
Нервно мышечные и нервно сухожильные веретена
13.1. Классификация нервных окончаний
а) Все нервные волокна заканчиваются нервными окончаниями.
б) Нервные окончания (н.о.) можно разделить на 4 группы. -
б) Обнаружены также
Далее, в соответствии с данной классификацией, мы рассмотрим
13.2. Рецепторные нервные окончания
13.2.1. Классификация рецепторов
Рецепторные нервные окончания, в свою очередь, тоже можно подразделить на разные виды, причём, - несколькими способами.
2. несвободные нервные окончания (вокруг осевого цилиндра сохраняются клетки глии) -
Рецепторные нервные окончания имеются во всех органах.
Познакомимся с некоторыми их конкретными видами.
13.2.2. Рецепторы в эпителии кожи
2. а) Одни из них просто проникаю т между клетками эпителия.
б) Другие контактирую т с основаниями осязательных эпителиоцитов (специфически изменённы х эпителиальны х клет ок ).
13.2.3. Рецепторы в соединительной ткани
13.2.3.1. Общие сведения
Рассмотрим эти два вида рецепторных окончаний на препаратах.
13.2.3.2. Осязательные (мейснеровы) тельца
б) Так, на снимке мы видим

б) Характерна их ориентация -
13.2.3.3. Пластинчатые (фатер-пачиниевы) тельца

б) Вследствие такого её строения, пластинчатое тельце воспринимает
13.2.4. Рецепторы в скелетных мышцах и сухожилиях
2. Они представляют собой особые структуры -
13.2.4.1. Нервно-мышечные веретёна: основные сведения
центральные части интрафузальных волокон;
от гамма-мотонейронов спинного мозга.
б) Поэтому при сокращении концевых отделов (под влиянием эфферентных нервных волокон)
13.2.4.2. Нервно-мышечные веретёна: дополнения
I. Виды интрафузальных мышечных волокон
Различают 2 вида интрафузальных мышечных волокон.
II. Виды афферентных нервных волокон
Чувствительные нервные волокна, иннервирующие нервно-мышечные веретёна, тоже бывают двух видов.
13.2.4.3. Нервно-сухожильные веретёна
б) При мышечном сокращении происходит
в) По рефлекторной цепи это приводит
13.3. Синапсы (межнейронные и нейроэффекторные)
13.3.1. Общие сведения
13.3.1.1. Составные части синапса
2. В синапсе различают
13.3.1.2. Синапсы химического и электрического типа
13.3.1.3. Подразделение синапсов по виду сигнала и по природе медиатора
2. Иногда синапсы называют по природе медиатора -
3. Большинство медиаторов оказывает возбуждающее действие на постсинаптическую клетку;
примеры же медиаторов тормозного действия -
13.3.1.4. Функционирование синапса химического типа
13.3.2. Межнейронные синапсы
б) А аксоаксональные синапсы бывают только тормозного типа.
в) Спектр медиаторов, используемых в межнейронных синапсах, весьма широк:
б) К телу нейрона подходят многочисленные аксоны (2) других нейронов, расширяющиеся в пресинаптической части.

13.3.3. Нейроэффекторные синапсы
(эффекторные нервные окончания)
В этих синапсах сигналы (приходящие по аксону) передаются на эффекторные органы -
13.3.3.1 Нервно-мышечные окончания:
общее описание
а) Подходя к мышечному волокну, аксон (4)
б) Последние погружаются в мышечное волокно вместе с прогибающейся сарколеммой , но
в) В пресинаптических окончаниях содержится много
г) Для синтеза медиатора используется фермент ацетилхолинсинтетаза.
б) Она имеет многочисленные инвагинации,
что увеличивает площадь её контакта с медиатором.
в) В этой мембране находятся два ключевых белка -
г) В подлежащей саркоплазме наблюдается скопление
13.3.3.2. Нервно-мышечные окончания: просмотр препарата


б) В области контактов терминалей с мышечным волокном - ядра (4)
Мышечные веретена состоят из интрафузальных мышечных волокон, количество которых не превышает 12. (В данном контексте экстрафузалъные мышечные волокна считают обычными). Крупные интрафузальные волокна начинаются от полюсов (концов) мышечных веретен и фиксируются к соединительной ткани — перимизию. Некрупные интрафузальные волокна фиксируются к коллагеновой капсуле веретена. В средней части (экваторе) веретена саркомеры практически полностью замещены ядрами, расположенными в форме сумки—сумчато-ядерные мышечные волокна (характерно для широких волокон) или цепочки — цепочечно-ядерные мышечные волокна (характерно для тонких волокон).
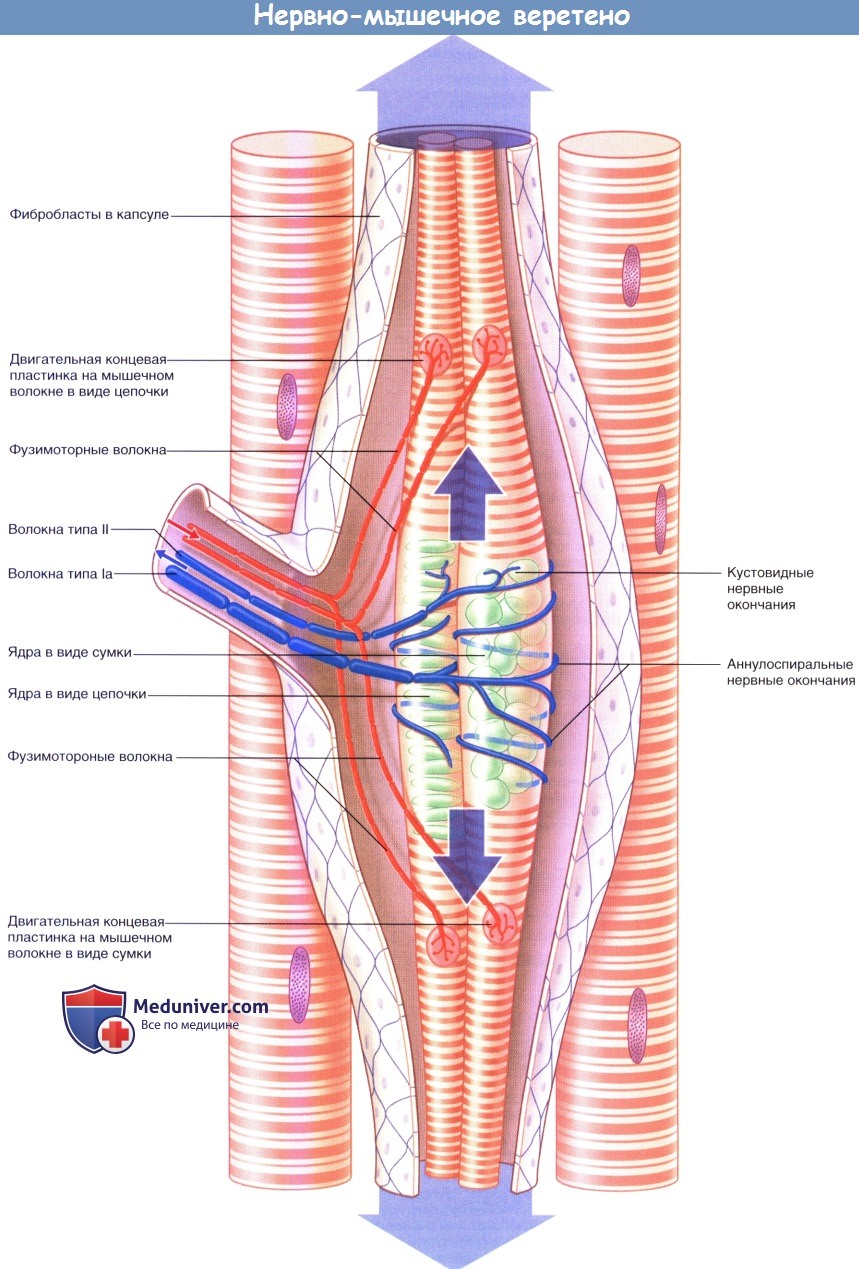
Упрощенное строение нервно-мышечного веретена. Большими стрелками обозначено пассивное растяжение аннулоспиральных нервных окончаний при удлинении мышцы в состоянии покоя.
Стрелками среднего размера обозначено активное растяжение аннулоспиральных нервных окончаний фузимоторными нервными волокнами.
Активное растяжение в достаточной степени компенсирует эффект разгрузки одновременного сокращения экстрафузальных мышечных волокон.
Стрелками маленького размера показаны направления проведения импульсов к мышечному веретену и от него при сокращении мышцы.
1. Иннервация. Иннервацию мышечных веретен осуществляют двигательные и чувствительные нервные волокна. Двигательные нервные волокна — фузимоторные—в соответствии с толщиной относят к типу Аγ, а нервные волокна экстрафузальных мышц — к типу Аα. Фузимоторные аксоны разветвляются и иннервируют поперечно-исчерченные сегменты концевых участков интрафузальных мышц. Единичные первичные чувствительные нервные волокна типа 1а оборачиваются спирально-кольцевым образом вокруг сумчато-ядерных или цепочечно-ядерных мышечных волокон. Вторичные кустовидные чувствительные нервные окончания образованы нервными волокнами II типа и располагаются с одной или с двух сторон от первичного волокна.
2. Активация. Мышечные веретена представляют собой рецепторы растяжения. При растяжении мышечного веретена ионные каналы поверхностной мембраны чувствительных нервных окончаний открываются, за счет чего создаются положительные электроволны. В результате их суммирования в области терминального участка аксона чувствительного нервного волокна происходит образование рецепторного потенциала, который при достижении пороговых значений генерирует нервные импульсы.
Растяжение мышечных веретен может происходить пассивным и активным путями.
3. Пассивное растяжение. Пассивное растяжение мышечного веретена происходит при пассивном удлинении всего мышечного брюшка. Например, при воспроизведении сухожильных рефлексов, в частности коленного, во время удара по сухожилию происходит пассивное растяжение мышечных веретен брюшка четырехглавой мышцы бедра. Волокна типов Iа и II направляются к спинному мозгу и образуют синапсы с дендритами α-мотонейронов. (Принадлежность мотонейронов к типу а определяется Аα-диаметром их аксонов.)
Сухожильные рефлексы относят к моносинаптическим. Для них характерен латентный период (промежуток времени до ответной реакции на стимул), составляющий 15-25 мс.
Помимо возбуждающего действия на гомонимные мотонейроны (т. е. двигательные нейроны, иннервирующие одни и те же мышцы), афферентные волокна мышечных веретен опосредованно через вставочные тормозные мотонейроны оказывают тормозное действие на а-мотонейроны мышц-антагонистов. Этот процесс получил название реципрокного торможения. Задействованные тормозные нейроны называют вставочными нейронами типа Iа.
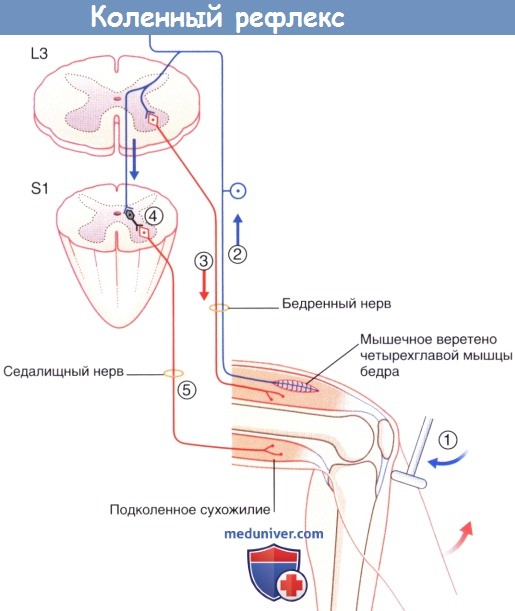
Коленный рефлекс, в котором задействовано реципрокное торможение.
Стрелочками указано проведение нервных импульсов.
(1) Удар по коленной связке вызывает растяжение мышечных веретен в четырёхглавой мышце спины.
(2) Мышечные веретена посылают возбуждающий потенциал в спинной мозг.
(3) α-Мотонейроны вызывают подергивание четырехглавой мышцы и разгибание колена.
(4 и 5) Тормозные вставочные нейроны типа Ia полностью подавляют активность подколенного сухожилия.
4. Кодирование информации. Первичные афференты мышечных веретен проявляют наибольшую активность во время растяжения. Они определяют скорость растяжения мышцы: чем быстрее происходит растяжение, тем больше импульсов они генерируют.
Вторичные афференты мышечных веретен по сравнению с первичными активнее задействованы в процессе поддержания определенной позиции и определяют степень растяжения мышцы: чем выше степень статического растяжения, тем больше формируется импульсов.
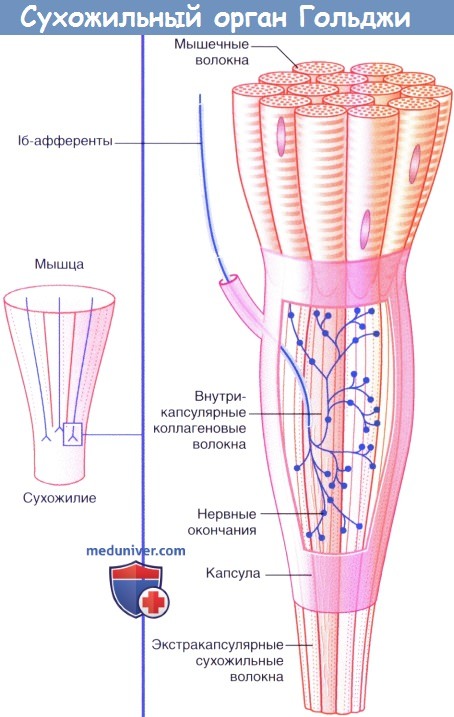
б) Нервные окончания сухожилий. Сухожильные органы Гольджи располагаются в местах соединения сухожилий с мышцами. Единичное нервное волокно типа Ib распадается на сложноорганизованные мельчайшие разветвления, оплетающие пучки сухожильных волокон, связанные с соединительно-тканной капсулой.
Десятки мышечных волокон погружаются в сухожильные интракапсулярные волокна, которые последовательно связываются с другими мышечными волокнами в пределах определенной мышцы. Под действием натяжения, возникающего в процессе сокращения мышцы, происходит активация луковицеобразных нервных окончаний. В связи с тем, что скорость возникновения импульсов в исходном нервном волокне зависит от силы натяжения, сухожильные нервные окончания определяют силу сокращения мышцы.
Сухожильный орган Гольджи. 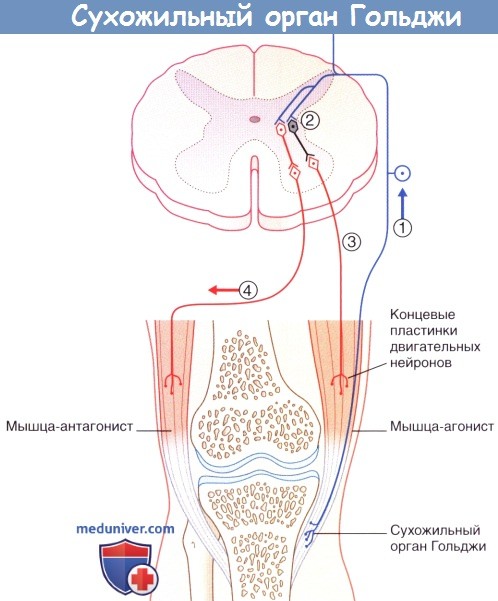
Рефлекторные эффекты при стимуляции сухожильного органа Гольджи.
(1) Сокращение мышцы-агониста вызывает возбуждение афферентов сухожильного органа Гольджи,
что приводит к (2) возбуждению тормозных вставочных нейронов, образующих синапсы с гомонимными мотонейронами,
а также (3) возбуждению тормозных вставочных нейронов, образующих синапсы с (4) мотонейронами мышцы-антагониста.
в) Свободные нервные окончания. В мышцах имеется множество свободных нервных окончаний, большинство которых расположено во внутримышечной соединительной ткани и покрывающих мышцы фасциях. Эти нервные окончания отвечают за болевую чувствительность, возникающую при непосредственном повреждающем воздействии или при накоплении продуктов распада, к которым относят молочную кислоту.
г) Иннервация суставов. Немиелинизированные нервные волокна со свободными нервными окончаниями в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Эти нервные волокна обеспечивают болевую чувствительность при деформациях суставов, а также участвуют в формировании защитного рефлекса для капсулы сустава. Так, например, переднюю капсулу запястья иннервируют срединный и локтевой нервы; внезапное растяжение при насильственном разгибании вызывает рефлекторную активацию двигательных волокон, что приводит к сгибанию верхней конечности в лучезапястном суставе.
В экспериментах на животных показано, что при воспалении сустава в возбужденном состоянии находится большее количество нервных волокон, чем при растяжении капсулы здорового сустава. Вероятно, существуют нервные окончания, единственным стимулом которых служит воспалительный процесс.
Инкапсулированные нервные окончания, расположенные внутри суставных капсул и вокруг них, представлены тельцами Руффини, реагирующими на натяжение, ламеллярными окончаниями, воспринимающими давление, а также тельцами Пачини, ответственными за ощущение вибрации.
д) Миофасциальный болевой синдром. Миофасциальный болевой синдром—распространенное заболевание, проявляющееся региональной мышечной болезненностью, связанной с чрезмерной чувствительностью пучков напряженных мышечных волокон. (Сходные синдромы наблюдают при другом заболевании—фибромиалгии. Однако при фибромиалгии происходит центральное нарушение болевой чувствительности—дисфункция системы модуляции боли.)
Прикосновение к пучкам мышечных волокон с повышенной чувствительностью вызывает боль; клинически эту область называют миофасциальной триггерной точкой. Для боли не характерно распространение в области дерматома какого-то определенного нерва; в некоторых случаях боль может выходить за пределы триггерной точки—иррадиировать. Кроме того, боль могут сопровождать вегетативные проявления, такие как покраснение и пилоэрекция. Возникновение триггерных точек может быть связано с травмой мышц, чрезмерными нагрузками во время профессиональной деятельности или спортивных занятий при нарушении процесса нормального восстановления.
Спонтанно активирующиеся очаги получили название активных миофасциальных триггерных точек (МТТ), а находящиеся в данный момент времени в неактивном состоянии—латентных миофасциальных триггерных точек. Происходящие процессы недостаточно ясны с точки зрения патофизиологии, однако установлено, что тканевая жидкость, окружающая активные МТТ, содержит большее количество ассоциированных с воспалением веществ (например, брадикинины, простагландины, протоны Н + ).
Со временем боль может захватывать новые области или усиливаться в результате раздражения нейронов задних рогов. Высвобождение другими ветвями раздраженных нейронов субстанции Р может привести к возникновению новых МТТ в области той же или прилежащей мышцы.
Стойкое сокращение мышечных волокон, прилежащих к узелковым утолщениям, объясняют инактивацией ацетилхолинэстеразы в базальной мембране их концевых двигательных пластинок. Принципы лечения этого заболевания—длительное пассивное растяжение пораженных мышц, длительное давление в горизонтальном положении больного (например, путем подкладывания теннисного мячика под пораженную область), а также механическое повреждение прокалыванием иглой или введением местных анестетиков и/или стероидов.
е) Резюме. Мышцы. Двигательная единица состоит из двигательного нейрона и иннервируемой им группы мышечных волокон. Каждая двигательная единица содержит определенный гистохимический вид мышечных волокон. Концевое утолщение нервно-мышечного синапса, содержащее пузырьки с АХ, отделено от синаптических складок сарколеммы базальной мембраной, в которой присутствует ацетилхолинэстераза.
В состав мышечных веретен входят интрафузальные мышечные волокна, активация которых происходит с обоих концов за счет γ-фузимоторных нейронов. Чувствительные волокна типа Iа образуют первичные аннулоспиральные нервные окончания в области средней части (экватора), а волокна типа II — вторичные нервные окончания. Оба типа волокон являются рецепторами растяжения. Растяжение может происходить пассивным (например, при сухожильном рефлексе) или активным путем во время фузимоторной активности. Гомонимные мотонейроны—моносинаптические; мышцы-антагонисты реципрокно ингибируются посредством вставочных нейронов Iа. Первичные афференты мышечных веретен определяют скорость мышечного сокращения, а вторичные — степень.
В процессе произвольных движений происходит одновременная активация А α- и Аγ-мотонейронов.
Сухожильные органы Гольджи определяют силу мышечных сокращений. В состав сухожильных органов входит инкапсулированное сухожилие, иннервацию которого осуществляют афференты типа Iб, вызывающие двухсинаптическое торможение гомонимных мотонейронов и реципрокное возбуждение мышц-анта-гонистов.
Свободные внутримышечные нервные окончания обеспечивают болевую чувствительность.
Суставы. Свободные нервные окончания в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Они обеспечивают болевую чувствительность и формирование суставного защитного рефлекса. Инкапсулированные нервные окончания принимают импульсы от движений суставов.
- Вернуться в оглавление раздела "Неврология."
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
В активной части опорно двигательного аппарата имеются проприорецепторы. Существует два типа проприорецепторов – нервно-мышечные веретёна, которые реагируют на изменение длины мышцы, а также сухожильные органы Гольджи, реагирующие на изменение напряжения, то есть на силу сокращения мышечных волокон (Рисунок 1).

Рисунок 1. Проприорецепторы: нервно-мышечное веретено и сухожильный орган Гольджи

Рисунок 2. Нервно-мышечное веретено
Интрафузальные волокна получают как афферентную, так и эфферентную иннервацию. Выделяют два типа афферентных окончаний – первичные и вторичные. Первичные окончания называют аннулоспиральными, они представляют собой спираль, окружающую центральную часть всех интрафузальных волокон. От первичных окончаний отходят волокна типа Ia, которые соответствуют типу Aα по Гассеру-Эрлангеру. Это самые толстые волокна (
17 мкм) с самой высокой скоростью проведения возбуждения (70 – 12 м/с). Каждое нервно-мышечное веретено иннервируется только одним волокном Ia. Другими словами одно афферентное нервное волокно Ia получает импульсацию со всех первичных окончаний в иннервируемом им веретене. Вторичные окончания имеют форму спирали, но могут также образовывать разветвлённую структуру. Они оплетают центральную часть интрафузальных волокон с ядерной цепочкой, а также статических волокон с ядерной сумкой, но их не бывает в динамических волокнах с ядерной сумкой. От вторичных окончаний отходят волокна типа II, соответствующие типу Аβ по Гассеру-Эрлангеру; диаметр этих волокон составляет
8 мкм, они проводят возбуждение со скоростью 40 – 70 м/с (Рисунок 3).

Рисунок 3. Схема нервно-мышечного веретена
Как первичные, так и вторичные нервные окончания чувствительны к изменению длины интрафузальных волокон, однако характер ответной реакции различается. При медленном растяжении центральной части интрафузальных волокон импульсация появляется как в первичных, так и во вторичных рецепторах. Частота импульсов пропорциональная степени растяжения волокна. Такой ответ афферентных окончаний называют статическим. Однако при быстром внезапном растяжении волокна в первичных (но не вторичных) окончаниях отмечается мощный всплеск импульсации – динамический ответ. Следовательно, волокна Ia передают информацию как об абсолютной длине интрафузальных волокон (то есть о степени растяжения), так о скорости её изменения, тогда как волокна II – только об абсолютной длине. Следует понимать, что растяжение центральной части интрафузальных волокон может быть связано как с удлинением всей мышцы (интрафузальные волокна расположены параллельно экстарфузальным), так и при сокращении периферических отделов волокна (Рисунок 3).

Рисунок 4. Статический и динамический ответы рецепторов нервно-мышечных веретён
Эфферентная иннервация интрафузальных волокон представлена нервными волокнами типа Aγ, которые являются аксонами γ-мотонейронов спинного мозга. Диаметр этих волокон составляет
5 мкм, скорость проведения возбуждения в них – 15 – 40 м/с. Окончания Aγ волокон подходят к периферическим отделам интрафузальных волокон, при возбуждении γ-мотонейронов концевые участки интрафузальных волокон сокращаются, в результате чего их центральные отделы растягиваются. Выделяют γ-динамические и γстатические волокна, γ-динамические волокна иннервируют динамичекие волокна с ядерной сумкой, их активация повышает динамическую чувствительность окончаний Iа типа.
γ-статические волокна иннервируют волокна с ядерной цепочкой и статические волокна с ядерной сумкой, их возбуждение повышает уровень активности как первичных, так и вторичных окончаний (см. ниже).
Миотатический рефлекс (рефлекс растяжения)
В ответ на увеличение длины мышцы рецепторы нервно-мышечных веретён возбуждаются, и импульсация по волокнам Ia (и при быстром, и при медленном растяжении) или II (при медленном растяжении) поступает в спинной мозг. Войдя в серое вещество спинного мозга, волокно афферентного нейрона разветвляется и образует возбуждающие синапсы практически со всеми α-моторными нейронами, иннервирующими гомонимную (то есть эту же) мышцу. Возбуждение α-моторных нейронов вызывает сокращение экстрафузальных волокон, и мышца укорачивается. Следует отметить, что коллатерали афферентных нейронов переключаются также на вставочные тормозные нейроны, которые угнетают α-мотонейроны мышц антагонистов. Таким образом, растяжение мышцы вызывает её рефлекторное укорочение, при этом мышцы антагонисты расслабляются. Описанный рефлекс с рецепторов нервно-мышечных веретён называется миотатическим, или рефлексом на растяжение (Рисунок 4). Миотатический рефлекс является моносинаптическим, поэтому имеет очень небольшой латентный период (

Рисунок 5. Миотатический рефлекс с нервно-мышечных веретён
Функции γ-моторных нейронов. Понятие о γ-петле.
Как уже упоминалось, интрафузальные волокна получают эфферентную иннервацию со стороны γ-моторных нейронов. При возбуждении γ-мотонейронов происходит сокращение периферических (концевых) отделов интрафузальных волокон, в результате их центральная часть растягивается. При этом первичные и вторичные нервные окончания активируются, импульсация от них поступает в спинной мозг, и возбуждаются α-моторные нейроны. Другими словами, запускается миотатический рефлекс. Такой механизм (активация рефлекса растяжения при возбуждении γ-мотонейронов) называется γ-петлёй. Наличие γ-петли обеспечивает сохранение чувствительности рецепторов нервномышечных веретён при разной длине мышцы. Во время сокращения мышцы её длина уменьшается, соответственно должны укорачиваться и интрафузальные волокна. В результате импульсация с рецепторов нервно-мышечных веретён должна была бы прекратиться, и рецепторы потеряли бы способность реагировать на изменение длины мышцы. Напротив, при расслаблении мышцы её длина увеличивается, и интрафузальные волокна могут быть чрезмерно растянуты. В таких условиях рецепторы будут генерировать импульсы с максимальной частотой и, следовательно, также потеряют способность реагировать на изменение длины. Однако в реальных условиях имеет место α-γ-коактивация, то есть α- и γмотонейроны возбуждаются одновременно. В результате сокращаются как экстра-, так и интрафузальные волокна. Сокращение экстрафузальных волокон приводит к укорочению мышцы, а сокращение периферических отделов интрафузальных волокон – к растяжению их центральной части. Следовательно, длина центральной части не меняется, поскольку её укорочение вследствие сокращения экстрафузальных волокон компенсируется растяжением из-за сокращения периферических отделов интрафузальных волокон. И наоборот – расслабление всей мышцы сопровождается расслаблением и интрафузальных волокон, при этом растяжение центральной части уменьшается. Таким образом, за счёт α-γ-коактивации рецепторы нервно-мышечных веретён сохраняют чувствительность (и, следовательно, передают информацию о длине мышцы в ЦНС) как при сокращении мышцы, так и при её расслаблении (Рисунок 6). На γ-мотонейронах заканчиваются нисходящие пути из различных отделов головного мозга. Меняя активность γ-мотонейронов, мозговые центры влияют на чувствительность рецепторов нервно-мышечных веретён и, следовательно, на порог рефлекса растяжения. Сухожильные органы Гольджи. Обратный миотатический рефлекс Кроме нервно-мышечных веретён в активной части опорно-двигательного аппарата имеется ещё один тип проприрецепторов – сухожильные органы Гольджи. Сухожильные органы представляют собой сеть коллагеновых нитей, окружённую тонкой соединительнотканной капсулой. Диаметр сухожильного органа Гольджи составляет 100 мкм, длина – 1 мм. Коллагеновые нити внутри органа оплетённых нервными окончаниями, от которых отходят нервные волокна типа Ib. эти нервные волокна, как и Ia, соответствуют типу Aα по Гассеру-Эрлангеру. Сухожильные органы Гольджи расположены в месте прикрепления мышечных волокон к сухожильным, при этом каждый сухожильный орган связан с 3 – 25 (в среднем – с 10 – 15) мышечными волокнами (Рисунок 7). Таким образом, в отличии от нервно-мышечных веретён, которые ориентированы параллельно мышечным
волокнам, сухожильные органы по отношению к мышечным волокнам расположены последовательно.

Рисунок 6. Функция γ-петли
Следовательно, сухожильные органы реагируют не на изменение длины мышцы, а на развиваемое ей усилие, другими словами, на степень её напряжения. Афферентные нервные волокна от сухожильных органов Гольджи следуют в спинной мозг, где, в отличие от чувствительных волокон с нервно-мышечных веретён, переключаются на вставочные тормозные нейроны. Вставочные нейроны, в свою очередь, образуют синаптические контакты с α-мотонейронами гомонимной мышцы. Таким образом, активация сухожильных органов приводит к расслаблению мышцы, к которой крепится данное сухожилие. Описанный рефлекс называется обратным рефлексом растяжения (обратным миотатическим рефлексом) (Рисунок 8). Этот рефлекс, по-видимому, имеет две функции. Во-первых, он предупреждает чрезмерное напряжение скелетных мышц, которое может привести к тяжёлым травмам (разрыву мышцы, отрыву сухожилия от кости или от мышцы). Во-вторых, обратный миотатический рефлекс обеспечивает равномерность силы сокращения разных мышечных волокон в пределах одной мышцы. Те волокна, которые напряжены слишком сильно, расслабляются за счёт усиления тормозных влияний со стороны спинного мозга. Напряжение волокон, которые сокращаются недостаточно сильно, напротив, усиливается за счёт уменьшения тормозного воздействия.

Рисунок 7. Сухожильный орган Гольджи
Проприорецепторы посылают в головной мозг информацию о состоянии скелетно-мышечной системы Импульсация с проприорецепторов поступает не только в спинной мозг, но и по коллатералям афферентных волокон в различные отделы головного мозга. В частности, импульсы с нервно-мышечных веретён и сухожильных органов следуют по спинномозжечковым трактам в мозжечок, а также по системе задних столбов (тонкий и клиновидный пучок) в продолговатый мозг, затем – в таламус, а оттуда – в кору больших полушарий. Таким образом, мозжечок и кора большого мозга постоянно получают информацию о состоянии скелетно-мышечной системы, а именно о длине мышц и о степени их напряжения. Эта информация является необходимой как для регуляции произвольных движений и позы, так и для осознания положения частей тела в пространстве и друг относительно друга.

Рисунок 8. Обратный миотатический рефлекс с сухожильных органов Гольджи
Нервно-сухожильные веретена представляют собой сенсорные рецепторы, через которые проходит небольшой пучок мышечно-сухожильных волокон. Они расположены проксимально ее места прикрепления сухожильных волокон к мышечным (рис. 33). Около 5 — 25 мышечных волокон, как правило, соединены с каждым нервно-сухожильным веретеном. Если нервно-мышечные веретена следят за длиной мышцы, то эти структуры чувствительны к напряжению в мышечно-сухожильном комплексе и действуют подобно тензиометру. Их чувствительность настолько высока, что они могут реагировать на сокращение отдельного мышечного волокна. По сути, они являются тормозными и выполняют защитную функцию, снижая вероятность травмы. При стимулировании они тормозят сократительные (агонисты) мышцы и возбуждают антагонистические.
По мнению некоторых ученых, уменьшение влияния нервно-сухожильных веретен приводит к растормаживанию активных мышц, обеспечивая более мощное их сокращение. Этот механизм объясняет, по крайней мере частично, прирост мышечной силы вследствие тренировок силовой направленности.
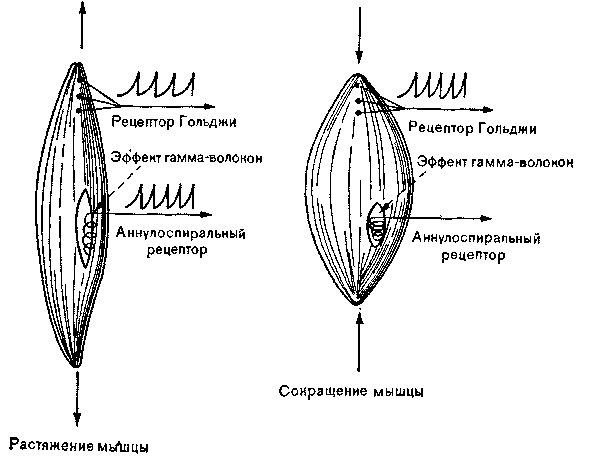
Рис. 34. Схема активации проприоцепторов.
Нервно-сухожильные и нервно-мышечные веретена представляют собой сенсорные рецепторы – классические представители проприоцепторов (рис. 34). Аннулоспиральный рецептор - нервно-мышечное веретено - разряжается только при растяжении мышцы, а рецептор Гольджи - нервно-сухожильное веретено - с более высоким порогом, отвечает и на растяжение, и на сокращение Механизм гамма-эфферентов контролирует только аннулоспиральные рецепторы.
ВЫСШИЕ ЦЕНТРЫ ГОЛОВНОГО МОЗГА
| Рефлексы представляют собой простейшую форму нервной интеграции. Однако большинство движений включает контроль и координацию со стороны высших центров головного мозга, а именно: двигательной области коры головного мозга; базальных ядер; мозжечка. |
Высший уровень сенсорно-двигательной (сенсомоторной) интеграции:
• Двигательная область коры головного мозга
• Базальные ядра
Выясним некоторые функции каждого из них.
Двигательная область коры головного мозга осуществляет контроль тонких и разрозненных мышечных движений. Она находится в лобной доле в передней центральной извилине. Находящиеся здесь нейроны, так называемые пирамидальные, позволяют нам сознательно контролировать движения скелетных мышц. Двигательная область коры головного мозга представляет собой часть головного мозга, которая решает, какое движение вы хотите выполнить. Участки, требующие тончайшего двигательного контроля, в большей степени представлены на ней, тем самым обеспечивается больший нервный контроль.
Тела пирамидных нейронов находятся в двигательной области коры головного мозга, и их аксоны образуют экстрапирамидные пути. Это так называемые корково-спинномозговые пути, поскольку нервные процессы проходят от коры головного мозга до спинного мозга без переключения. Эти пути обеспечивают главный произвольный контроль скелетных мышц.
Базальные ядра. Базальные ядра не являются частью коры головного мозга. Они находятся в белом веществе, глубоко под корой головного мозга. Базальные ядра представляют собой скопления нейронов. Сложные функции их изучены недостаточно; известна их важная роль в инициации движений продолжительного или повторяющегося характера (таких, как движения рук при ходьбе), следовательно, они контролируют сложные полупроизвольные движения, например, ходьбу и бег. Кроме того, они участвуют в поддержании мышечного тонуса и позы.
Мозжечок. Мозжечок играет важнейшую роль в контроле всех быстрых и сложных видов мышечной деятельности. Он помогает синхронизировать двигательную деятельность и быстрый переход от одного движения к другому, следя и внося необходимые изменения в двигательную деятельность, вызванную другими участками головного мозга. Мозжечок содействует функционированию как двигательной области коры головного мозга, таки базальных ядер. Он “смягчает” структуру движений, в противном случае они бы были резкими и не координируемыми.
Мозжечок выполняет роль интеграционной системы, сравнивая запрограммированную деятельность с изменениями, которые происходят в организме, и производя затем соответствующие корригирующие действия с помощью двигательного отдела. Он получает информацию из головного мозга, а также от проприорецепторов, находящихся в мышцах и суставах, которые сообщают о положении, занимаемом в данный момент телом. Кроме того, мозжечок получает зрительные импульсы, а также импульсы о равновесии. Следовательно, он обрабатывает всю поступающую информацию о напряжении и положении всех мышц, суставов и сухожилий, а также положении тела относительно окружающих условий и затем определяет наилучший план действий, направленный на выполнение необходимого движения. Таким образом, в мозжечке формируется и при необходимости корректируется программа выполнения произвольного движения.
Дата добавления: 2016-12-16 ; просмотров: 2698 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Читайте также: