
Виды рнк геномов вирусов
Химический состав вирусов отличается от других форм жизни необычайной простотой. Кроме геномной ДНК или РНК вирусы позвоночных содержат белки, масса которых составляет 57—90% массы вириона. Количество вирионных белков может колебаться в широких пределах в зависимости от сложности строения вируса. Среди белков, кодируемых вирусным геномом, различают структурные и неструктурные вирусспецифические белки. Первые входят в структуру вириона, вторые не входят. Структурными белками являются капсидные белки, белки оболочки и в некоторых случаях белки тегумента и ферменты.
У многих вирусов некоторые структурные вирусные белки не являются первичными продуктами генов, а образуются в результате посттрансляционного расщепления полипептида-предшественника вирусспецифическими и клеточными протеазами.
Неструктурными вирусными белками являются белки, кодируемые вирусным геномом, но не являющиеся составными частями вириона. Одни из них участвуют в репликации вирусных компонентов, другие — в сборке вирионов, третьи нарушают нормальный клеточный синтез. Они включают различные типы репликаз, связанные с репликацией ДНК, транскриптазы, транскрибирующие мРНК на геномной дцДНК или дцРНК или (-)оцРНК, вирусные протеазы, геликазы и лигазы. У ретро- и гепаднавирусов найдена обратная транскриптаза, транскрибирующая ДНК на матрице РНК. У ретровирусов найден уникальный фермент, участвующий в интеграции ДНК-продукта в хромосомальную ДНК клетки. Вирусы оспы, которые размножаются в цитоплазме и имеют много уникальных ферментов для процессинга РНК-транскриптов и репликации вирусной ДНК, мало зависят от метаболизма клетки. Вирусные белки имеют молекулярную массу 5-200 кД. Наиболее просто устроенные вирусы (вирусы-сателлиты, дефектные вирусы) кодируют синтез только одного белка, многие патогенные вирусы кодируют синтез 5—10 белков, крупные вирусы, такие как вирусы оспы, герпесвирусы, кодируют синтез до 200 белков. Хотя это немного по сравнению с клетками прои эукариотов (кодируют соответственно более 5000 и 100000 белков).
Разные вирусы демонстрируют различные варианты стратегии экспрессии своих генов и репликации геномов.
Все вирусные геномы являются гаплоидными, т.е. содержат одну копию каждого гена. Исключение составляют ретровирусы, которые обладают диплоидным геномом. Геномы ДНК-вирусов позвоночных представлены одной двуспиральной молекулой за исключением парво- и цирковирусов.
Геномы полиома-, папиллома-, гепадна- и цирковирусов представлены кольцевой ДНК. ДНК гепаднавирусов частично двуспиральная, частично односпиральная. ДНК вирусов полиомы и папилломы является суперспиральной. Большинство линейных вирусных ДНК обладает способностью приобрести циркулярную конфигурацию, которая требуется для репликации по вращающемуся кольцевому механизму. Две цепи ДНК вируса оспы ковалентно связаны своими концами и при денатурации образуют большое одноцепочечное кольцо. У некоторых ДНК-вирусов (так же как у РНК-ретровирусов) имеются концевые повторяющиеся последовательности. Инвертированные концевые повторы обнаружены у адено- и парвовирусов. У адено-, гепадна- и парвовирусов, так же как у некоторых РНК-вирусов (пикорна- и калицивирусов), с 5'-концом генома ковалентно связан белок, играющий важную роль в его репликации.
Все РНК-вирусы позвоночных за исключением рео- и бирнавирусов имеют одноцепочечные геномы. Геном некоторых РНК-вирусов состоит из нескольких (2-12) уникальных фрагментов, каждый из которых кодирует, как правило, один белок. РНК-вирусы с односпиральным геномом могут иметь различную полярность. Если они имеют ту же полярность, что и мРНК, то они могут прямо индуцировать синтез вирусного белка и считаются положительно (+) полярными.
Если геномная нуклеотидная последовательность комплементарна мРНК, то они считаются отрицательно (—) полярными. К ним относятся: парамиксо-, рабдо-, фило-, ортомиксо-, арена- и буньявирусы. Все они имеют вирионную РНК-зависимую полимеразу (транскриптазу), которая в инфицированной клетке транскрибирует положительно-полярную РНК на матрице геномной вирусной РНК. У аренавирусов, по крайней мере, у одного рода буньявирусов, один из РНК-сегментов является двуполярным. Обычно у (+)полярных РНК-вирусов З'-конец имеет polyA-последовательность, а 5'-конец имеет кэп-структуру.
Размер геномов РНК-вирусов (одноцепочечных 1,7—21 т.н.; двуцепочечных — 18—27 т.п.н.) значительно меньше размера генома многих ДНК-вирусов. Поэтому РНК-вирусы, как правило, кодируют меньше белков, чем ДНК-вирусы. Масса генома различных вирусов находится в пределах от 1 % (орто- и пара-миксовирусы) до 32% (парвовирусы) от массы вириона.
Различные семейства вирусов позвоночных значительно различаются по структуре и функции генома. Основные типы вирусных геномов можно представить следующим образом:
1) двуцепочечной линейной молекулой ДНК с открытыми (герпесвирусы, аденовирусы, иридовирусы) или ковалентно связанными концами (вирусы оспы, асфаровирусы);
2) одноцепочечной линейной молекулой ДНК (парвовирусы);
3) одноцепочечной кольцевой молекулой ДНК (цирковирусы);
4) двуцепочечной кольцевой молекулой ДНК (папилломавирусы, полиомавирусы);
5) частично двуцепочечной кольцевой незамкнутой молекулой ДНК (гепаднавирусы);
6) одноцепочечной молекулой РНК, являющейся мРНК (положительно-геномные вирусы: пикорнавирусы, тогавирусы, флавивирусы, астровирусы, калицивирусы, коронавирусы, артеривирусы, нодавирусы);
7) одноцепочечной единой (рабдовирусы, парамиксовирусы, филовирусы, бор-навирусы) или фрагментированнои (ортомиксовирусы) линейной молекулой РНК, комплементарной мРНК — отрицательно-геномные вирусы;
8) одноцепочечной фрагментированнои кольцевой ковалентно несвязанной отрицательной или двуполярной РНК (буньявирусы, аренавирусы);
9) двуцепочечной линейной фрагментированнои молекулой РНК (реовирусы, бирнавирусы);
10) двумя идентичными линейными молекулами плюс-РНК, являющимися матрицами для синтеза ДНК (ретровирусы).
Молекулярная масса ДНК различных вирусов позвоночных варьирует в широких пределах: от 0,7—1,5 МД у цирковирусов и парвовирусов, до 150—200 МД у вирусов оспы. Молекулярная масса генома у РНК вирусов колеблется менее значительно - от 2,0 до 20,0 МД.
Лекция 7
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ГЕНОМА ВИРУСА
Вирусы являются одним из излюбленных объектов молекулярной генетики благодаря простому строению и малой молекулярной массе их геномов, которая в 10 6 раз меньше массы генома эукариотической клетки. Организация генетического аппарата у ряда вирусов, например у SV40, настолько сходна с таковой генов эукариотической клетки, что получила название минихромосомы. Минихромосома широко используется для изучения организации и репликации ДНК.
Число генов у вирусов значительно варьирует: от 3—4 генов у просто устроенных вирусов (парвовирусы) до 150 генов и больше у сложно устроенных (вирус оспы). Геном вирусов животных является гаплоидным, за исключением ретровирусов, которые имеют диплоидный геном, представленный двумя идентичными молекулами РНК. У вирусов с фрагментарным геномом (вирусы гриппа, реовирусы) каждый фрагмент обычно представляет собой один ген.
Так же, как и геном эукариотической клетки, ДНК-геном ряда вирусов животных имеет мозаичную структуру, при которой смысловые последовательности чередуются с неинформативными последовательностями. Механизм сплайсинга при формировании иРНК широко распространен и среди вирусов, имеющих ядерную локализацию транскрипции (адено-, папова-, герпесвирусы), поскольку ферменты, осуществляющие сплайсинг, находятся в ядре. Однако сплайсинг был обнаружен и у РНК-содержащих вирусов. Например, у вирусов гриппа происходит сплайсинг транскриптов 7-го и 8-го генов; в результате сплайсинга и сдвига рамки трансляции продуктами каждого из этих генов являются по два уникальных белка.
В составе генов ДНК-содержащих вирусов есть регуляторные участки, в том числе промотор, контролирующие функцию структурных генов. Сильными промоторами являются концы многих вирусных ДНК, представляющие собой длинные концевые повторы, сильный промотор имеют гены тимидинкиназы вирусов оспы и герпеса. Эти промоторы используются в генной инженерии для усиления транскрипции изучаемого гена.
СПОСОБЫ УВЕЛИЧЕНИЯ ИНФОРМАЦИОННОЙ ЕМКОСТИ ВИРУСНОГО ГЕНОМА
У многих вирусов молекулярная масса синтезирующихся белков превышает теоретически рассчитанную. Этот феномен объясняется наличием у вирусов механизмов, позволяющих получить развернутую генетическую информацию при максимальной экономии генетического материала; подобные механизмы выработаны в процессе эволюции вирусов как генетических паразитов.
Способами увеличения генетической информации являются: 1) двукратное считывание одной и той же иРНК, но с другого инициирующего кодона; 2) сдвиг рамки трансляции; 3) сплайсинг; 4) транскрипция с перекрывающихся областей ДНК и др.
Трансляция может происходить без сдвига рамки и со сдвигом рамки. Генетический код является триплетным, это означает, что три нуклеотида, составляющих триплет, или кодон, кодируют одну аминокислоту. В том случае, если триплеты сохранены и генетический код не изменился, то при трансляции с двух разных инициирующих кодонов будут синтезироваться полипептиды, представляющие собой укороченную копию первого полипептида (трансляция без сдвига рамки).
В том случае, если произошел сдвиг на один или два нуклеотида, образуются новые триплеты (кодоны) и появляется новый генетический код. В этом случае одна молекула иРНК может транслироваться с образованием двух уникальных белков, т. е. таких белков, у которых нет идентичных аминокислотных последовательностей.
Сплайсинг со сдвигом рамки широко используется у ряда вирусов (вирусы гриппа, парамиксовирусы, буньявирусы, аденовирусы, паповавирусы, парвовирусы и др.). Например, все три иРНК аденоассоциированного вируса образуются при транскрипции одного гена и имеют общий З'-конец; самая короткая иРНК образуется путем сплайсинга и транслируется с образованием трех структурных белков, остальные две иРНК транслируются с образованием неструктурных белков. В результате сплайсинга и сдвига рамки иРНК 7-го и 8-го генов вируса гриппа транслируются с образованием двух белков: полипептидов М, и М2 (продукты 7-го гена) и NSi и NS2 (продукты 8-го гена). Белки NSt и NS2 имеют лишь первые 10 идентичных аминокислот, а затем — уникальные аминокислотные последовательности. Один и тот же ген парамиксовирусов (вирус Сендай) кодирует два уникальных белка: структурный белок Р и неструктурный белок С.
Одним из способов экономии генетического материала является нарезание полипептида-предшественника на участки разной длины, в результате чего образуются разные полипептиды с перекрывающимися аминокислотными последовательностями. Подобный механизм нарезания имеет место у аденоассоциированных вирусов и у SV40.
Таким образом, число реальных генов превосходит молекулярную массу генома. Основанный на длине генома расчет числа генов неизменно приведет к ошибочным результатам. Более точные представления о числе генов можно получить путем биохимического и генетического анализов.
ОСНОВНЫЕ ПРОЦЕССЫ, КОНТРОЛИРУЮЩИЕ НАСЛЕДСТВЕННОСТЬ И ИЗМЕНЧИВОСТЬ ВИРУСОВ
Модификации. Модификациями называются не наследуемые (фенотипические) изменения у вирусов, обусловленные клеткой-хозяином. Эти изменения лежат в основе адаптации вируса к новому хозяину и преодоления зависимого от хозяина ограничения. Модификации нуклеиновых кислот вирусов осуществляют клеточные ферменты, ответственные за ограничение (рестрикцию) репродукции вируса.
Мутации. В основе изменчивости вирусов лежат мутации, т. е. изменения состава и последовательностей нуклеотидов вирусного генома. Мутации происходят у всех вирусов, независимо от того, является ли их генетическим аппаратом ДНК или РНК. В результате мутаций отдельные вирионы могут приобретать новые свойства. Дальнейшая судьба таких вирусов зависит от естественного отбора, сохраняющего популяцию, наиболее приспособленную к условиям существования.
Мутации могут иметь разные последствия. В одних случаях они ведут к изменению фенотипических проявлений в нормальных условиях. Например, увеличивается или уменьшается размер бляшек под агаровым покрытием; увеличивается или ослабляется нейровирулентность для определенного вида животных; вирус становится более чувствительным к действию химиотерапевтического агента и т. п.
В других случаях мутация является летальной, так как вследствие ее нарушается синтез или функция жизненно важного вирусспецифического белка, например вирусной полимеразы.
В некоторых случаях мутации являются условно летальными, так как вирусспецифический белок сохраняет свои функции в определенных, оптимальных для него, условиях и теряет эту способность в неразрешающих (непермиссивных) условиях. Типичным примером таких мутаций являются температурно-чувствительные (temperature sensitive) — ts-мутации, при которых вирус теряет способность размножения при повышенных температурах (39—42° С), сохраняя эту способность при обычных температурах выращивания (36—37° С).
По своему механизму мутации могут быть тоже разными. В одних случаях происходит делеция, т. е. выпадение одного или нескольких нуклеотидов, в других случаях происходит встраивание одного или нескольких нуклеотидов, а в некоторых случаях — замена одного нуклеотида другим.
Мутации могут быть прямыми и обратными. Прямые мутации меняют фенотип, а обратные мутации — реверсии — его восстанавливают. Возможны истинные реверсии, когда обратная мутация происходит в месте первичного повреждения, и псевдореверсии, если мутация происходит в другом участке дефектного гена (интрагенная супрессия) или в другом гене (экстрагенная супрессия). Реверсия не является редким событием, так как ревертанты обычно более приспособлены к данной клеточной системе. Поэтому при получении мутантов с заданными свойствами, например вакцинных штаммов, приходится считаться с возможной их реверсией к дикому типу.
Мутации носят случайный характер и объясняются статистическими законами.
В качестве физических мутагенов наиболее часто применяется ультрафиолетовое облучение, так как его энергия сопоставима с энергией химических связей. Реже применяются более жесткие виды облучения — рентгеновское и γ-облучение, а также обработка вирусных суспензий нейтронами, протонами, электронами и ядрами гелия, так как они вызывают сильные разрушения вирусных геномов и их инактивацию.
В качестве химических мутагенов применяют аналоги оснований (бромурацил, бромдезоксиуридин, 2-аминопурин, нитрозогуанидин и пр.), алкилирующие и флуоресцирующие соединения (профлавин), интеркалирующие агенты (актиномицин, этидий бромид), азотистую кислоту, гидроксиламин и многие другие.
ГЕНЕТИЧЕСКИЕ И НЕГЕНЕТИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ МЕЖДУ ВИРУСАМИ
Как в естественных, так и в экспериментальных условиях одна клетка может быть заражена не одним, а несколькими вирусами. В процессе такой смешанной инфекции могут иметь место различные формы взаимодействия как между вирусными геномами, так и между продуктами генов. При взаимодействии геномов могут наблюдаться такие формы генетических взаимодействий, как множественная реактивация, рекомбинация, пересортировка генов, кросс-реактивация, гетерозиготность. При взаимодействии на уровне продуктов генов могут иметь место негенетические взаимодействия: комплементация, интерференция, фенотипическое смешивание и др.
Множественная реактивация. Вирусная инфекция может возникнуть при заражении клетки несколькими ви-рионами с поврежденными геномами вследствие того, что функцию поврежденного гена может выполнять вирус, у которого этот ген не поврежден. Этот феномен был вначале обнаружен на бактериофагах и получил название множественной реактивации. В основе множественной реактивации лежит кооперативный процесс, при котором вирионы с поражением разных генов дополняют друг друга путем генетической рекомбинации, в результате чего репродуцируется исходный неповрежденный вирус. Эффективность множественности реактивации зависит от многих причин: степени повреждения генома вирионов, числа проникших в клетку вирионов, концентраций их в определенных участках клетки, аутоинтерференции поврежденных вирионов. Для множественной реактивации важное значение имеет расстояние между вирионами с поврежденными геномами внутри клетки. Обработка вирионов двухвалентными ионами металлов, ведущая к их агрегации, усиливает множественную реактивацию.
Рекомбинация. Генетической рекомбинацией называют обмен генетическим материалом, происходящий между родительскими вирусами. Возможен обмен полными генами (межгенная рекомбинация), так и участками одного и того же гена (внутригенная рекомбинация). Образующийся вирус-рекомбинант обладает свойствами, унаследованными от разных родителей.
Обычно рекомбинируемые штаммы обладают характерными признаками, которые обозначаются как маркеры. Например, были получены рекомбинанты между вирусами полиомиелита, обладающие повышенной устойчивостью и повышенной чувствительностью к гуанидину, разной нейровирулентностью, разной устойчивостью к повышенной температуре, разной чувствительностью к ингибиторам сывороток лошадей и коров и т. п. Для получения рекомбинантов используют штаммы, содержащие два или большее число маркеров.
Тест рекомбинации применяют для генетических исследований вирусов. С его помощью возможно построение генетических карт вирусов, в которых определяется, в каких участках генома произошли мутации, а также в условных единицах измеряется расстояние между разными мутациями.
Пересортировка генов. Вариантом рекомбинации является феномен, получивший название пересортировки генов. Она наблюдается при генетических взаимодействиях между вирусами, имеющими сегментированный геном. Образующиеся при этом гибридные формы вирусов называют реассортантами. Реассортанты вирусов гриппа получают при совместном культивировании вирусов с разными генами гемагглютинина и нейраминидазы. В этом случае из общего потомства путем нейтрализации соответствующих антигенов можно выделить интересующие исследователя варианты.
Существуют определенные группировки (констелляции или созвездия) генов, которые в данной системе клеток более стойки и делают вирус более жизнеспособным.
Сходные процессы пересортировки генов имеют место у вирусов гриппа типов А, В и С и у других вирусов с фрагментарным геном — у буньявирусов, аренавирусов (однонитчатые РНК) и реовирусов (ротавирусов) (двунитчатая РНК). Однако эти процессы не столь интенсивны и доступны изучению, как у вирусов гриппа.
Гетерозиготность. При совместном культивировании двух штаммов вируса может происходить формирование вирионов, содержащих в своем составе два разных генома или по крайней мере один полный геном и часть второго генома. Это явление названо гетерозиготностью.
Комплементация. Комплементация (дополнение) является таким видом негенетического взаимодействия при смешанной инфекции двумя вирусами, которое стимулирует репродукцию обоих партнеров или одного из них, но не изменяет генотипы вирусов. Принцип комплементации заключается в том, что вирус снабжает партнера недостающими компонентами, обычно белками, структурными или неструктурными.
Комплементация широко распространена среди вирусов и встречается как между родственными, так и неродственными вирусами. Феномен тесно связан с проблемой дефектности вирусов.
Поскольку в вирусной популяции помимо стандартных обычно присутствуют дефектные неинфекционные вирусные особи, в частности дефектные частицы, утратившие часть генетического материала, комплементация имеет место в инфекционном цикле многих вирусов и заключается в том, что члены популяции снабжают друг друга продуктами генов, которые дефектны у партнеров. Отличие комплементации от генетической рекомбинации заключается в отсутствии обмена генетическим материалом.
Комплементация встречается и у неродственных вирусов, принадлежащих к разным семействам. Одним из семейств, вирусы которого наиболее часто участвуют в комплементации, является семейство аденовирусов. В одних системах аденовирусы могут действовать как дефектные вирусы, в других — как помощники. Например, в культуре клеток почек макак резусов аденовирусы могут репродуцироваться только в присутствии SV40, который является в данном случае вирусом-помощником. В других системах сами аденовирусы действуют как вирусы-помощники, а вирусом-сателлитом является аденоассоци-ированный вирус, относящийся к семейству парвовирусов. Репродукция этого вируса полностью зависит от комплементирующего действия аденовирусов. Вирус гепатита В является помощником для дельта-агента, который покрывается его наружным белком — HBs-антигеном. Сочетание обоих вирусов обнаружено при наиболее тяжелых формах гепатита.
Возможна не только межцистронная, но и внутрицистронная комплементация в том случае, когда один ген кодирует несколько белков.
Фенотипическое смешивание. При совместном культивировании двух вирусов может наблюдаться феномен фенотипического смешивания, когда геном одного вируса бывает заключен в капсид, состоящий частично или полностью из белков другого вируса.
Фенотипическое смешивание наблюдается при смешанной инфекции,многими вирусами, причем эти вирусы могут быть как близкими друг другу (например, вирусы гриппа А и В или разные серологические подтипы вируса гриппа А), так и весьма далекими (онковирусы и рабдовирусы).
РЕСТРИКТАЗЫ И ФИЗИЧЕСКИЕ КАРТЫ ВИРУСОВ
Подлинную революцию в физическом картировании геномов вирусов произвело применение рестриктаз и секвенирование вирусных геномов. Рестриктазы имеют исключительное значение в молекулярной генетике вообще и генетической инженерии в частности. Их открытие (1968—1970 гг.) впервые дало возможность специфически расщеплять ДНК на строго определенные фрагменты, доступные для препаративного выделения и анализа.
Рестриктазы или эндодезоксирибонуклеазы — это просто организованные белки, являющиеся ферментами, широко распространенными среди прокариотов и участвующими в генетических процессах. В отличие от экзонук-леаз, отщепляющих концевые нуклеотиды или свободные остатки фосфорной кислоты, эндонуклеазы расщепляют молекулу ДНК изнутри, обычно — в местах, где преобладают пиримидиновые основания. Рестриктазы характеризуются высоковыраженной специфичностью, распознавая строго определенные последовательности нуклеотидов в двунитчатой ДНК.
Число новых рестриктаз стремительно нарастает и со временем, по-видимому, будут обнаружены рестриктазы, узнающие любую последовательность нуклеотидов.
Использование разных рестриктаз позволяет получать фрагменты разной величины, которые затем разделяются и анализируются путем электрофореза в агарозных или полиакриламидных гелях. Сочетание рестрикционного анализа с другими методами позволяет составить физические карты геномов вирусов. Физические карты вирусных геномов обозначают взаимное расположение генов, их границы, локализацию начала репликации, промоторов, лидеров, экзонов и интронов, сигнальных последовательностей и других генетических элементов.
Генетический код для синтеза белков вируса SV-40 записан не на одной, а на обеих нитях ДНК, а транскрипция разных генов идет в разных направлениях.
В настоящее время полностью расшифрованы нуклеотидные последовательности отдельных генов и целых геномов методом секвенирования (от англ. sequence — последовательность). Если речь идет о РНК-содержащих вирусах, то предварительным условием для дальнейшего их анализа является переписка РНК на ДНК с помощью РНК-зависимой ДНК-полимеразы (обратной транскриптазы), после чего генетический материал может быть подвергнут рестрикционному анализу.
Проектное задание к модулю
В качестве проектного задания студентам предлагается написание рефератов по следующим темам:
1. Природа дефектных вирусных геномов. Вирусы-сателлиты.
2. Вирусная интерференция
3. Необычные свойства ретровирусов
4. Трансформация клетки опухолеродными ДНК-вирусами
5. Классификация и основные свойства вирусов гриппа
6. Индукция специфического иммунного ответа на вирусы
7. Особенности репродукции пикорнавирусов
8. Семейство тогавирусов. Особенность репродукции и инфекционного процесса.
9. Вирус клещевого энцефалита.
10. Вирус бешенства
11. Вирусная персистенция
12. Основные свойства парамиксовирусов
13. Вирус кори. Биология возбудителя. Особенности патогенеза
14. Аденовирусы, их репликация и связь с другими вирусами
16. Пути распространения вирусных болезней растений.
17. Прионы. Возбудители или провокаторы или….
18. Вироиды как вирусоподобные инфекционные агенты.
19. Вирусы, вызывающие респираторные инфекции. Сравнительная характеристика
20. Место вирусов в биосфере.
21. Бактериофаги. Их морфологическое многообразие и взаимодействие с бактериями.
22. Атипичная пневмония и возможность происхождения новых вирусов
23. Роль вирусов в возникновении злокачественных опухолей
24. Особенности транскрипции РНК- содержащих вирусов.
25. Морфогенез вирусов или морфологические превращения в процессе репродукции.
Тест рубежного контроля
Геном вирусов, заключенный внутри вирионов, может быть представлен одноцепочечными или двухцепочечными ДНК или РНК.
Гены вирусов могут быть заключены в одной хромосоме или разделены на несколько блоков (хромосом), которые все вместе и составляют геном таких вирусов.
Геном РНК-содержащих вирусов представлен только линейными молекулами РНК. Все известные ДНК- содержащие вирусы позвоночных имеют геном, заключенный в одной хромосоме, линейной или кольцевой, одно- или двухцепочечной.
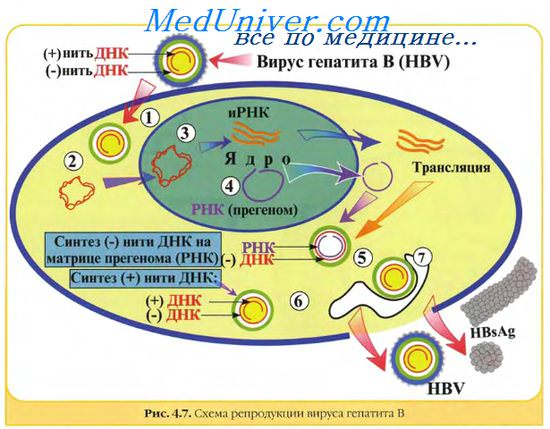
У некоторых вирусов, например, у вируса гепатита В, геном представлен кольцевой ковалентно замкнутой молекулой двухцепочечной ДНК, в обеих цепях которой в разных местах обнаружены одноцепочечные участки. У нескольких родов, например, адено-ассоциированных вирусов, комплементарные цепи ДНК находятся в различных вирусных частицах.
Геном вирусов включает:
- · Структурные гены. Занимают примерно 95 % вирусной хромосомы. Белки вирусов можно разделить на несколько групп: структурные, ферменты, регуляторы.
- · Регуляторные последовательности, которые не кодируют белки: промоторы, операторы и терминаторы.
- · Прочие некодирующие участки (сайты), в том числе:
участок attP, обеспечивающий интеграцию вирусной хромосомы в хромосому клетки-хозяина;
участки cos - липкие концевые участки линейных вирусных хромосом, обеспечивающие замыкание линейной хромосомы в кольцевую форму.
· Гены, кодирующие рРНК и тРНК, в геноме вирусов обычно отсутствуют (за исключением, генома о фага Т 4, который имеет гены, кодирующие тРНК).
Геном вирусов отличается высокой плотностью упаковки информации. Например, у фага ц Х 174 в пределах одного гена может располагаться еще один ген (на рисунке кольцевая ДНК представлена в линейной форме). В частности, ген В находится в пределах гена А, а ген Е - в пределах гена D:

У мелкого РНК-содержащего фага f2 ген регуляторного белка, блокирующего лизис (созревание вирионов и разрушение клетки), перекрывается с двумя другими генами, удаленными друг от друга:
Экспрессия (транскрипция и трансляция) вирусных генов происходит в том случае, если геном вируса представлен двунитевой ДНК (у РНК-содержащих вирусов необходим перевод информации в ДНК). Из-за полярности ДНК транскрипция идет только в одном направлении, то есть ген имеет начало и конец. Тогда "правые" гены не будут транскрибироваться РНК-полимеразой, движущейся влево, и наоборот. При этом один и тот же ген может транскрибироваться с разных промоторов; в этом случае экспрессия генов контролируется разными механизмами.
Особенности вирусов эукариот
У вирусов эукариот обнаружены следующие особенности:
- 1. Перекрывание генов (обезьяний вирус SV 40, вирус гриппа).
- 2. Интрон-экзонная структура генов.
- 3. Модификация белков после синтеза полипротеинов: весь геном транскрибируется в виде одной молекулы иРНК, которая служит матрицей для синтеза полипротеина - одного гигантского инертного белка, и лишь затем происходит расщепление полипротеина на белки, выполняющие определенные функции.
К ДНК-содержащим вирусам относятся многие вирусы бактерий - бактериофаги (или просто фаги). Некоторые мелкие фаги (например, фаг М 13) при репродукции не разрушают клетку. Репродукция крупных фагов (например, фага Т-4) приводит к гибели клетки. Фаг Т-4 - это один из наиболее сложно организованных вирусов. Белковый капсид включает не менее 130 белков, образующих головку, воротничок, сократимый хвост, базальную пластинку и хвостовые нити. Такое строение капсида позволяет впрыскивать ДНК в бактериальную клетку через толстую оболочку, поэтому подобные вирусы образно называют "живыми шприцами". Т-фаги могут существовать в виде профага длительное время. К ДНК-содержащим вирусам относятся возбудители многих заболеваний человека и животных: вирусы оспы, герпеса, гепатита В, аденовирусы млекопитающих и человека (вызывают желудочно-кишечные заболевания, ОРВИ, конъюнктивиты), вирусы бородавок человека. К ДНК-содержащим вирусам относятся и некоторые вирусы растений (вирус золотистой мозаики фасоли, вирус мозаики цветной капусты). Некоторые вирусы используются в генной инженерии для переноса генов от одних организмов к другим, например, обезьяний вирус SV 40.
Вирионы ДНК-содержащих вирусов содержат ДНК. Объемом ДНК определяется количество белков в вирионе: один полипептид кодируется отрезком ДНК длиной примерно 1 тысяча нуклеотидов (нуклеотидных пар). После проникновения в клетку вирусная ДНК становится матрицей для синтеза ДНК и РНК.
Примеры организации генома ДНК-содержащих вирусов
1. Кольцевая двухцепочечная ДНК длиной около 5 тпн.
ь Обезьяний вирус SV 40. Мелкий эукариотический вирус. Вирионы в виде икосаэдра. Капсид белковый. Используется в генной инженерии как вектор переноса генов. Кодирует 5 белков.
ь Вирусы бородавок человека.
2. Кольцевая одноцепочечная ДНК длиной около 5 тн; может быть как кодирующей, так и антикодирующей.
Мелкие бактериофаги типа М 13. Не разрушают клетку. Капсид включает 8 белков
Вирус золотистой мозаики фасоли.
3. Линейная двухцепочечная ДНК длиной 30-150 тпн.
Бактериофаги типа Т 4. Вирионы крупные. Белковый капсид из 130 белков включает: головку, хвостовой отдел и хвостовые нити. Эти вирусы могут существовать в виде профага длительное время.
Аденовирусы млекопитающих и человека. Вирионы средних размеров в виде икосаэдра. Капсиды белковые. Вызывают ОРВИ, конъюнктивиты, желудочно-кишечные заболевания, иногда обладают онкогенными свойствами.
Вирусы оспы, герпеса и им подобные. Вирионы крупные. Имеется липопротеиновая оболочка.
- 4. Линейная одноцепочечная ДНК длиной около 5 тн; ДНК может быть как кодирующей, так и антикодирующей. У человека известны как спутники аденовирусов.
- 5. Двухцепочечная ДНК, замкнутая в кольцо из перекрывающихся сегментов. Длина ДНК - 3-8 тн.
ь Вирус гепатита В. Вирион сферический, средних размеров. Имеется дополнительная оболочка из вирусных и клеточных белков. Кодирует 5 белков.
ь Вирус мозаики цветной капусты.
К РНК-содержащим вирусам относятся многие вирусы растений, возбудители заболеваний человека и животных: вирус полиомиелита, вирусы гриппа А, В и С, вирусы паротита (свинки), кори, чумы плотоядных животных (чумки), бешенства, вирус иммунодефицита человека (ВИЧ). В отдельную группу выделяются арбовирусы, которые переносятся членистоногими (клещами, москитами), например, вирусы клещевого энцефалита, желтой лихорадки. Многие РНК-содержащие вирусы вызывают ОРВИ (например, коронавирусы), желудочно-кишечные заболевания (реовирусы птиц, млекопитающих и человека). Некоторые РНК-содержащие вирусы используются в биотехнологии, например, вирусы полиэдроза насекомых.
Вирионы РНК-содержащих вирусов содержат РНК. После проникновения в клетку вирусная РНК становится матрицей для синтеза ДНК и РНК.
Примеры организации генома РНК-содержащих вирусов
1. Линейная одноцепочечная мРНК (плюс-цепь) длиной около 4 тн; в виде единой молекулы или в виде нескольких разных молекул. Плюс-цепь сразу же может использоваться для трансляции. Вегетативно-репродуктивная фаза этих вирусов протекает в цитоплазме. В плюс-цепи закодирована РНК-репликаза (РНК-зависимая РНК-полимераза). Представители:
ь Вирус табачной мозаики (ВТМ) - сегментированная РНК. Вирион нитевидный (18х 300 нм). ВТМ открыт Д.И. Ивановским в 1982 г.
ь Вирус полиомиелита - несегментированная РНК. Вирионы мелкие, в виде икосаэдра. Капсид белковый.
ь Вирус бешенства. Нитевидный вирион. Имеется дополнительная липопротеиновая оболочка.
ь Арбовирусы (переносятся членистоногими: клещами, москитами) - вирусы клещевого энцефалита, желтой лихорадки. Морфология и размеры вирионов разнообразны, например, вирус энцефалита содержит 9 белков. Имеется дополнительная липопротеиновая оболочка.
ь Мелкие бактериофаги (с несегментированной РНК).
2. Линейная одноцепочечная кРНК (минус-цепь, порядок нуклеотидов комплементарен по отношению к мРНК). Минус-цепь не может служить для трансляции и используется как матрица для синтеза плюс-цепи. Плюс-цепь служит для трансляции вирусных белков и используется как матрица для синтеза вирусной кРНК. Вегетативно-репродуктивная фаза этих вирусов также протекает в цитоплазме.
ь Вирусы гриппа А, В, С. Вирус гриппа А содержит минус-цепь РНК, состоящую из 8 фрагментов. Фрагменты РНК связаны с вирусными белками и образуют спиральный нуклеокапсид. Поверх нуклеокапсида располагается гликолипопротеиновый суперкапсид. В составе вириона 10 белков. В состав суперкапсида входит два белка, определяющих антигенные свойства вируса: гемагглютинин и нейраминидаза. Кроме того, в состав вириона входит уже готовая РНК-репликаза, обеспечивающая синтез плюс-цепи на матрице минус-цепи.
ь Вирусы паротита (свинки), кори, чумы плотоядных животных (чумки). Сферический вирион средних размеров. Имеется дополнительная липопротеиновая оболочка.
- 3. Линейная двухцепочечная РНК
- 1. Мелкие бактериофаги. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый.
- 2. Вирусы полиэдроза насекомых. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Используются в биотехнологии (для синтеза интерферона).
ь Реовирусы птиц, млекопитающих и человека. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Вызывают ОРВИ, желудочно-кишечные заболевания. РНК фрагментированная (10. 11 фрагментов), кодирует 11 белков.
4. Две линейные одноцепочечные одинаковые молекулы мРНК длиной около 10 тн
Ретровирусы. Способны интегрироваться в ДНК. В состав вирионов входит фермент обратная транскриптаза (ревертаза). Имеется дополнительная липопротеиновая оболочка. Многие ретровирусы вызывают онкологические заболевания: лейкозы, саркомы, опухоли молочных желез. К ретровирусам относится и вирус иммунодефицита человека, вызывающий СПИД.
ь Вирус иммунодефицита человека (ВИЧ). Содержит одну плюс-цепь РНК, кодирует 13 белков. Сферический вирион. Имеется дополнительная липопротеиновая оболочка, включающая фрагменты мембран человека. Избирательно поражает Т-лимфоциты.
Читайте также: