транскрипции Тёмин высказал еще в 1964 г., работая к тому времени несколько лет под руководством своего учителя Р. Дульбекко с опухолеродными вирусами животных, но она была решительно отвергнута, так как считалось, что в природе не существует фермента, способного осуществить такой процесс. В 1970 г. Тёмин (одновременно независимо от него Д. Балтимор) экспериментально обнаружил такой фермент — ревертазу, фермент класса трансфераз, осуществляющий синтез молекулы ДНК на РНК как матрице. Одновременно Тёмин (и независимо от него Р. Дульбекко) выдвинул вскоре подтвержденную экспериментами теорию провируса — генома вируса, который полностью объединен с генетическим материалом клетки-хозяина в единую молекулу ДНК, что делает возможным перерождение нормальной клетки в раковую. Тем самым концепция обратной транскрипции была окончательно доказана, и в 1975 г. Тёмин (вместе с Д. Балтимором и Р. Дульбекко) был удостоен Нобелевской премии по физиологии и медицине.
По общему признанию обратная транскрипция, это открытие которое относится к числу выдающихся событий в молекулярной биологии. Оно имело крупное общебиологическое значение, и его влияние испытали многие разделы биологии и медицины, такие как молекулярная биология, молекулярная генетика, вирусология, онкология, энзимология и др.
1. Значение обратной транскрипции
Обратной транскрипцией, как известно, называют синтез полинуклео-тидных цепей молекул ДНК по матрице РНК, в отличие от обычной (прямой) транскрипции, заключающейся в синтезе молекул РНК на матрице ДНК (рис. 2).
Рисунок 2. Синтез молекул РНК на матрице ДНК
Обратная транскрипция катализируется особым ферментом, который находится в составе РНК-содержащих опухолеродных вирусов, именуемых, по современной терминологии, ретровирусами. Фермент, осуществляющий обратную транскрипцию, получил название РНК-зависимой ДНК-полимеразы, или обратной транскриптазы, а по предложению академика В. А. Энгельгардта ему было дано короткое наименование — ревертаза.
Таким образом, обратная транскрипция положила начало двум совершенно новым направлениям в биологии: с одной стороны, изучению детальных механизмов размножения опухолеродных вирусов и их трансформирующего действия на клетки, а с другой — получению генетического материала любой заданной специфичности, а это незаменимый инструмент исследования для широкого круга вопросов не только молекулярной биологии, но и молекулярной генетики, биологии развития, эмбриологии и др. От синтеза генетического материала в однотяжевой форме существует путь к получению двутяжевой формы ДНК, а следовательно, сочетание обратной транскрипции и репликации позволяет получать фрагменты ДНК, в том числе структурные гены и в принципе даже полные гены, включающие регуляторные участки, которые можно размножить методами генетической инженерии.
Все эти перспективы, открывшиеся перед молекулярной биологией, были достаточно быстро осознаны представителями молекулярной биологии, и в ряде стран началась широкая, интенсивная работа.
В 1971 г. академик В. А. Энгельгардт выдвинул идею организации особого международного научного проекта для целенаправленного развития исследований по обратной транскрипции на основе единой согласованной научной программы. К сотрудничеству были привлечены академии наук Германии и Чехословакии.
Развитие работ по обратной транскрипции требовало согласованных усилий представителей разных научных дисциплин — молекулярных биологов, энзимологов, вирусологов, химиков-органиков, генетиков, онкологов.
Чтобы дать представление о характере этих работ, следует хотя бы коротко упомянуть некоторые полученные результаты.
В 1976 г., вскоре после работ американских и швейцарских исследователей, участники проекта по изучению обратной транскрипции синтезировали с использованием ревертазы структурные гены глобинов. Затем синтезированные гены были использованы для получения рекомбинантных молекул ДНК, которые в свою очередь после введения в бактерии были размножены. Этот общий подход к синтезу генов и их размножению методами генетической инженерии был разработан на генах белков-глобинов, а затем использован для генов иммуноглобулинов мышей, инсулина рыб и др. Подобные исследования весьма важны. Некоторые полученные рекомбинантные ДНК — весьма ценные инструменты исследования, другие — имеют большое практическое значение.
В результате детального анализа механизма обратной транскрипции, удалось установить, что в начале синтеза главную роль играет не двуспиральный комплекс матрица—затравка (как предполагали некоторые исследователи), а З'ОН-группа, расположенная на определенном расстоянии от полинуклеотидной цепи матрицы. При удлинении цепи в ходе синтеза было обнаружено новое, весьма своеобразное явление: ревертаза может вести синтез непрерывной цепи ДНК даже в том случае, когда в полинуклеотидной цепи матрицы имеется перерыв ковалентной структуры. Возможно, что способность ревертазы переходить с одной матрицы на другую без перерыва в цепи синтезируемой ДНК — это специальное приспособление для синтеза провирусной ДНК при попадании ретровирусов в клетки.
Как уже упоминалось, фермент ревертаза входит в состав ретровирусов. В нормальных клетках также обнаруживаются ферменты, имеющие ревертазоподобную активность, однако они представляют собой либо обычные клеточные ДНК-полимеразы, либо происходят от ретровирусов. Уже давно в работах одного из первооткрывателей ревертазы X. Темина было высказано предположение о существовании обратной транскрипции в нормальных клетках, однако до сих пор этот вопрос остался невыясненным.
При тщательном изучении препарата ДНК-полимеразы из кишечной палочки обнаружили фракцию, обладающую РНК-зависимой ДНК-полиме-разной активностью, полностью отделимую от обычной ДНК-зависимой активности. Ряд свойств новооткрытого фермента резко отличает его от обычной ДНК-полимеразы, поэтому можно думать, что это отдельный, самостоятельный белок, а не часть какого-либо другого фермента. Весьма интересно, что количество бактериальной ревертазы при переносе с бедной среды на богатую в клетках кишечной палочки значительно возрастает, то есть индукция, сопровождающаяся активацией синтеза РНК и белков, активирует и ревертазу. Еще не известно, какие функции в нормальной бактериальной клетке выполняет этот фермент. В последнее время удалось обнаружить, что при индукции ферментов в клетках животных, по-видимому, наблюдаются сходные явления, то есть не исключено, что и в клетках эукариот при определенных состояниях может существовать обратная транскрипция, не связанная с размножением ретровирусов.
§ 15. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ В КЛЕТКЕ:
РЕПЛИКАЦИЯ, ТРАНСКРИПЦИЯ
В процессе размножения клеток генетическая информация передается от одного поколения клеток другому. При этом все клетки получают одинаковую информацию. Это, возможно, вследствие того, что перед делением клетки осуществляется репликация (удвоение) ДНК, в результате образуются две идентичные молекулы ДНК, которые и передаются потомкам. В структуре ДНК заложена способность этой молекулы к копированию.
Закодированная в ДНК генетическая информация реализуется в результате экспрессии генов. Экспрессия генов включает транскрипцию (копирование информации с ДНК на синтезируемую РНК) и последующую трансляцию (синтез на матрице РНК соответствующего белка).
Возможен поток информации и в направлении от РНК к ДНК, этот процесс носит название обратная транскрипция. В то же время информация не передается от белков нуклеиновым кислотам. Однако следует отметить, что белки играют важную роль в осуществлении процессов передачи информации, как между нуклеиновыми кислотами, так и от нуклеиновых кислот к белкам.
Описанные информационные взаимоотношения между ДНК, РНК и белками могут быть представлены в виде схемы:
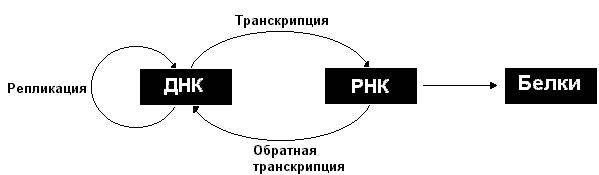
Рассмотрим более подробно процессы репликации, транскрипции и трансляции.
Интересно знать! В 1982 году Р.Д.Пальмитер с коллегами опубликовали фотографию, на которой рядом сидели две мыши. Одна из них была трансгенной, в ее ДНК встроили ген гормона роста крысы, другая была обычной мышкой. Трансгенная мышь в сравнении с обычной имела в два раза большие размеры. Причиной тому была экспрессия гена гормона роста, в результате которой у мыши синтезировался соответствующий гормон, который и определил ее гигантские размеры.
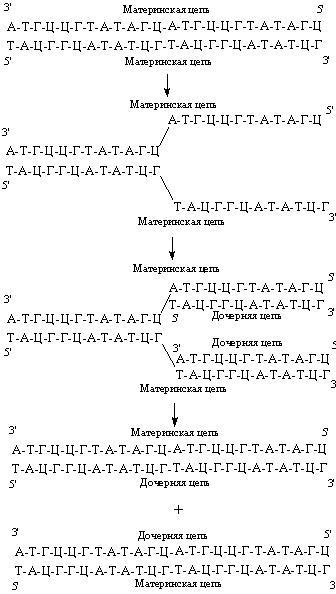
Рис. 44. В результате репликации образуются две дочерние молекулы ДНК.
Репликация ДНК
В основе репликации, или удвоения, ДНК лежит принцип комплементарности. Репликация начинается с разделения цепей, каждая из которых становится матрицей, определяющей нуклеотидную последовательность новой комплементарной цепи (рис. 44). В результате репликации образуются две дочерние молекулы ДНК, нуклеотидные последовательности которых идентичны между собой и с материнской молекулой ДНК. При этом две дочерние молекулы ДНК содержат одну вновь синтезированную (дочернюю) и одну материнскую цепи ДНК.
Субстратом для синтеза ДНК являются дезоксинуклеотид-5’-трифосфаты: дАТФ, дГТФ, дЦТФ и дТТФ. Уравнение этой реакции в простейшей виде выглядит так:
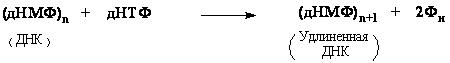
Данную реакцию катализирует фермент ДНК-полимераза. Она последовательно добавляет нуклеотиды к 3’-концу полинуклеотидной цепи. Следовательно, синтез новой цепи происходит в направлении от 5’-конца к 3’-концу. ДНК-полимераза может только наращивать цепь ДНК, начать же синтез ДНК с нуля она не может, т.е. для ее работы требуется цепь-затравка. ДНК-полимераза способна удлинять цепь только в присутствии цепи, играющей роль матрицы (рис. 45.). Нуклеотиды присоединяются к цепи-затравке в соответствии с принципом комплементарности, напротив аденина всегда будет встроен тимин, а напротив гуанина – цитозин.
Итогом репликации является образование двух дочерних молекул ДНК, являющихся точными копиями материнской. Благодаря этому возможна передача идентичной генетической информации от клетки к клетке.
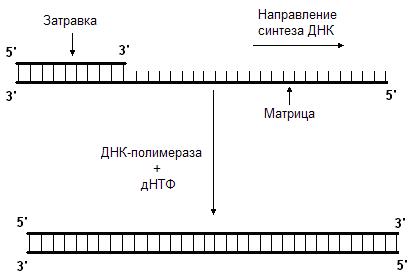
Рис. 45. ДНК-полимераза наращивает цепь в направлении 5’ 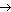
3’.
Для этого ей нужны матрица и затравка
Транскрипция
В результате транскрипции происходит синтез цепи РНК, нуклеотидная последовательность которой комплементарна одной из цепей ДНК; при этом генетическая информация, содержащаяся в ДНК, передается молекуле РНК. В результате транскрипции образуются три класса РНК: иРНК, тРНК, рРНК. Поскольку в основе транскрипции лежит принцип комплементарности, этот процесс имеет определенное сходство с репликацией. Однако между ними существует и важное различие. Если в процессе репликации копируется вся молекула ДНК, то при транскрипции транскрибируется только ее незначительная часть. Катализирует синтез РНК фермент РНК-полимераза. Транскрибируемый участок ДНК ограничен со стороны 3’-конца промотором, – участком с которым связывается РНК-полимераза, со стороны 5’-конца – терминатором – участком, в котором прерывается синтез РНК. Последовательность ДНК, ограниченная промотором и терминатором, представляет собой единицу транскрипции – транскриптон (рис. 46).
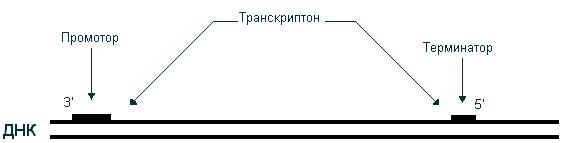
Рис. 46. Транскриптон
РНК-полимераза в качестве субстратов для синтеза РНК использует рибонуклеозид-5’-фосфаты (АТФ, ГТФ, ЦТФ и УТФ). Она удлиняет цепь, присоединяя нуклеотиды к 3’-концу, т.е. так же, как и ДНК-полимераза, наращивает цепь в направлении5’®3’. Этот фермент требует для своей работы в качестве матрицы одну из цепей ДНК. Из двух цепей ДНК транскрибируется только одна. В отличие от ДНК-полимеразы, РНК-полимераза не нуждается в затравке. Удлинение цепи РНК описывается уравнением:

Нуклеотиды присоединяются к цепи в соответствии с принципом комплементарности. Напротив аденина матричной цепи ДНК в растущую цепь РНК всегда будет встроен урацил, напротив гуанина – цитозин, напротив тимина – аденин и напротив цитозина – гуанин соответственно. Т.о., образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК (рис. 47).
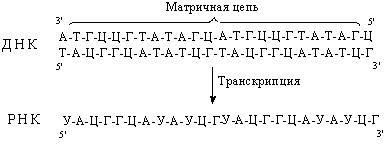
Рис. 47. Образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК
В процессе транскрипции выделяют три стадии: инициацию, элонгацию и терминацию. На стадии инициации РНК-полимераза, взаимодействуя с промотором, вызывает расхождение цепей ДНК и начинает синтез молекулы РНК. В ходе элонгации новосинтезированная цепь РНК образует (за счет спаривания ее оснований с основаниями матричной цепи ДНК) короткие отрезки гибридной двойной спирали ДНК – РНК, которые необходимы для правильного считывания цепи ДНК. Как только РНК-полимераза достигнет терминирующих последовательностей, запускается последняя стадия – терминация. По ее завершении происходит освобождение вновь синтезированной цепи РНК (рис. 48).
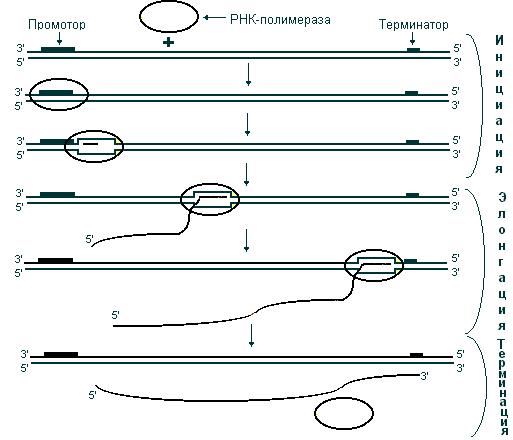
Рис. 48. Стадии транскрипции
Синтезированные молекулы РНК могут подвергаться посттранскрипционным ковалентным модификациям, так называемому процессингу (созреванию). Наиболее ярко процессинг выражен у эукариот.
Процессинг РНК
Рассмотрим процессинг иРНК эукариот.
иРНК синтезируется в виде предшественника пре-иРНК. Модификация пре-иРНК начинается на стадии элонгации, в это время происходит кэпирование 5’-конца. Остаток ГТФ присоединяется своим 5’-концом к 5’-концу пре-иРНК с образованием так называемого кэпа. После завершения транскрипции к 3’-концу присоединяется полиА-последовательность, состоящая из 100 – 200 последовательно соединенных адениновых нуклеотидов. В составе пре-иРНК могут содержаться интроны и экзоны. Интроны при созревании РНК вырезаются, концы экзонов соединяются друг с другом. Этот процесс называется сплайсингом. В результате сплайсинга зрелая молекула иРНК становится приблизительно в 4 раза короче пре-иРНК. Этапы процессинга иРНК представлены на рис. 49. Процессинг для иРНК прокариот не характерен.
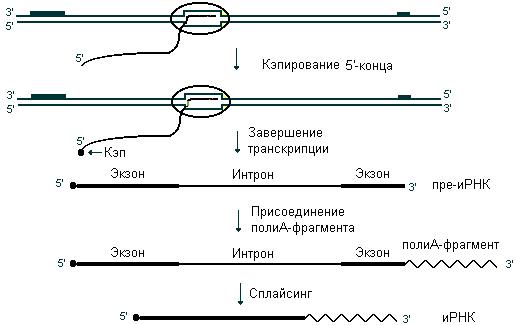
Рис. 49. Процессинг иРНК
рРНК и тРНК также синтезируются в виде более длинных предшественников, которые затем расщепляются и модифицируются. У прокариот рРНК представлены тремя молекулами: 16S-рРНК, 23S-рРНК и 5S-рРНК, у эукариот – четырьмя: 18S-рРНК, 28S-рРНК, 5,8S-рРНК и 5S-рРНК. рРНК как прокариот, так и эукариот образуются из предшественников – пре-рРНК. Предшественник расщепляется, образуя индивидуальные рРНК (рис. 50). рРНК и тРНК не содержат кэпа и полиА-последовательности. У эукариот рРНК и тРНК могут подвергаться сплайсингу. В процессе созревания у некоторых предшественников тРНК наряду с удалением концевых последовательностей может происходить и присоединение нуклеотидных последовательностей, играющих важную роль в их функционировании.
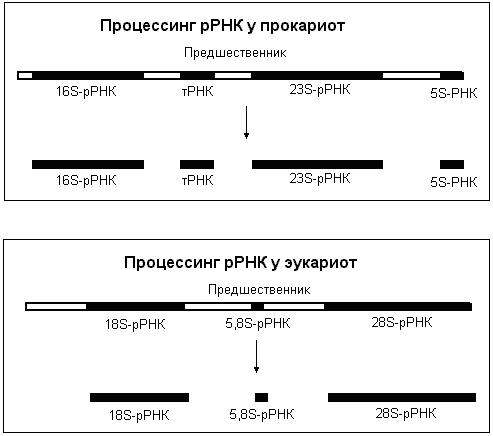
Рис.50. Процессинг рРНК.
Обратная транскрипция
Синтез ДНК при использовании в качестве матрицы РНК носит название обратной транскрипции. Данный процесс катализирует фермент обратная транскриптаза или ревертаза. Существование обратных транскриптаз в составе РНК-содержащих вирусов было показано Г.Темином и Д. Балтимором. Обнаружение обратной транскриптазы позволило ответить на вопрос: как генетическая информация РНК-содержащих вирусов может включиться в ДНК клетки-хозяина. Процесс обратной транскрипции, катализируемый ревертазой, и последующая интеграция генетического материала в геном клетки хозяина представлены на рис. 51. В процессе обратной транскрипции вначале образуется дуплекс РНК – ДНК, затем РНК в составе этого дуплекса разрушается, синтезированная цепь ДНК далее служит матрицей для синтеза второй цепи ДНК. На заключительной стадии происходит интеграция ДНК, синтезированной в результате обратной транскрипции, в ДНК клетки-хозяина. Обратные транскриптазы способны синтезировать ДНК, комплементарную самым различным РНК. Благодаря этой особенности ревертаза нашла широкое применение в научных исследованиях. С помощью обратной транскриптазы можно получить, например, искусственный ген, используя в качестве матрицы иРНК.
Читайте также: