
Я диски в мышечных тканях
Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости.) Важнейшие функции мышечной ткани: сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечно-полосатая (скелетная) и сердечная мышечные ткани.

Эта мышечная ткань встречается в стенках внутренних органах (кишечник, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов - коротких одноядерных клеток. Слабо выражено межклеточное вещество, клетки сближены друг с другом: благодаря этому возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру, мочевого пузыря), практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов - миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их изучим.)
Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой: человек не может управлять ей произвольно. К примеру, невозможно по желанию сузить или расширить зрачок.

Скелетная ткань образует мышцы туловища, конечностей и головы.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер - миосимпластами. Миосимпласт представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметром.
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой.

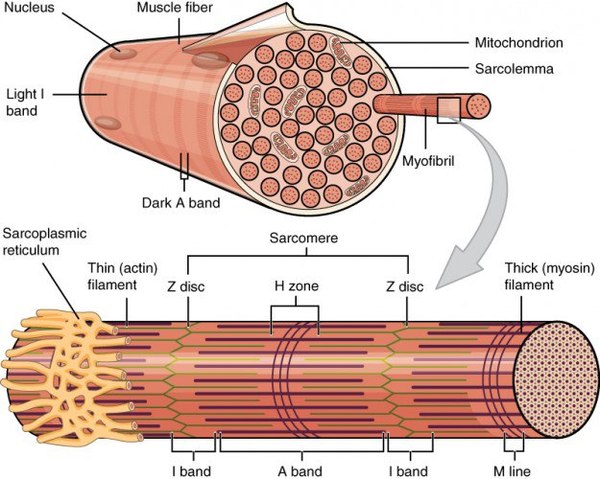
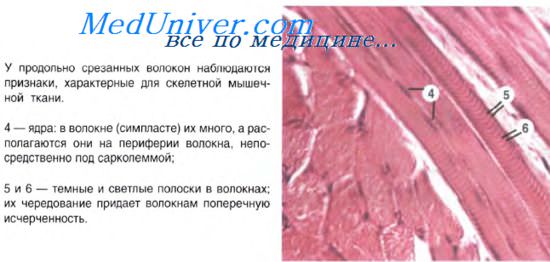
Характерная черта данной ткани - поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы - саркомер.
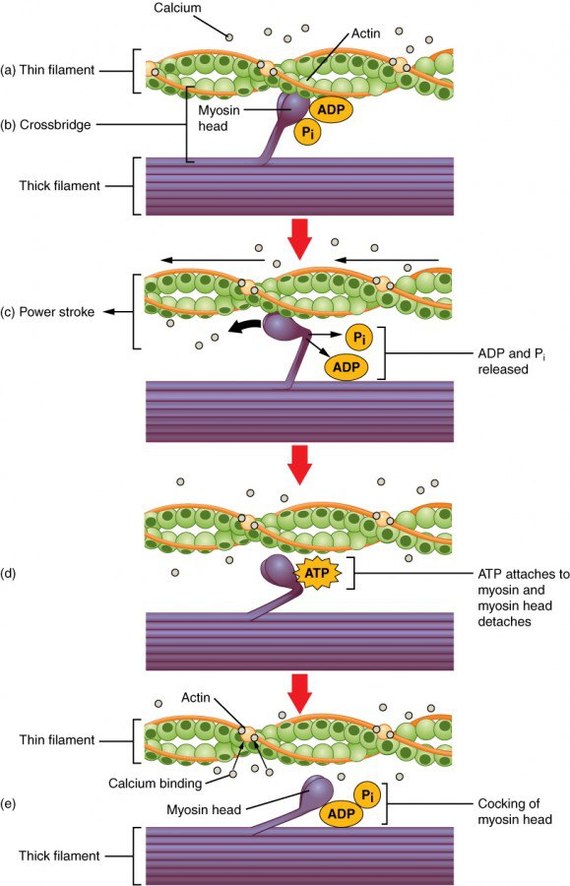
Сократимость мышечной ткани обусловлена наличием в клетках миофиламентов. Саркомер - элементарная сократительная единица мышцы. Состоит из тонкого белка - актина, и толстого - миозина. Сокращение осуществляется благодаря трению нитей актина о нити миозина, в результате чего саркомер укорачивается.

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином (белком между нитями актина), что обуславливает соединение актина и миозина. При сокращении мышц выделяется тепло.
Замечу, что трупное окоченение - посмертное затвердевание мышц - связано именно с ионами кальция, которые устремляются в область низкой концентрации (мышцы), способствуя связыванию актина и миозина. Мертвый организм не способен разорвать цикл, возникший в мышцах, в связи с чем наблюдается стойкая мышечная контрактура: конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние волокна не возбуждают друг друга, в отличие от гладких миоцитов. Скелетные мышцы быстро утомляются и сокращаются мгновенно (у гладких мышц фазы сокращения и расслабления растянуты во времени.)
Скелетные мышцы поддаются нашему осознанному контролю, их скоращение регулируется произвольно. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.


Этот тип мышечной ткани удивительным образом сочетает характеристики двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство. Сердечная мышечная ткань состоит из одиночных клеток, имеющих поперечно-полосатую исчерченность.
В некоторых участках эти клетки смыкаются, образуя между собой контакты, благодаря которым возбуждение одной клетки волнообразно передается на соседние, таким образом, охватываются новые участки миокарда. Сокращается эта ткань непроизвольно, не утомляется.
Сердечная ткань обладает уникальным свойством - автоматизмом - способностью возбуждаться и сокращаться без влияний извне, самопроизвольно. Это легко можно подтвердить, изолировав сердце лягушки из организма в физиологический раствор: сокращения сердца в нем будут продолжаться еще несколько часов.

Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.


Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца - состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Мышцы развиваются из среднего зародышевого листка - мезодермы.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
11.1. Введение
б) Благодаря данной способности, мышечные ткани обеспечивают
11.1.1.1. Виды мышечных тканей и их происхождение
| Виды мышечных тканей | П р о и с х о ж д е н и е | |
| I. Поперечно- полосатые (исчерченные) мышечные ткани | 1. Скелетная мышечная ткань | Из миотомов |
| 2. Сердечная мышечная ткань | Из миоэпикардиальной пластинки (находящейся в составе висцерального листка спланхнотома). | |
| II. Гладкие (неисчерченные) мышечные ткани | 1. Гладкая мышечная ткань сосудов и внутренних органов | Из мезенхимы |
| 2. Мышечная ткань нейрального происхождения (мышцы радужки глаза) | Из клеток нейрального зачатка в составе стенки глазного бокала. | |





| Тип мышечной ткани | |||
| Поперечно-полосатая скелетная | Поперечно-полосатая сердечная | Гладкая | |
| Источник развития | Миотомы мезодермы | Миоэпикардиальная пластинка (висцеральный листок спланхнотома) | Мезенхима |
| Структурно-функциональная единица | Мышечное волокно - миосимпласт | Клетка - кардиомиоцит | Клетка - гладкий миоцит |
| Расположение ядер | многочисленные, по периферии | В центре (одно или два) | В центре (одно) |
| Исчерченность | Актин и миозин формируют характерные полоски | Актин и миозин формируют характерные полоски | Характерных полосок нет |
| Сократительный аппарат | Миофибриллы | Миофибриллы | Миофиламенты |
| Т-система | Т-трубочки на уровне A-I дисков; триады | Т-трубочки на уровне Z-линии, диады | Нет Т-трубочек, нет триад и диад, есть кавеолы |
| Z-линия | есть | есть | Нет, имеются плотные тельца |
| Кальций-связывающий белок | тропонин | тропонин | кальмодулин |
| Межклеточные соединения | нет | Вставочные диски – комплекс межклеточных контактов | Щелевые контакты (нексусы) |
| Источник регенерации | Деление миосателлитоцитов, гипертрофия | внутриклеточная гипертрофия | Деление миоцитов, гипертрофия |
| Источник иннервации | Соматический отдел нервной системы | Вегетативный отдел нервной системы | Вегетативный отдел нервной системы |
| Характер сокращения | Тетанический произвольный | Ритмический непроизвольный | Тонический непроизвольный |
| Несократительные функции | Участие в терморегуляции и углеводном обмене | Синтез гормона – натрийуретического пептида (секреторные кардиомиоциты правого предсердия) | Синтез эластических волокон, коллагена III типа |
СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ
РАЗВИТИЕ
Источником развития скелетной мышечной ткани являются миотомы сомитов. Стадии развития:
- Миобластическая стадия. Клетки миотомов превращаются в миобласты и мигрируют к местам закладки мышц. Миобласты делятся митозом. Часть миобластов обособляется в виде миосателлитоцитов, которые сохраняют свойства малодифференцированных клеток до конца жизни.
- Миосимпластическая стадия. Миобласты располагаются в виде цепочек и сливаются друг с другом. Образуются миосимпласты. В их цитоплазме образуются миофибриллы из сократительных белков, которые начинают синтезироваться ещё в миобластах. Миофибриллы лежат на периферии миосимласта, ядра занимают центральное положение.
- Стадия миотубул. В симпластах увеличивается число миофибрилл. Длина их увеличивается.
- Стадия зрелого мышечного волокна. В эту стадию объём миофибрилл увеличивается до такой степени, что они занимают основную массу волокна, смещаясь в центр и сдвигая ядра на периферию.
СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА.
Мышечные волокна являются структурно-функциональным элементом скелетной мышечной ткани. Они имеют длину до 20-30 см, толщину около 100 мкм. Состоят из двух частей:
Мышечное волокно снаружи покрыто сарколеммой. Сарколемма состоит из толстой базальной мембраны и плазмолеммы мышечного волокна. Между базальной мембраной и плазмолеммой в отдельных участках имеются углубления, в которых расположены миосателлитоциты. Миосателлитоциты – камбиальные клетки скелетной мышечной ткани. Миосателлитоцит – типичная одноядерная клетка, окруженная своей плазмолеммой, имеет слабо развитые органеллы.
Миосимпласт содержит множество (до несколько тысяч) ядер, лежащих на периферии волокна, под сарколеммой.
Протоплазму волокна называют саркоплазмой. В ней находятся органеллы общего значения (за исключением центриолей), органеллы специального значения – миофибриллы, и включения.

СТРОЕНИЕ МИОФИБРИЛЛ.
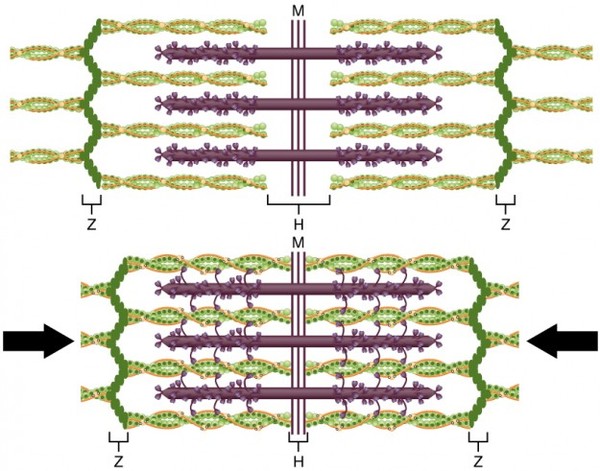
Миофибриллы, числом до двух тысяч в одном волокне, занимают основную часть волокна. Их длина равна длине волокна, диаметр до 2 мкм. В каждой миофибрилле при световой микроскопии обнаруживается исчерченность – чередование светлых и тёмных дисков. В поляризованном свете тёмные диски имеют двойное лучепреломление и поэтому тёмные диски называются анизотропными, или А-дисками. Светлые диски не имеют двойного лучепреломления и называются изотропными, или I-дисками.
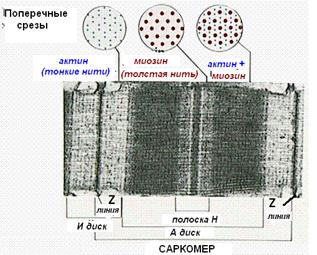
Посередине I-диска проходит тёмная полоска, которая называется Z-линией,или телофрагмой.На поперечном разрезе телофрагма представляет собой решётку, в узлах которой закрепляются актиновые филаменты.
В центре А-диска находится более светлая полоска Н, а посередине её проходит тёмная линия М, или мезофрагма.
Участок миофибриллы, лежащий между соседними Z-линиями, называется саркомером. Саркомер – структурно-функциональная единица миофибриллы.В состав саркомера последовательно входят:
· Z-линия;
· ½ диска I;
· диск А;
· ½ диска I
· вторая Z-линия.
Каждый саркомер состоит из тонких актиновых и толстых миозиновых филаментов.
В составе тонких (диаметр 5 нм) актиновых филаментов входят белки:
· актин;
· тропонин;
· тропомиозин.
Молекулы актина имеют глобулярное строение – G-актин. Эти молекулы соединяются вместе в длинные цепочки – фибриллярный, F-актин. В актиновых филаментах две цепи F-актина образуют двойную спираль. В бороздках между цепями спирали лежат молекулы тропомиозина. К молекулам тропомиозина на равных расстояниях друг от друга прикрепляются молекулы тропонина. Молекула тропонина состоит из трёх субъединиц: TnT, TnI, TnC. TnTосуществляет прикреплениетропонина к тропомиозину.TnCотвечает за связывание с ионами кальция. TnIпрепятствует взаимодействию миозина с актином.

Толстые филаменты (диаметр 12 нм) содержат белок миозин. Каждая молекула миозина состоит из двух частей: головки и хвоста и может сгибаться в двух местах – шарнирных участках. Головка миозина имеют АТФ-азную активность и способна расщеплять АТФ с образованием энергии. Молекулы миозина соединяются в пучки и образуют толстые миозиновые филаменты. По периферии толстых филаментов находятся участки, содержащие головки миозина. Центральная часть не содержит головок.
В составе саркомера толстые филаменты лежат только в диске А. Тонкие филаменты расположены в дискеI, но концами частично заходят в диск А между миозиновыми филаментами. Та часть диска А, которая содержит и актиновые и миозиновые филаменты, выглядит на срезах более тёмной, а та его часть, которая содержит только миозиновые филаменты, светлее. Эта часть диска А, которая содержит только миозиновые филаменты, и составляет полоску Н. Таким образом:
· диск I состоит из актиновых филаментов;
· полоска Н диска А состоит из миозиновых филаментов;
· на периферии диска А есть зона пересечения актиновых и миозиновых филаментов.
На поперечном срезе миофибриллы можно видеть, что в зоне пересечения вокруг одной толстой филаменты лежат шесть тонких филамент.
Тонкие филаменты неподвижно прикреплены к Z-линиям. В состав Z-линий входят белки α-актинин, десмин, виментин.
Линия М в центре Н-полоски – место соединения всех миозиновых филаментов друг с другом. В их скреплении участвуют белки миомезин и С-белок.
Читайте также:
- Болят все суставы что делать чем лечить и отзывы
- Поражения почек при системных васкулитах
- Фиброма на голосовых связках как лечить
- Информатика как наука шпора
- Дэнас аппарат для лечения суставов отзывы