Динамическая нестабильность микротрубочек
Добавил пользователь Владимир З. Обновлено: 27.01.2026
Микротрубочки — белковые внутриклеточные структуры, входящие в состав цитоскелета.
Микротрубочки представляют собой полые внутри цилиндры диаметром 25 нм. Длина их может быть от нескольких микрометров до, вероятно, нескольких миллиметров в аксонах нервных клеток. Их стенка образована димерами тубулина. Микротрубочки, подобно актиновым микрофиламентам, полярны: на одном конце происходит самосборка микротрубочки, на другом — разборка. В клетках микротрубочки играют роль структурных компонентов и участвуют во многих клеточных процессах, включая митоз, цитокинез и везикулярный транспорт.
Содержание
Строение
Микротрубочки — это структуры, в которых 13 протофиламентов, состоящих из гетеродимеров α- и β-тубулина, уложены по окружности полого цилиндра. Внешний диаметр цилиндра около 25 нм, внутренний — около 15.
Один из концов микротрубочки, называемый плюс-концом, постоянно присоединяет к себе свободный тубулин. От противоположного конца — минус-конца — тубулиновые единицы отщепляются.
В образовании микротрубочки выделяют три фазы:
- замедленная фаза, или нуклеация. Это этап зарождения микротрубочки, когда молекулы тубулина начинают соединяться в более крупные образования. Такое соединение происходит медленнее, чем присоединение тубулина к уже собранной микротрубочке, поэтому фаза и называется замедленной;
- фаза полимеризации, или элонгация. Если концентрация свободного тубулина высока, его полимеризация происходит быстрее, чем деполимеризация на минус-конце, за счет чего микротрубочка удлиняется. По мере её роста концентрация тубулина падает до критической и скорость роста замедляется вплоть до вступления в следующую фазу;
- фаза стабильного состояния. Деполимеризация уравновешивает полимеризацию, и рост микротрубочки останавливается.
Лабораторные исследования показывают, что сборка микротрубочек из тубулинов происходит только в присутствии гуанозинтрифосфата и ионов магния.
Динамическая нестабильность
Микротрубочки являются динамическими структурами и в клетке постоянно полимеризуются и деполимеризуются. Центросома, локализованная вблизи ядра, выступает в клетках животных и многих протистов как центр организации микротрубочек (ЦОМТ): они растут от неё к периферии клетки. В то же время микротрубочки могут внезапно прекратить свой рост и укоротиться обратно по направлению к центросоме вплоть до полного разрушения, а затем вырасти снова. При присоединении к микротрубочке молекулы тубулина, несущие ГТФ, образуют «шапочку», которая обеспечивает рост микротрубочки. Если локальная концентрация тубулина падает, связанная с бета-тубулином ГТФ постепенно гидролизуется. Если полностью гидролизуется ГТФ «шапочки» на ±конце, это приводит к быстрому распаду микротрубочки. Таким образом, сборка и разборка микротрубочек связана с затратами энергии ГТФ.
Динамическая нестабильность микротрубочек играет важную физиологическую роль. Например, при делении клетки микротрубочки растут очень быстро и способствуют правильной ориентации хромосом и образованию митотического веретена.
Функция
Микротрубочки в клетке используются в качестве «рельсов» для транспортировки частиц. По их поверхности могут перемещаться мембранные пузырьки и митохондрии. Транспортировку по микротрубочкам осуществляют белки, называемые моторными. Это высокомолекулярные соединения, состоящие из двух тяжёлых (массой около 300 кДа) и нескольких лёгких цепей. В тяжёлых цепях выделяют головной и хвостовой домены. Два головных домена связываются с микротрубочками и являются собственно двигателями, а хвостовые — связываются с органеллами и другими внутриклеточными образованиями, подлежащими транспортировке.
Выделяют два вида моторных белков:
Динеины перемещают груз только от плюс-конца к минус-концу микротрубочки, то есть из периферийных областей клетки к центросоме. Кинезины, напротив, перемещаются к плюс-концу, то есть к клеточной периферии.
Перемещение осуществляется за счёт энергии АТФ. Головные домены моторных белков для этого содержат АТФ-связывающие участки.
Помимо транспортной функции, микротрубочки формируют центральную структуру ресничек и жгутиков — аксонему. Типичная аксонема содержит 9 пар объединённых микротрубочек по периферии и две полных микротрубочки в центре. Из микротрубочек состоят также центриоли и веретено деления, обеспечивающее расхождение хромосом к полюсам клетки при митозе и мейозе. Микротрубочки участвуют в поддержании формы клетки и расположения органоидов (в частности, аппарата Гольджи) в цитоплазме клеток.
Растительные микротрубочки
Микротрубочки растений являются высокодинамическими составляющими цитоскелета, которые вовлечены в важные клеточные процессы, в частности, сегрегацию хромосом, формирование фрагмопласта , микрокомпартментализацию, внутриклеточный транспорт, а также в поддержание постоянной формы и полярности клетки. Мобильность микротрубочек обеспечивается динамической нестабильностью, передвижением полимеров моторными белками, тредмилингом (en:Treadmilling) и гибридным механизмом тредмилинга с динамической нестабильностью плюс-конца и медленной деполимеризацией минус-конца [1] .
Организация и динамика
Микротрубочки чрезмерно чувствительны к биотическим и абиотическим факторам окружающей среды (холоду, освещению, засухе, засолению, влиянию гербицидов и пестицидов, затоплению, сжатию, воздействию электрического поля, давлению и силе тяжести), а также к фитогормонам, антимитотическим препаратам и ряду других биологически активных соединений [2] . Микротрубочки являются полыми полярными цилиндрическими филаментами диаметром свыше 24 нм, которые собираются из гетеродимеров α-и β-тубулина, которые в положении «голова-к-хвосту» формируют 13 протофиламентов.
Существенное ограничение иммуногистохимических исследований состоит в невозможности прижизненной визуализации динамики микротрубочек эукариотических и прокариотических клеток в режиме реального времени. Это ограничение было преодолено благодаря применению конфокальной микроскопии с зеленым флуоресцентным белком, изолированным из медузы Aequorea victoria L. [3] . Репортёрная конструкция GFP-MBD для гетерологической трансформации даже при низком уровне транзиентной экспрессии in vivo и in vitro позволяет визуализировать динамическую нестабильнось микротрубочек в разных типах клеток корня [4] [5] .
В клетках высших растений присутствуют четыре типа построений микротрубочек:
- сетка кортикальных и эндоплазматических микротрубочек,
- препрофазная лента,
- митотическое веретено,
- фрагмопласт [6] .
Белки, ассоциированные с микротрубочками
Все компоненты цитоскелета и другие органеллы связаны между собой рядом специфических белков, ассоциированных с микротрубочками (БАМ). В животных клетках наиболее исследованными БАМ является tau и БАМ2, которые стабилизируют микротрубочки и присоединяют их к другим клеточным структурам, а также транспортные белки динеин и кинезин [7] . Функционирование различных групп растительных микротрубочек зависит от наличия изоформ БАМ из семьи БАМ65 и регуляторных киназ и фосфатаз. В частности, высококонсервативный животный гомолог семьи БАМ65 важен для получения микротрубочками определенных конфигураций на протяжении развития растения [5] . Ориентация и организация различных популяций и типов построений микротрубочек является ткане- и органоспецифической [8] .
Построение корня Резуховидки Таля Arabidopsis thaliana L. типично для двудольных растений. Ближайшим к поверхности корня является эпидермальный слой, клетки которого в зрелой зоне в зависимости от способности инициировать развитие корневых волосков являются трихобластами или атрихобластами [9] . Глубже расположены накопительный безхлоропластный кортикальный слой с многочисленными межклетниками и плазмодесмами и слой эндодермальных клеток с поясками Каспари на антиклинальных поверхностях [10] . Центральный цилиндр корня формируют паренхимные клетки перицикла [10] , которые способны к быстрому делению, и элементы ксилемы и флоэмы. Присутствует и функциональное разграничение корневых зон: зоны деления, элонгации, созревания, а также переходная зона на границе зон инициации и элонгации. С перициклом формируются боковые корни, а с трихобластами эпидермального слоя — корневые волоски [11] [10] . Кончик корня покрыт корневым чехликом со специфической морфологией клеток колумеллы.
Кортикальные микротрубочки
Ацентросомальные кортикальные микротрубочки (КМТ) важны для морфогенеза растений, регуляции клеточного деления и элонгации [12] . Высокодинамическая популяция мембраносвязанных коротких КМТ быстро реориентуеться из интерфазного поперечного положения в продольное при элонгации клетки [13] . Ацентросомальные кортикальные микротрубочки имеют неупорядоченное размещение плюс-концов и обнаруживают динамическую нестабильность, а свободные минус-концы КМТ медленно деполимеризируются, то есть КМТ самоорганизуются гибридным механизмом динамической нестабильности и тредмилинга [1] . Энуклеация происходит по всей поверхности плазматической мембраны [13] [1] . Белок SPR1 регулирует динамику и организацию плюс-конца КМТ растений, что сказывается на анизотропном росте клетки [14] [15] . Ацентросомальные кортикальные микротрубочки располагаются параллельно целлюлозным микрофибриллам [16] , правильная организация КМТ является существенной для нормального синтеза клеточной стенки [17] . Установлено, что КМТ объединяются в узлы, которые часто пересекаются для стабилизации микротрубочек и удержания белков на их поверхности [15] .
Латеральные цилиндрические выросты трихобластов, корневые волоски, достигают значительной длины относительно собственной толщины с достаточно постоянным диаметром у Arabidopsis thaliana L. (незрелые ~ 6-10 нм; зрелые — более 1 мм) и характеризуются высокополярной цитоархитектурой [18] . Удлинение их происходит посредством верхушечного роста (англ. tip growth ) путем поляризованного экзоцитоза, который отмечается возвратно-фонтанным током цитоплазмы, градиентом цитоплазматического Ca 2+ , активностью F-актина и смещением клеточного содержимого к верхушке волоска. На ранних стадиях развития корневые волоски 3-дневных проростков Arabidopsis thaliana L. растут со скоростью 0,4 мкм / мин, ускоряясь позже до 1-2,5 мкм / мин [18] .
Растительным клеткам присуща организованная популяция кортикальных микротрубочек [7] , которая в корневых волосках присутствует на всех уровнях развития [19] [20] . При переходе из зачаточного состояния в состояние удлинения, кортикальные микротрубочки верхушки волосков не визуализируются, поскольку появляются эндоплазматические микротрубочки. Кортикальные микротрубочки ориентированы продольно или спирально [20] [21] . У кукурузы Zea mays L. и салата Lactuca sativa L. инициация роста корневых волосков связана с реорганизацией популяции КМТ в трихобластах [22] [23] [20] . Эта популяция контролирует стабильность и направление апикального роста корневых волосков [24] [25] . Сравнение четырех стандартных параметров динамической нестабильности КМТ in vivo — уровня ростовой активности, скорости разборки, частоты переходов от разборки к росту («спасение») и наоборот («катастрофа») выявило, что кортикальные микротрубочки (КМТ) молодых корневых волосков являются динамичными, потому что зрелые. Сетка микротрубочек реорганизуется в ответ на меняющиеся параметры окружающей среды и стимулы дифференциации путем варьирования показателей динамической нестабильности [25] .
Динамическая нестабильность микротрубочек
Динамическая нестабильность микротрубочек
• Микротрубочки постоянно переключаются с фазы роста на фазу укорачивания; этот процесс обозначается как динамическая нестабильность
• Переход от состояния роста к состоянию укорачивания называется катастрофой
• Переход от состояния укорачивания к состоянию роста называется спасением
• Популяция микротрубочек растет и укорачивается асинхронно; в любой момент времени большая их часть находится в стадии роста и меньшая часть — в стадии укорачивания
• Строение растущих и укорачивающихся концов микротрубочек различно: растущие концы имеют выступы протофиламентов, а укорачивающиеся характеризуются участками, отклоняющимися в сторону от оси микротрубочки
В экспериментах in vitro, предпринятых с целью выяснения динамики и кинетики полимеризации микротрубочек, вначале использовали такие методы, как светорассеяние. Этим методом можно было измерять лишь общее количество полимера, присутствующее в данный момент времени. В середине 1980-х гг. были разработаны методы иммунофлуоресценции и электронной микроскопии, которые позволяли измерять степень полимеризации тубулина. В отличие от светорассеяния, эти методы позволяли визуализировать отдельные микротрубочки. В дальнейшем, эти микротрубочки можно было сосчитать и измерить их длину. Результаты этих экспериментов оказались совершенно неожиданными и помогли ответить на вопрос, почему обмен субъединиц тубулина в полимере происходит с большей скоростью, чем это можно предполагать на основании представления об установлении равновесия.
Если бы микротрубочки относились к равновесным полимерам, то в состоянии равновесия они бы обнаруживали лишь незначительные изменения длины, и их общее количество со временем существенно бы не менялось. Однако, используя новые методы, удалось показать, что в состоянии равновесия изменяются как длина, так и количество микротрубочек. В продолжении примерно 40 мин некоторые микротрубочки становятся в десять раз длиннее, в то время как общее их количество снижается. В дополнительных экспериментах по исследованию микротрубочек, образующихся на центросоме, также получились неожиданные результаты. Вначале микротрубочки полимеризовали на центросоме, а затем пробу разбавляли до концентрации тубулина ниже критической. В случае равновесного полимера все микротрубочки должны были бы сразу же начать разбираться, и этот процесс продолжался бы до повторного установления критической концентрации.
Однако, хотя некоторые микротрубочки, действительно, диссоциировали, остальные продолжали полимеризацию, т. е. полученный результат оказался несовместимым с представлением о равновесном полимере. Эти, а также другие результаты позволили предложить модель полимеризации микротрубочек в ходе процесса, который получил название динамическая нестабильность. При этом процессе микротрубочки постоянно находятся либо в фазе роста, либо укорачивания, и между этими фазами существуют резкие переходы. Эти переходы называются катастрофа, если происходит переключение от роста к укорачиванию, и спасение, если укорачивание сменяется ростом.
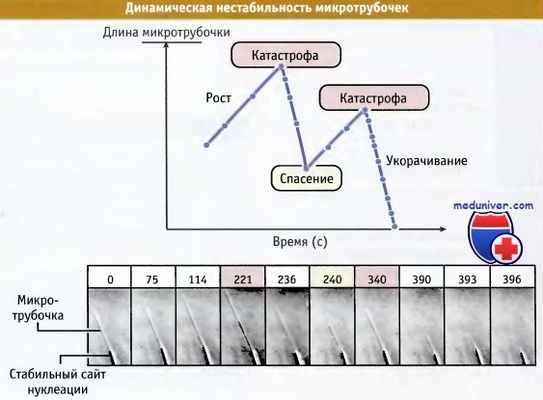
Одиночная микротрубочка растет от стабильного сайта нуклеации in vitro.
Минус-конец присоединен к нуклеирующей структуре, а плюс-конец свободен.
Рост микротрубочки регистрируется во времени с помощью микровидеосъемки. Процесс также представлен графически.
Вначале микротрубочка растет, затем начинает резко укорачиваться. После достижения минимальной длины она начинает расти снова.
Вскоре после этого происходит вторая катастрофа, и микротрубочка полностью разбирается.
Отметим, что разборка происходит в несколько раз быстрее, чем рост. Сверху каждой видео рамки обозначено время в сек.
Модель динамической нестабильности была подтверждена исследованиями с использованием метода световой микроскопии при наблюдении роста и укорачивания индивидуальных микротрубочек. На рисунке ниже представлен такой эксперимент. В этих экспериментах было показано, что индивидуальная микротрубочка дорастает до длины несколько микрон, после чего наступает катастрофа и трубочка быстро укорачивается. Может наступить спасение, в результате чего микротрубочка возобновит рост, или она может полностью диссоциировать. На рисунке ниже представлены два фрагмента видеозаписи микротрубочек, находящихся в фазе перехода между ростом и диссоциацией. Показаны две микротрубочки, которые предварительно получили из стабильной структуры (короткого фрагмента аксонемы жгутика — органеллы, состоящей из стабильных микротрубочек), одна растет на плюс-конце, а другая на минус-конце.
Эти эксперименты показывают, что плюс концы микротрубочек растут быстрее, чем минус. Также на плюс-концах чаще происходит катастрофа, что делает их более динамичными по сравнению с минус концами.
Существенным признаком динамической нестабильности микротрубочек является преимущественное состояние их элонгации или диссоциации, иногда прерываемое переходами между ними. Поскольку катастрофы и спасения наступают через произвольные интервалы времени, поведение отдельных микротрубочек в группе носит гетерогенную и асинхронную природу. Большинство микротрубочек будут расти медленно, но в то же время некоторые будут расти быстро. Отдельные микротрубочки не достигают равновесной длины; а каждая из них обычно становится длиннее или короче.
Структура концов растущих и диссоциирующих микротрубочек различна. Концы микротрубочек при полимеризации часто образуют листообразные выступы за счет того, что некоторые протофиламенты оказываются длиннее других. Субъединицы добавляются только к концам протофиламентов. По мере элонгации каждого протофиламента, субъединицы образуют боковые связи с соседними, и в конце концов выступы превращаются в структуру микротрубочки. При деполимеризации микротрубочки, концы отдельных протофиламентов закручиваются в кольца, внутренней стороной наружу. Субъединицы в отдельном, частично деструктурированном протофиламенте держатся вместе только за счет продольных связей.
Как обсуждалось в отдельных статьях на сайте, связи между субъединицами в пределах одного протофиламента нарушаются с одинаковой вероятностью, приводя к быстрой деградации протофиламентов за счет диссоциации концевых субъединиц и образование разломов в других местах.
В качестве стабильного сайта нуклеации при сборке очищенного тубулина in vitro использовали небольшой участок аксонемы жгутика.
Показаны два видео кадра. На верхнем кадре микротрубочка полимеризуется на каждом конце аксонемы.
У микротрубочки слева свободен плюс-конец; у микротрубочки справа — минус-конец.
За время между съемкой верхнего и нижнего кадров левая микротрубочка диссоциировала полностью; микротрубочка справа диссоциировала частично, и затем снова начала рост. Сверху представлены микрофотографии плюс-концов растущих и диссоциирующих микротрубочек, сделанные в электронном микроскопе.
С одной стороны конца растущей микротрубочки выходят длинные протофиламенты.
Эти протофиламенты расположены на плоской поверхности, к которой были присоединены микротрубочки при приготовлении препарата.
Поэтому на конце протофиламенты видны расположенными в плоскости. На диссоциирующем конце протофиламенты отсоединяются от стенки микротрубочки и закручиваются в обратную сторону.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Биология клетки/Часть 1. Клетка как она есть/7/6
Микротрубочки - трубочки диаметром около 24-25 нм, состоящие из белка тубулина. Микротрубочки есть во всех клетках эукариот и могут собираться в цитоплазме, а у многих эукариот и внутри ядра. Микротрубочки:
- Участвуют в поддержании формы клеток
- Влияют на взаимное расположение органелл (например, ЭПС и аппарата Гольджи)
- Обеспечивают транспорт веществ (по их поверхности, как по рельсам, мс помощью двигательных белков кинезинов и динеинов могут передвигаться мембранные пузырьки с грузом и макромолекулы)
- Составляют основу жгутиков и ресничек эукариот, при участии динеинов обеспечивают ресничное и жгутиковое движение
- Составляют основу центриолей, которые во многих клетках эукариот выполняют роль ЦОМТ - центра организации микротрубочек
- Образуют веретено деления и обеспечивают расхождение хромосом к полюсам клетки в ходе ее деления
Содержание
Тубулин — глобулярный белок. В клетках эукариот есть несколько слегка различающихся генов-ортологов, которые кодируют три разных формы тубулина — α-тубулин, β-тубулин и γ-тубулин. У каждого из этих белков выделяют три домена. Одна молекула α-тубулина и одна молекула β-тубулина в цитоплазме клеток объединяются в димер (изображен на рисунки). В составе такого димера к каждой молекуле тубулина присоединено по одной молекуле ГТФ.
На самом деле каждую из форм тубулина кодирует не один ген, а около десятка, так что каждая форма существует в виде множества подтипов. Они могут быть специфичны для определенных тканей. Например, тубулин III-β встречается только в нейронах. Кроме того, по одному гену кодируют еще две формы тубулина — δ- и ε-тубулин, которые, как и γ-тубулин, встречаются в основном в центриолях.
Новые молекулы тубулина могут прикрепляться как к ±, так и к --концу микротрубочки, но легче (при более низкой концентрации) присоединяются к + -концу (и легче отделяются от --конца).
Тубулин способен связываться в растворе с молекулами ГТФ. Рост микротрубочек осуществляется только за счет присоединения димеров тубулина, в которых обе субъединицы связаны с молекулами ГТФ. В стенках микротрубочек происходит гидролиз ГТФ, связанной с β-субъединицей, до ГДФ (связанная с α-субъединицей ГТФ стабильна). Связанная с ГДФ форма тубулина легче отделяется от микротрубочек, что определяет динамическую нестабильность микротрубочек — при определенных условиях они быстро распадаются почти полностью.
Микротрубочка распадается, когда расщепление ГТФ происходит быстрее, чем присоединение новых димеров. Тогда защитная "шапочка" (кэп) на +-конце из димеров, связанных с ГТФ, разрушается, и после эого быстро отваливаются от микротрубочки димеры, связанные с ГДФ.
[[1]] Анимация сборки и разборки микротрубочки
По поверхности микротрубочек, как по рельсам, могут транспортироваться мембранные пузырьки (везикулы) с грузом или макромолекулы. Их перемещение осуществляют две группы моторных белков - кинезины и динеины. Кинезины перемещаются по микротрубочке от --конца к +-концу, то есть обеспечивают транспорт к периферии клеток. Динеины двигаются в противоположном направлении - от +-конца микротрубочки к --концу, то есть обычно перемещают грузы к центру клетки.
Транспорт к периферии клетки часто называют антероградным транспортом, транпорт к центру - ретроградным (особенно эти термины применимы к движению по отросткам клеток - например, аксонам или дендритам. Одно из семейств кинезинов (кинезины-14, в том числе NCD дрозофилы, KAR3 почкующихся дрожжей и ATK5 арабидопсиса, двигаются по микротрубочкам от +-конца к --концу.
[2] Строение и функции кинезинов и динеинов (англ.)
Реснички и жгутики — органеллы, которые есть у многих групп эукариот. Это тонкие (диаметром 0,1-0,6 мкм) волосковидные выросты на поверхности клетки. Длина их составляет от 3-15 мкм до 2 мм (реснички гребных пластинок гребневиков).
Жгутики и реснички всех эукариот имеют сходное строение, поэтому иногда их обозначают общим термином ундулиподии. Это сходство — одно из свидетельств единого (монофилетического) происхождения эукариот. Ундулиподии есть у большинства групп протистов, у споровых и некоторых голосеменных растений раст и практически у всех животных. Настоящие грибы (Eumycota) лишены жгутиков и ресничек.
Различие между жгутиками и ресничками на самом деле заключается в характере их движения: реснички совершают удары в одной плоскости, а жгутик вращается (одновременно по нему может пробегать волна). Но в зоологии принято называть жгутиковыми любые клетки, у которых есть один подвижный жгутик (или ресничка). При наличии многочисленных ресничек клетки называют ресничными.
У многих беспозвоночных животных ими покрыта вся поверхность тела (ресничные черви, личинки кишечнополостных и губок) или отдельные его участки (например, жабры у полихет и двустворчатых моллюсков, подошва ноги у брюхоногих моллюсков). У коловраток из специализированных ресничек состоит коловращательный аппарат. У многих беспозвоночных (кишечнополостные, гребневики, турбеллярии и др.) реснички также имеются на клетках кишечного эпителия.
У позвоночных (в том числе человека) клетки с подвижными ресничками также есть во многих органах. У человека ресничным эпителием выстланы дыхательные пути, евстахиевы трубы, семявыносящие канальцы, желудочки мозга и спинномозговой (центральный) канал. Видоизмененные реснички служат световоспринимающим аппаратом фоторецепторов сетчатки глаза и воспринимающим запахи аппаратом хеморецепторов обонятельного эпителия.
Подвижные реснички и жгутики либо служат для движения, либо создают токи жидкости. Эти токи могут приносить пищу или кислород из внешней среды или служить для транспорта растворенных веществ внутри организма. Неподвижные жгутики и реснички (часто сильно видоизмененные) служат рецепторами у самых разных групп организмов.
Биофизик РУДН смоделировал поведение элементов микротрубочек клетки для химического воздействия на их рост и распад
Биофизик РУДН смоделировал молекулярную динамику роста важнейших элементов жизнедеятельности клетки — микротрубочек. Исследователи построили модель взаимодействия субъединиц микротрубочек с учётом их внутренних и внешних связей. Результаты позволяют сформировать более полную модель динамической нестабильности микротрубочек. Это позволит подобрать химические агенты для терапии некоторых заболеваний, в том числе новообразований и нейродегенеративных патологий.
Микротрубочки — важный компонент цитоскелета, отвечающий за деление и развитие клетки. Структурная единица микротрубочки — димер тубулина, который собирается в более крупный компонент — нить протофиламента. В структуре димера тубулина также выделяют два типа молекул — ГТФ (гуанинтрифосфат) и ГДФ (гуаниндифосфат). Сейчас считается, что процесс замены молекул ГТФ на ГДФ влияет на динамику процессов сборки и разборки микротрубочки. Эти процессы приводят к так называемой динамической нестабильности микротрубочек, которая определяет транспорт веществ внутри клетки и процесс разделения хромосом при делении клетки.
Илья Коваленко из РУДН, доктор физико-математических наук, разработал компьютерную модель на основе молекулярной и броуновской динамики с учётом внешних и внутренних взаимодействий димеров тубулина. Каждый димер вовлечён в два типа внешнего взаимодействия: боковое (взаимодействие между соседними димерами по кольцу микротрубочки) и продольное (взаимодействие между соседними димерами вдоль нити микротрубочки). Модель позволяет визуализировать в динамике форму как отдельных протофиламентов (тубулиновых нитей) разной длины, так и протофиламентов, включённых в состав стенки микротрубочек.
«Броуновская динамика позволяет моделировать крупные структуры на больших временных интервалах, а молекулярная динамика позволяет наряду с электростатическими взаимодействиями учесть взаимодействия с молекулами растворителя, а также локальные и глобальные изменения тубулина» — Илья Коваленко, биофизик РУДН.
Микротрубочки довольно прочны и по своей жёсткости сравнимы с органическим стеклом. На степень жёсткости микротрубочки влияет процентное соотношение димеров с ГТФ и ГДФ димерами тубулина. ГДФ-связанные димеры всегда стремятся выгнуться из стенки микротрубочки. При этом нарушится целостность стенки и начнётся процесс её разборки, который непосредственно повлияет на степень жёсткости микротрубочки. От этих искривлений ГДФ-димеров спасают ГТФ-димеры, которые «натягивают» ГДФ-димеры и не позволяют ему выгибаться. Ранее экспериментально было показано влияние присутствия ГТФ-димеров на жёсткость микротрубочки, однако детальные механизмы этого процесса остаются неясными.
Модель, разработанная Ильей Коваленко вместе с коллегами, продемонстрировала влияние длины свободной нити протофиламента на степень её жёсткости: наращивание нити на основе димера тубулина со временем приводит к отклонению и закручиванию только концевой области нити от положения центра масс димеров. Такое изменение формы связано с влиянием продольных внешних связей между тубулиновыми димерами. Это наблюдается для димеров как с ГТФ-, так и с ГДФ-молекулами.
Также результатом моделирования стал вывод о влиянии типа молекулы (ГТФ или ГДФ) на степень гибкости внешних, но не внутренних связей для отдельной нити. До момента замены в димере тубулина ГТФ на ГДФ его внешние связи слабее внутренних на величину связи с одной фосфатной группой. При встраивании новых димеров тубулина в стенку микротрубочки ей энергетически выгоднее выпрямлять свободные искривлённые протофиламенты с ГТФ-молекулами, чем более жёсткие с ГДФ-молекулами.
Однако различие в степени сопротивления протофиламентов к выпрямлению для димеров с ГТФ и ГДФ молекулами не столь значительно, как считалось ранее. Это ставит под сомнение представление о решающей роли процесса замены ГТФ молекул на ГДФ в димере тубулина.
Если добавить во внутреннюю среду клетки препараты, замедляющие процесс присоединения нитей димеров тубулина не только с ГТФ-молекулами, но и с ГДФ-молекулами, можно добиться замедления роста микротрубочек. Такое химическое управление трубочками позволяет проводить терапию новообразований и нейродегенеративных заболеваний.
Динамическая нестабильность микротрубочек
Организация клетки. Цитоскелет
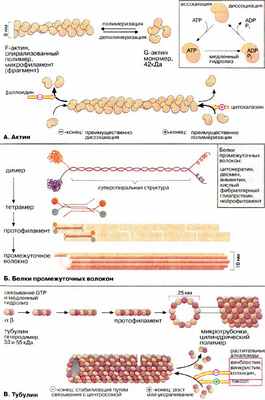
Цитоплазма эукариотических клеток пронизана трехмерной сеткой из белковых нитей (филаментов), называемой цитоскелетом. В зависимости от диаметра филаменты разделяются на три группы: микрофиламенты (6-8 нм), промежуточные волокна (около 10 нм) и микротрубочки (около 25 нм). Все эти волокна представляют собой полимеры, состоящие из субъединиц особых глобулярных белков.
Микрофиламенты (актиновые нити) состоят из актина — белка, наиболее распространенного в эукариотических клетках. Актин может существовать в виде мономера ( G-актин , «глобулярный актин») или полимера ( F-актин , «фибриллярный актин»). G-актин — асимметричный глобулярный белок (42 кДа), состоящий из двух доменов. По мере повышения ионной силы G-актин обратимо агрегирует, образуя линейный скрученный в спираль полимер, F-актин. Молекула G-актина несет прочно связанную молекулу АТФ (АТР), которая при переходе в F-актин, медленно гидролизуется до АДФ (ADP), т.е. F-актин проявляет свойства АТФ-азы.
При полимеризации G-актина в F-актин ориентация всех мономеров одинакова, поэтому F-актин обладает полярностью . Волокна F-актина имеют два разноименно заряженных конца — (+) и (-), которые полимеризуются с различной скоростью. Эти концы не стабилизированы специальными белками (как, например, в мышечных клетках), и при критической концентрации G-актина (+)-конец будет удлиняться, а (-)-конец укорачиваться. В условиях эксперимента этот процесс может быть ингибирован токсинами грибов. Например, фаллоидин (яд бледной поганки) связывается с (-)-концом и ингибирует деполимеризацию, в то время как цитохалазин (токсин из плесневых грибов, обладающий свойством цитостатика) присоединяется к (+)-концу, блокируя полимеризацию.
Актинассоциированные белки. В цитоплазме клеток имеются более 50 различных типов белков, которые специфически взаимодействуют с G-актином и F-актином. Эти белки выполняют различные функции: регулируют объем G-актинового пула (профилин), оказывают влияние на скорость полимеризации G-актина (виллин), стабилизируют концы нитей F-актина (фрагин, β-актинин) , сшивают филаменты друг с другом или с другими компонентами (как, например, виллин, α-актинин, спектрин, MARCKS) или разрушают двойную спираль F-актина (гельзолин) . Активность этих белков регулируется ионами Са 2+ и протеинкиназами.
Б. Белки промежуточных волокон
Структурными элементами промежуточных волокон являются белки, принадлежащие к пяти родственным семействам и проявляющие высокую степень клеточной специфичности. Типичными представителями этих белков являются цитокератины, десмин, виментин, кислый фибриллярный глиапротеин [КФГП (GFAP)] и нейрофиламент . Все эти белки имеют в центральной части базовую стержневую структуру, которая носит название суперспирализованной α-спирали (см. кератин, с. 76). Такие димеры ассоциируют антипараллельно, образуя тетрамер. Агрегация тетрамеров по принципу "голова к голове" дает протофиламент . Восемь протофиламентов образуют промежуточное волокно.
В отличие от микрофиламентов и микротрубочек свободные мономеры промежуточных волокон едва ли встречаются в цитоплазме. Их полимеризация ведет к образованию устойчивых неполярных полимерных молекул.
Микротрубочки построены из глобулярного белка тубулина , представляющего собой димер α- и β-субъединиц (53 и 55 кДа). α, β- Гетеродимеры образуют линейные цепочки, называемые протофиламентами. 13 протофиламентов образуют циклический комплекс. Затем кольца полимеризуются в длинную трубку. Как и микрофиламенты, микротрубочки представляют собой динамические полярные структуры с (+)- и (-)-концами. (-)-Конец стабилизирован за счет связывания с центросомой (центр организации микротрубочек), в то время как для (+)-конца характерна динамическая нестабильность. Он может либо медленно расти, либо быстро укорачиваться. Тубулиновые мономеры связывают ГТФ (GTP), который медленно гидролизуется и ГДФ (GTP). С микротрубочками ассоциируют два вида белков: структурные белки (MAP от англ. microtubuls-associated proteins) и белки-транслокаторы.
Читайте также: