Фолликулярные клетки. Физиология фолликулярных клеток
Добавил пользователь Валентин П. Обновлено: 27.01.2026
Щитовидная железа, расположенная на передней поверхности шеи сразу под перстневидным хрящом, состоит из 2 соединенных перешейком долей. Фолликулярные клетки щитовидной железы вырабатывают 2 основных тиреоидных гормона:
Тетрайодтиронин (тироксин, T4)
Эти гормоны действуют практически на все ткани организма, присоединяясь к ядерным рецепторам и меняя экспрессию многих генных продуктов. Тиреоидные гормоны необходимы для нормального развития головного мозга и соматических тканей у плода и новорожденного и в любом возрасте регулируют белковый, углеводный и жировой обмен.
Наиболее активным тиреоидным гормоном, который прочнее связывается с ядерными рецепторами, является T3; T4 обладает минимальной гормональной активностью. Однако T4 действует гораздо более продолжительное время и (в большинстве тканей) может превращаться в T3. являясь, таким образом, источником T3. Третий тиреоидный гормон, реверсивный T3 (rT3), лишен метаболической активности; уровень rT3 повышается при некоторых патологических состояниях.
Синтез и секреция тиреоидных гормонов
Синтез тиреоидных гормонов требует присутствия йода (см. рисунок Синтез тиреоидных гормонов Синтез тиреоидных гормонов ). Йод, попадающий в организм с пищей и водой в виде йодида, активно концентрируется щитовидной железой и в фолликулярных клетках под действием тиреоидной пероксидазы превращается в органический йод (органификация). Фолликулярные клетки окружают пространство, заполненное коллоидом, который состоит из тиреоглобулина – гликопротеина, содержащего в своем составе тирозин. Тирозин, контактируя с мембраной фолликулярных клеток, йодируется с образованием 1 (монойодтирозин) или 2 (дийодтирозин) продуктов, затем распадается на несколько частей, образовывая 2 формы тиреоидных гормонов.
Дийодтирозин + дийодтирозин → T4
Дийодтирозин + монойодтирозин → T3
Синтез тиреоидных гормонов
T3 и T4 остаются в составе молекул тиреоглобулина внутри фолликула до тех пор, пока фолликулярные клетки не начнут поглощать тиреоглобулин в виде коллоидных капель. В фолликулярных клетках происходит выщепление T3 и T4 из тиреоглобулина.
Свободные T3 и T4 затем высвобождаются в кровоток, где связываются с транспортными сывороточными белками. Первичным транспортным белком является тироксинсвязывающий глобулин (ТБГ), который обладает высокой аффинностью к T3 и T4, но при этом низкой связывающей способностью. В норме ТСГ транспортирует около 75% связанных тиреоидных гормонов.
К другим связывающим белкам относятся
Тироксинсвязывающий преальбумин (транстиретин), который обладает высокой аффинностью к T4, но при этом низкой связывающей способностью.
Альбумин, который имеет низкое сродство, но высокую функциональную активность в отношении T3 и T4
Около 0,3% от общего сывороточного T3 и 0,03% от общего сывороточного T4 являются свободными и находятся в равновесии со связанными гормонами. Только свободный T3 и свободный T4 могут воздействовать на периферические ткани.
Все реакции синтеза и секреции T3 и T4 контролируются тиреотропным гормоном (ТТГ), который секретируется тиреотрофными клетками гипофиза. Секреция ТТГ гипофизом контролируется механизмом отрицательной обратной связи: повышенный уровень свободных T4 и T3 тормозит синтез и секрецию ТТГ, тогда как сниженный их уровень стимулирует секрецию этого гипофизарного гормона. На секрецию ТТГ влияет также тиреотропин-рилизинг гормон (ТРГ) , который синтезируется в гипоталамусе. Точные механизмы регуляции синтеза и секреции ТРГ неизвестны, но тиреоидные гормоны по механизму обратной связи ингибируют его синтез.
Бóльшая часть присутствующего в крови T3 образуется вне щитовидной железы путем монодейодирования T4. Лишь 1/5 количества T3 непосредственно секретируется щитовидной железой.
Лабораторные методы оценки функции щитовидной железы
Измерение уровня тиреотропного гормона (ТТГ)
Наиболее надежным способом выявления дисфункции щитовидной железы является определение уровня ТТГ (см. таблицу Результаты тестов для определения функции щитовидной железы в различных клинических ситуациях Результаты тестов для определения функции щитовидной железы в различных клинических ситуациях ). Нормальный его уровень, как правило, позволяет исключить гипертиркеоз Гипертиреоз Гипертиреоз характеризуется гиперметаболизмом и повышенными уровнями свободных тиреоидных гормонов в сыворотке. Среди многочисленных симптомов обычно имеют место тахикардия, слабость, потеря. Прочитайте дополнительные сведения , за исключением случаев центрального гипотиреоза (обусловленного патологией гипоталамуса или гипофиза) или редких случаев резистентности к тиреоидным гормонам. При очень тяжелых нетиреоидных заболеваниях уровень ТТГ в сыворотке может быть низким, даже в отсутствие гипертиреоза. По уровню ТТГ в сыворотке судят также о наличии субклинического гипертиреоза Субклинический гипертиреоз Гипертиреоз характеризуется гиперметаболизмом и повышенными уровнями свободных тиреоидных гормонов в сыворотке. Среди многочисленных симптомов обычно имеют место тахикардия, слабость, потеря. Прочитайте дополнительные сведения (повышенный уровень ТТГ); оба этих состояния характеризуются нормальными уровнями сывороточного T4, свободного T4, сывороточного T3 и свободного и T3.
Измерение тироксина (T4)
Уровень общего T4 в сыворотке отражает суммарное содержание связанного и свободного гормона. Изменение концентрации белков, связывающих тиреоидные гормоны, сопровождается соответствующим изменением уровня общего T4, тогда как содержание более активного T4 не меняется. Таким образом, несмотря на повышенный уровень общего T4 в сыворотке, пациент может находиться в физиологически нормальном, эутиреоидном состоянии. Прямое определение содержания свободного T4 позволяет избежать ошибок в интерпретации повышения общего уровня T4.
Расчет индекса свободного T4 является расчётной величиной и вносит в показатели общего T4 поправку на различное содержание ТГ-связывающих белков в сыворотке и, таким образом, дает возможность оценить уровень свободного T4 при имеющихся показателях общего T4. Для оценки связывания тиреоидных гормонов с белками сыворотки используют показатель поглощения T4. Индекс свободного T4 определять легче, и он хорошо согласуется с результатами прямого определения уровня свободного T4.
Определение уровня трийодтиронина (T3)
Также можно определить уровни общего T3 и свободного T3. Поскольку T3 прочно связывается с ТСГ (хотя и в 10 раз слабее, чем T4), общее содержание в сыворотке T3 зависит от колебаний уровня ТСГ и от действия веществ, меняющих связывание гормона с этим белком. Уровни свободного T3 в сыворотке измеряют с помощью тех же самых прямых и косвенных методов (индекс свободного T3), применимых для T4, и используются в основном при исследовании тиреотоксикоза.
Тироксин-связывающий глобулин (ТСГ)
Уровень ТСГ может быть определен. Его концентрация повышена при беременности, при терапии эстрогенами или приеме пероральных контрацептивов и в острой фазе инфекционного гепатита Обзор острых вирусных гепатитов (Overview of Acute Viral Hepatitis) Острый вирусный гепатит – это диффузное воспалительное заболевание печени, вызванное специфическими гепатотропными вирусами, которые имеют разные пути передачи и особенности эпидемиологии. Неспецифический. Прочитайте дополнительные сведения . Повышение уровня ТСГ встречается также при Х-сцепленной патологии. Его снижение чаще всего регистрируется при заболеваниях, сопровождающихся нарушением синтеза белка в печени, при использовании анаболических стероидов и передозировке кортикостероидов. Большие дозы некоторых лекарственных средств (например, фенитоина, аспирина и их производных) вытесняют T4 из связи с ТСГ, что создает ложное впечатление о снижении уровня общего T4 в сыворотке.
Аутоантитела к тиреоидной пероксидазе
Аутоантитела к тиреоидной пероксидазе присутствуют почти у всех больных с тиреоидитом Хасимото Болезнь Хашимото Тиреоидит Хашимото – это хроническое аутоиммунное воспаление щитовидной железы с лимфоцитарной инфильтрацией. Характерны безболезненное увеличение железы и симптомы гипотиреоза. Диагноз основывается. Прочитайте дополнительные сведения (у некоторых из них определяются и антитела к тиреоглобулину), а также у большинства больных с болезнью Грейвса. Эти аутоантитела служат маркерами аутоиммунного процесса, но, по всей вероятности, не являются причиной заболевания. Однако аутоантитела к рецепторам тиреотропного гормона на поверхности фолликулярных клеток щитовидной железы ответственны за развитие гипертиреоза при болезни Грейвса. В сыворотке больных с аутоиммунными заболеваниями щитовидной железы могут присутствовать и антитела к T4 и T3; они влияют на результаты определения уровней T4 и T3, но редко имеют клиническое значение.
Тиреоглобулин
Щитовидная железа является единственным источником тиреоглобулина, который легко обнаружить в сыворотке крови здоровых людей, и он, как правило, повышен у пациентов с нетоксическим или токсическим диффузным зобом. Основное применение показателей определения уровня тиреоглобулина в сыворотке крови состоит в выявлении дифференцированного рака щитовидной железы у пациентов после субтотатьной или полной тиреоидэктомии (с абляцией или без йодом-131). Нормальные или повышенные значения тиреоглобулина в сыворотке крови указывают на наличие остаточной нормальной или раковой ткани щитовидной железы у пациентов, получающих дозы L-тироксина, подавляющие ТТГ, или после отмены L-тироксина. Присутствие антител к тиреоглобулину мешает его точному определению.
Обследование на предмет дисфункции щитовидной железы
Скрининговое обследование каждые 5 лет с определением содержания ТТГ в сыворотке крови рекомендовано всем мужчинам ≥ 65 лет и всем женщинам ≥ 50 лет. Скрининговое обследование также рекомендовано всем новорожденным и беременным. Пациентам с факторами риска тиреоидной патологии уровень ТТГ в сыворотке следует определять чаще. Скрининг на гипотиреоз, равно как и на гипертоническую болезнь, гиперхолестеринемию и рак молочной железы, экономически оправдан. Одно только определение уровня ТТГ с высокой чувствительностью и специфичностью позволяет диагностировать или исключить две распространенные и тяжелые патологии ( гипотиреоз Гипотиреоз Гипотиреоз – это дефицит тиреоидных гормонов. Диагноз устанавливают на основании клинической картины (типичное выражение лица, грубый голос и замедленная речь, сухая кожа) и сниженного уровня. Прочитайте дополнительные сведения ), которые поддаются лечению. Учитывая высокую частоту гипотиреоза среди пожилых людей, лиц старше 70 лет целесообразно обследовать ежегодно.
Исследование захвата и отображения радиоактивного йода щитовидной железой
Измеряют также поглощение щитовидной железой радиоактивного йода. Следовые дозы радиойода вводят внутрь или внутривенно, а затем с помощью сканера определяют его количество, поглощенное щитовидной железой. Преимущественно используют изотоп йод-123, который создает минимальную лучевую нагрузку (гораздо меньшую, чем йод-131). Поглощение йода-123 щитовидной железой сильно зависит от приема йода и у пациентов, потребляющих избыток йода, резко снижено.
Это исследование имеет значение в дифференциальной диагностике гипертиреоза Диагностика Гипертиреоз характеризуется гиперметаболизмом и повышенными уровнями свободных тиреоидных гормонов в сыворотке. Среди многочисленных симптомов обычно имеют место тахикардия, слабость, потеря. Прочитайте дополнительные сведенияПосле введения радиоактивного изотопа (радиойода или 99m-технеция пертехнетата) с помощью визуализации при использовании сцинтилляционной камеры можно получить графическое изображение поглощения изотопа. Обнаружение участков повышенного («горячие» узлы) или сниженного («холодные» узлы) поглощения способствует выявлению возможного ракового поражения (рак присутствует менее, чем в 1% «горячих» узлов и в 10–20% «холодных»).
Авторское право © 2022 Merck & Co., Inc., Rahway, NJ, США и ее аффилированные лица. Все права сохранены.
Фолликулярные клетки. Физиология фолликулярных клеток
Фолликулярные клетки. Физиология фолликулярных клеток
Фолликулярные клетки не только играют роль трофических и опорных элементов, но их совокупность является железой внутренней секреции, вырабатывающей гормон фолликулин (эстрин), который содержится в фолликулярной жидкости. Эстрин через капиллярную сеть, разветвленную во внутреннем слое теки и проникающую даже между фолликулярными клетками (васкуляризированный эпителий в гранулезной мембране), попадает в кровяное русло.
Действие фолликулина будет рассмотрено ниже. В процессе созревания граафова фолликула происходит первое деление созревания, которое, в сущности, можно сравнить с первым делением созревания при спермиогенезе, поскольку при этом также происходит уменьшение количества (редукция) ядерного вещества.
В отличие от спермиогенеза при развитии граафова фолликула из ооцита первого порядка возникают не две полноценные и одинаковые клетки, а два различных по величине и физиологически неравноценных элемента. Одна из клеток остается крупной, она имеет приблизительно такую же величину, как ооцит первого порядка, ее называют ооцитом второго порядка, который аналогичен соответствующей стадии спермиогенеза.
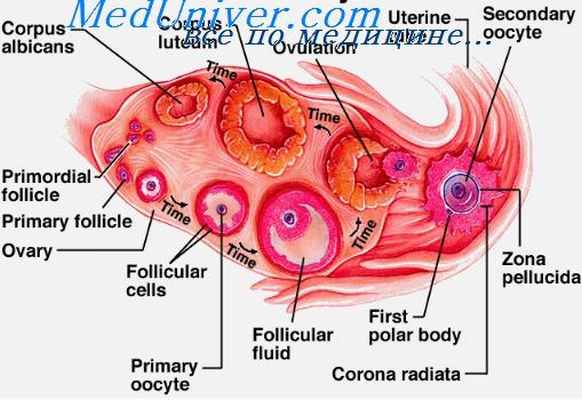
Вторая клетка является рудиментарной, она не увеличивается в размерах, остается недоразвитой, неполноценной. При делении в нее переходит половина ядерного вещества из ооцита I порядка. Эта несовершенная клетка, так называемое первое полярное тельце, располагается на поверхности ооцита в пространстве между ним и оолеммой, а позднее, разделившись иногда на две маленькие клетки, погибает.
Между тем граафов фолликул, объем которого уже значительно увеличился, все больше и больше выдается над поверхностью яичника; в месте его наибольшего выступления (stigma) стенка пузырька истончается. Под давлением фолликулярной жидкости и в результате дегенерации сдавленной ткани на выступающем полюсе (атрофия давления), а также под действием протеолитических энзимов, образующихся в ткани фолликула, наконец происходит разрыв стенки граафова пузырька: фолликулярная жидкость изливается из него струей, увлекая с собой и яйцеклетку с непосредственно окружающими ее оболочками (оолеммой и лучистым венцом). Процесс разрыва фолликула и освобождения из него яйцевой клетки называется овуляцией.
Лишь после овуляции, по всей вероятности, одновременно с оплодотворением яйцеклетки, происходит второе деление созревания, при котором ооцит второго порядка в результате митотического деления снова делится на одну полноценную клетку — зрелую яйцевую клетку и на маленькую рудиментарную клетку — второе полярное тельце (полоцит), прилипшее к поверхности яйцеклетки.
При этом может разделиться и первое тельце, так что в результате обоих последовательных делений созревания возникает одна полноценная крупная клетка (зрелая яйцевая клетка) и три захиревшие, неполноценные клетки (полярные тельца), которые позднее погибают. Таким образом, в отличие от спермиогенеза, при оогенезе из одной оогонии возникает только одна яйцевая клетка, которая является уже зрелой, способной после оплодотворения к дальнейшему развитию, в то время как гомологичный сперматид должен еще пройти трансформационным, преобразующим процессом спермиогистогенеза для того, чтобы стать функционально и морфологически зрелым спермием.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
ФИЗИОЛОГИЯ ЖЕНСКОГО ЗДОРОВЬЯ
Каждый месяц организм женщины претерпевает циклические изменения, связанные с половым циклом. Это проявляется колебаниями уровней половых гомонов, и связанных с ними изменениями в так называемых «органах-мишенях» — это, прежде всего, половые органы, молочные железы, а также центральная нервная система.
Регуляция этого процесса осуществляется тонко отлаженной работой системы органов, которая реагирует на любое изменение в организме – заболевание, стресс, неправильное питание, вредные зависимости и т.д.
Первым звеном в цепи регуляции выступают отделы головного мозга – гипоталамус и гипофиз. Гипоталамус, участвующий в регуляции циркадных ритмов, т.е. в настройке «внутренних часов организма», стимулирует в гипофизе выработку гонадотропных гормонов – фолликулостимулирующего (ФСГ) и лютеинизирующего (ЛГ). Попадая в кровь, они достигают яичников и вызывают в них изменения, приводящие к началу синтеза половых гормонов – эстрогенов и прогестерона, которые в свою очередь влияют на «органы-мишени». Это влияние обуславливает регулярность полового цикла, т.е. реализацию детородной функции женщины, а также изменения в некоторых органах, не относящихся к репродуктивным, например – коже, жировой ткани, центральной нервной системе и др.
Что происходит в женском организме во время менструального цикла?
В данной статье мы рассмотрим изменения, касающиеся репродуктивной функции. Для облегчения понимания процессов, происходящих в течение полового цикла, его принято делить на 2 фазы: первую — фолликулярную и вторую — лютеиновую.
Фолликулярная фаза: длится с первого дня менструального кровотечения и до овуляции. Продолжительность её может различаться у разных женщин и зависит от общей длительности менструального цикла.
Лютеиновая фаза: занимает период с момента овуляции до первого дня следующей менструации и у всех женщин длится 2 недели. Общая длительность менструального цикла в среднем составляет 28 дней, но может варьировать от 24 до 38 дней.
Фолликулярная фаза
В яичнике половые клетки женщины – ооциты, или яйцеклетки — находятся в фолликулах — структурах, обеспечивающих их питание и созревание. ФСГ, вырабатываемый гипофизом, стимулирует рост фолликулов – начинается первая (фолликулярная) фаза цикла.
Во время каждого менструального цикла в рост вступает около 20 фолликулов, но полного созревания достигает только один – доминантный, остальные фолликулы гибнут. В отличие от сперматозоидов, которые постоянно продуцируются в мужском организме, ооциты к делению не способны и погибшие клетки не восстанавливаются новыми. Гибель ооцитов связана как с менструальным циклом, так и с причинами, от него не зависящими – гинекологические и общие заболевания, операции, воздействие лекарств и химических веществ. В настоящий момент известны не все причины гибели половых клеток, но установлено, что с течением жизни женщины их количество в яичниках значительно уменьшается.
Под воздействием ФСГ фолликулы начинают продуцировать эстрогены. Чем больших размеров достигает фолликул, тем больше эстрогенов он продуцирует. Соответственно уровень эстрогенов в первую фазу цикла постепенно нарастает. Эти гормоны отвечают за разрастание и утолщения эндометрия — слизистой оболочки полости матки (при УЗИ врач диагностирует утолщение эндометрия), усиление продукции слизи в шейке матки (выделения из влагалища становятся обильными, прозрачными и тягучими). Эти изменения призваны облегчить проникновение сперматозоидов в полость матки и подготовить организм женщины к возможному оплодотворению. Достигая определенной концентрации, эстрогены вызывают выброс ЛГ гипофизом, это приводит к разрыву стенки доминантного фолликула и выходу из него яйцеклетки – овуляции. Яйцеклетка захватывается фимбриями маточных труб, и попадает в их полость, где завершается её созревание.
Лютеиновая фаза
Тем временем, на месте разорвавшегося фолликула под действием ЛГ образуется желтое тело, состоящее из клеток, продуцирующих прогестерон. Функция прогестерона – сохранение жизнеспособности плодного яйца и поддержание беременности в течение первых 3 месяцев при возможном оплодотворении. Он вызывает перестройку сосудов эндометрия (эти изменения также видны при УЗИ), снижение продукции слизи в шейке матке, расслабление мускулатуры стенки матки и маточных труб, разрастание выводных протоков молочных желез (может наблюдаться их незначительное увеличение и нагрубание во второй фазе цикла). При наступлении беременности желтое тело, под действием гормонов, продуцируемых оплодотворенным плодным яйцом, сохраняется до 3 месяцев, продолжая выделять прогестерон.
Если же оплодотворение не произошло, то со временем происходит обратное развитие желтого тела, уровень прогестерона начинает снижаться. В отсутствии гормона в эндометрии начинаются деструктивные изменения, нарушается его питании, он отторгается от стенки матки и выводится из ее полости вместе с неоплодотворенной яйцеклеткой – начинается менструация. В это время концентрация половых гормонов в крови женщины минимальна, гипофиз отвечает на это выработкой ФСГ – вторая фаза цикла сменяется первой.
Таким образом, каждый месяц организм женщины подготавливается к наступлению беременности. Регуляция этого процесса очень сложна, в данной статье мы рассмотрели только основные факторы, принимающие участие в ежемесячной перестройке женского организма.
Здоровый образ жизни, правильное питание, рациональные физические нагрузки и отказ от вредных привычек необходимы для поддержания нормального менструального цикла, хорошего самочувствия и здоровой беременности.
РОСТ И РАЗВИТИЕ ФОЛЛИКУЛОВ
Ооцит млекопитающих развивается в составе фолликула — основной функциональной единицы яичника. Фолликулярные клетки, окружающие ооцит, изолируют его от окружающей среды. Через плотные контакты из фолликулярных клеток поступают питательные вещества и биологически активные вещества, необходимые для роста и созревания фолликулов. Еще в раннем пренатальном онтогенезе герминативный эпителий яичников млекопитающих дифференцируется на оогонии и фолликулярные клетки. Предполагается, что именно первичные фолликулярные клетки тормозят митотическое размножение оогоний и вызывают начало мейоза. В ходе пролиферации и дифферен- цировки однослойный фолликулярный эпителий преобразуется в многослойный, выделяется прилегающая к ооциту зона гранулезных клеток, а позднее — базальная мембрана (неклеточное соединительнотканное образование, разделяющее зоны гранулезы и теки). Рост фолликула сопровождается параллельным ростом ооцита, накоплением в нем белков, нуклеиновых кислот, ферментов, кортикальных гранул, других цитоплазматических включений. Фолликулярные клетки участвуют также в образовании жидкости, заполняющей формирующуюся внутри фолликула полость, а клетки гранулезы, прилегающие к ооциту (клетки яйце-
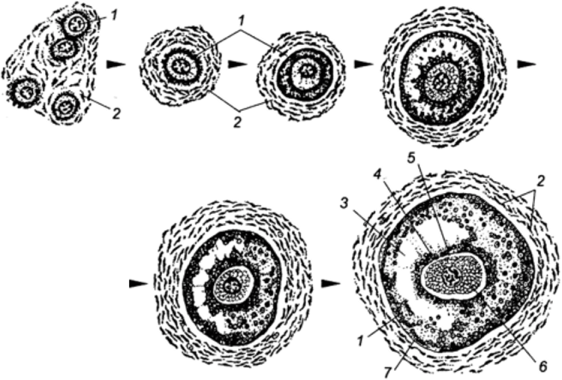
Рис. 9 Основные стадии развития фолликула (Carlson, 1981; по Гилберт С., 1993):
1 — клетки грануле- зы; 2 — клетки теки; 3 — полость; 4 — corona radiata; 5 — zona pellucida; 6 — ооцит; 7 — гранулеза.
носного бугорка, или кумулюса) — в образовании вокруг яйцеклетки блестящей оболочки, состоящей из гликопротеидов.
К моменту полового созревания у самок в корковом слое яичников присутствует большое количество различных по размерам и фазам роста первичных (примордиальных) и развивающихся фолликулов. Развитие фолликулов и яйцеклеток является циклическим процессом. Одновременно развивается один или несколько фолликулов и соответственно одна или несколько яйцеклеток. Выделяют несколько стадий развития фолликула (рис. 9).
В яичнике новорожденных самок присутствуют первичные фолликулы. Количество их достигает сотни тысяч. Каждый из них состоит из половой клетки (ооцита первого порядка), окружающего ее одного слоя фолликулярных клеток и соединительнотканной оболочки — теки. Во время развития эпителий фолликула из плоского становится сначала кубическим, а затем призматическим. Размножаясь, фолликулярные клетки окружают ооцит первого порядка сначала в два, а затем в несколько слоев. В результате формируются вторичные фолликулы.
В дальнейшем в центре фолликула появляется наполненная жидкостью полость, образующаяся из слившихся небольших наполненных жидкостью полостей между фолликулярными клетками. Вторичный фолликул превращается в третичный (антральный) фолликул, или Граафов пузырек. В процессе развития фолликула происходит рост ооцитов. Они покрываются блестящей оболочкой, которая состоит из мукополисахаридов и является продуктом обмена веществ ооцита. Вокруг ооцита фолликулярный эпителий располагается в виде правильных радиальных лучей (лучистый венец). Эти клетки выполняют по отношению к ооциту трофическую (питательную) роль. По мере роста зрелый фолликул начинает выпячиваться над поверхностью яичника в виде прозрачного пузырька.
Развитый фолликул представляет собой пузырек, центр которого заполнен жидкостью. Полость окружена зоной фолликулярных клеток, располагающихся в 10-12 слоев. На участке, где находится яйцеклетка, фолликулярные клетки образуют утолщение, которое вдается в полость фолликула в виде бугорка. На его вершине расположена окруженная лучистым венцом яйцеклетка.
Снаружи фолликул покрыт соединительнотканной оболочкой, образованной из стромы яичника. В ней различают два слоя: внутренний, прилегающий к клеткам гранулезы, и наружный — к строме яичника. Во внутреннем слое имеется много мелких кровеносных сосудов и клеток. Он отделен от клеток гранулезы соединительнотканной мембраной. Клетки внутренней оболочки в строении сходны с клетками желтого тела, поэтому их иногда называют текалютеиновыми. Клетки внутреннего слоя служат основным местом выработки прогестерона, андрогенов и частично эстрогенов. Наружный слой оболочки беден кровеносными сосудами. В том месте, где фолликул выступает над поверхностью яичника, он покрыт снаружи также белочной оболочкой яичника и слоем зародышевого эпителия.
Из растущих фолликулов только часть развивается полностью. Большинство из них погибает на разных стадиях развития. Это явление носит название атрезии фолликулов. Учитывая, что атрезия фолликулов происходит постоянно, в последнее время большинство исследователей приходит к выводу, что она является физиологическим явлением, необходимым для нормального течения циклических процессов в яичниках. В яичнике млекопитающих число генеративных клеток в сотни раз превышает число потомков, которых может выносить и выкормить животное.
Большинство овариальных фолликулов на разных стадиях развития подвергается атрезии. Например, в яичниках половозрелых коров атрезией затронуто 69,8% однослойных фолликулов и 10-41% многослойных. По данным ряда исследователей при рождении в яичниках собак насчитывается 700 000, при наступлении половой зрелости — 250 000, в возрасте 5 лет — 33 000, в возрасте 10 лет — 500 первичных фолликулов (МсБопаИ, 1980; по Дюльгер, 2002).
Сейчас уже известно, что атрезия фолликулов происходит путем апоптоза. Сначала погибает ооцит, что приводит к образованию неправильной формы фолликула, состоящего только из клеток гранулезы. Это позволяет предположить, что ооцит имеет свой собственный генетический механизм апоптоза. Судьба фолликула зависит от баланса экспрессии апоптотических и антиапоптотических факторов. Среди этих факторов протеины Ьс1-2 и Ьах играют основную роль. С одной стороны, у мышей, лишенных гена Ьс1-2, снижается число фолликулов, присутствующих после рождения, а по- вышеная экспрессия этого белка приводит к торможению апоптоза фолликулов. С другой стороны, дефицит Ьах приводит к продолжению репродуктивного периода. В целом исследования показывают, что даже если эти протеины вовлечены в регуляцию апоптоза фолликулов, фактор, инициирующий каскад апоптоза в физиологических условиях, остается неизвестным.
По-видимому, первичные изменения в атретичных фолликулах происходят в ооцитах, что в дальнейшем приводит к перестройке обмена веществ в клетках гранулезы. В клетках гранулезы снижается синтез эстрогенов из тестостерона, и в фолликулах образуется повышенное количество андрогенов, препятствующих нормальному развитию фолликула. Механизм, посредством которого происходит отбор фолликула для овуляции — одна из интригующих и еще не разрешенных тайн биологии.
Для каждого вида характерна определенная частота овуляции, на которую влияют генетические, сезонные, пищевые и некоторые другие внешние факторы. Вероятно, рост фолликула может происходить в строго определенных гормональных условиях, причем эти условия меняются в зависимости от стадии развития фолликула. Поскольку фолликулы растут асинхронно, очевидно, что в каждый из отдельно взятых дней секреция гонадотропинов будет оптимальной для ограниченного числа фолликулов. У человека и у других видов, для которых характерна одиночная овуляция, происходит отбор только одного фолликула.
Известно, что в фолликулярной жидкости крупных и здоровых фолликулов коров и человека содержится больше ФСГ и ЛГ, чем в малых и атретических фолликулах. Предполагают, что избирательное проникновение ФСГ из крови в один из фолликулов является одним из пусковых механизмов выделения «ведущего» фолликула.
У высших приматов отбор этого фолликула, вероятно, осуществляется в течение первых нескольких дней после начала менструации. После того, как отбор уже произошел, фолликул, который в конечном итоге будет овулировать, становится доминирующим, т. е. он растет и дифференцируется со скоростью, определяемой экспоненциальной зависимостью, тогда как другие полостные фолликулы становятся атретическими. Подсчитано, что у женщин фолликул, который будет овулировать, на 1-й день цикла равен приблизительно 2 мм, ко времени овуляции спустя 14 суток он увеличивается приблизительно до 23 мм. Этот рост сопровождается 100-кратным увеличением объема фолликулярной жидкости, а число гранулезных клеток возрастает с 0,5-10® до 50 10 6 . Доминирующий фолликул секретирует увеличивающиеся количества 17(>-эстрадиола, отчего уровень этого гормона в крови экспоненциально возрастает. Увеличение роста преовуляторного фолликула происходит на фоне падения концентрации ФСГ в периферической крови. Представляется вероятным, что доминирующий фолликул становится относительно менее зависимым от уровня ФСГ в крови, поскольку фолликулярная жидкость теперь сама содержит ФСГ и эстрадиол в высоких концентрациях.
Когда доминирующий фолликул достигает зрелости, секреция эстрадиола достаточно интенсивна для того, чтобы индуцировать эффект положительной обратной связи, вследствие чего происходит массированное выделение гипофизарного ЛГ. ЛГ вызывает ряд изменений в структуре и функции фолликула, а также стимулирует возобновление мейоза в ооците. В конечном итоге эти структурные и функциональные преобразования приводят к разрыву стенки фолликула и освобождению созревающей яйцеклетки — овуляции (рис. 10).
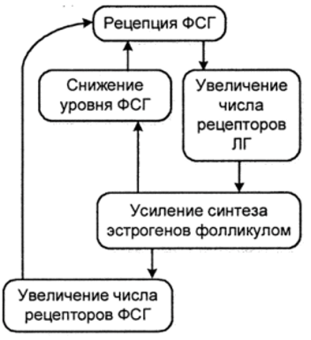
Рис. ю
Цикл с положительной обратной связью в фолликулярных клетках млекопитающих (Гилберт С., 1993)
Каким образом ЛГ нарушает мейотический блок?
Поиски ответа на этот вопрос привели к интенсивному изучению природы блока мейоза. Эта стадия оогенеза очень важна, поскольку в это время ооцит растет, в нем дифференцируются специфические структуры и он приобретает способность завершить деления мейоза. Ранние исследования показали, что заключенный в фолликул ооцит (как in vivo, так и in vitro) без воздействия гонадотропинов не созревает, тогда как ооцит, извлеченный из фолликула, способен возобновить мейоз даже без гормональной стимуляции.
Отсюда можно заключить, что при нормальном развитии мейоз ингибирован фолликулярными клетками, а его инициация вызывается гонадотропинами. Это предположение о регуляции мейоза фолликулярными клетками подтверждается данными о том, что клетки кумулюса связаны с ооцитом посредством своих отростков, пронизывающих блестящую оболочку, и что эти отростки несут щелевые контакты, позволяющие небольшим молекулам проходить между клетками кумулюса и ооцитом.
Наблюдения, свидетельствующие о том, что при повышении уровня цАМФ наблюдается ингибирование созревания ооцита, послужили основанием для предположения, что блок мейоза поддерживается транспортом цАМФ через щелевые контакты из фолликулярных клеток в ооцит. Всплеск синтеза ЛГ мог бы служить триггером созревания, если бы он прерывал коммуникации через щелевые контакты, подавляя таким образом транспорт цАМФ в ооцит. Несколько групп данных подкрепляют эту гипотезу.
Во-первых, снижение концентрации цАМФ играет решающую роль для возобновления мейоза. Дезинтеграция зародышевого пузырька может быть предотвращена, если в выделенных из фолликулов яйцеклетках ингибировать разрушение цАМФ или прямо воздействовать на них цАМФ. Снижение концентрации цАМФ в ооците происходит непосредственно перед возобновлением мейоза. Предполагается, что падение уровня цАМФ приводит к снижению концентрации цАМФ-зависимой протеинкиназы. Этот фермент, по-видимому, отвечает за фосфорилирование особых белков, которые в фосфорилированном состоянии поддерживали бы мей- отический блок. Если ингибиторы этой протеинкиназы инъецировать в ооцит мыши, то мейоз может возобновиться даже в присутствии цАМФ.
Во-вторых, гонадотропины, по-видимому, ответственны за нарушение связи между ооцитом и фолликулярными клетками. Последние служат важным источником цАМФ ооцита, поэтому изменения концентрации цАМФ фолликулярных клеток скажутся на уровне цАМФ ооцита. Этим можно объяснить, почему окруженный фолликулярными клетками ооцит находится в состоянии блокированного мейоза и освобождается от этого блока при удалении фолликулярных клеток.
Всплеск синтеза гонадотропина, однако, может поднять концентрацию цАМФ фолликулярных клеток на новые уровни. В ответ на это повышение зрелые фолликулярные клетки синтезируют гиалуроновую кислоту, которая вызывает физический разрыв контактов между отростками фолликулярных клеток и ооцитом. В результате мостики между фолликулярными клетками гранулезы и ооцитом, по которым
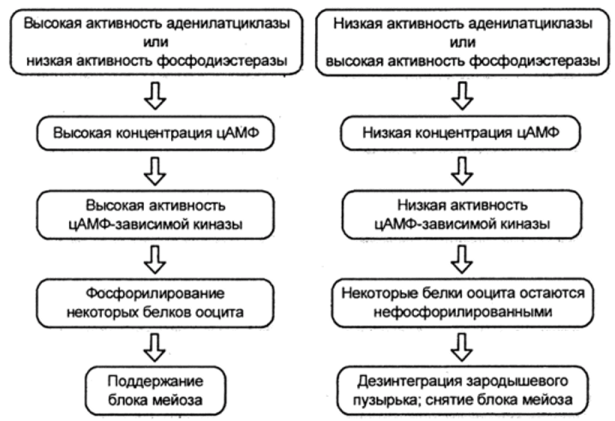
Последовательность событий, приводящих к возобновлению мейоза в ооцитах млекопитающих (Гилберт С., 1993)
перетекает цАМФ, прерываются, и ооцит млекопитающего возобновляет мейоз (рис. 11).
Ооциты являются электрогенными клетками, развитие которых блокируется на определенной стадии мейотическо- го деления. У млекопитающих до приобретения Яйцеклеткой функциональной зрелости сохраняется тесный контакт между ней и окружающими ее клетками кумулюса. Подобные межклеточные взаимодействия осуществляются посредством плотных контактов (gap junctions). Интересно, что, несмотря на такую тесную взаимосвязь, клетки кумулюса и ооцит имеют различные мембранные потенциалы. Клетки кумулюса передают ооциту специфические электрические сигналы, которые либо поддерживают мейотический арест, либо снимают блок мейоза. В первом случае мембрана ооцита поддерживается в гиперполяризованном состоянии. В другом случае наступает деполяризация мембраны ооцита в результате действия гонадотропинов на кумулюсные клетки. Деполяризация клеток гранулезы, таким образом, является важным событием созревания фолликула. Эта гипотеза подтверждается тем, что пролиферация клеток гранулезы,
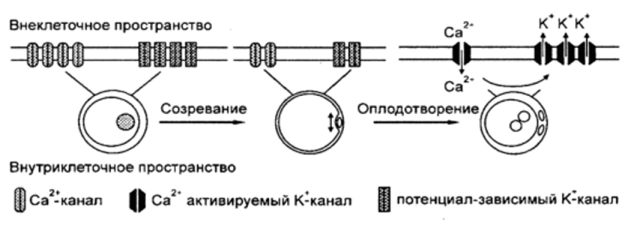
Динамика активности ионных каналов в ооцитах млекопитающих (по ТозН Е., Вот Я. 2004)
синтез ими стероидов и апоптоз могут изменяться под действием избирательных антагонистов К + -каналов. Такая тесная связь между ооцитом и клетками гранулезы уменьшается в конце созревания благодаря разрушению плотных контактов и увеличению объема кумулюсных масс. Искусственное удаление кумулюса и выделение незрелых ооцитов также приводит к изменению мембранного потенциала.
Эксперименты на ооцитах крупного рогатого скота продемонстрировали, что в сложных метаболических преобразованиях ооцитов в ходе их созревания важную роль играет изменение внутриклеточной концентрации ионов кальция. Активность Са 2+ -каналов Ь-типа, локализованных в плазматической мембране ооцитов, уменьшается в ходе завершения мейотического деления.
Имеющаяся на сегодня картина динамики активности ионных каналов в мембране ооцитов млекопитающих представлена на рисунке 12. На стадии зародышевого пузырька наибольшей активностью обладают К 4 -каналы и Са 2+ -кана- лы Ь-типа. Их активность заметно снижается к метафазе второго мейотического деления.
ПРЕЖДЕВРЕМЕННОЕ ИСТОЩЕНИЕ ЯИЧНИКОВ
На приёме репродуктолога пациенты часто задают вопрос: «Каковы риски развития преждевременного истощения яичников после проведения программы искусственного оплодотворения?». Врач почти в половине случаев сталкивается с проблемой страхов и переживаний пациентов, убеждённых в том, что гормональная стимуляция сократит количество существующих фолликулов в яичниках и неминуемо приведёт к ранней менопаузе. В связи с этим, второй вопрос, наиболее часто обсуждаемый на приёме, является программа ЭКО в спонтанном цикле без гормональной стимуляции.
Для ответа на эти вопросы следует напомнить особенности формирования фолликулов в яичниках. К моменту рождения девочки в яичниках находится около двух миллионов яйцеклеток, это известный факт. К моменту наступления пубертатного периода в яичниках остаётся всего лишь 200000-500000 клеток. С возрастом количество фолликулов уменьшается. Каждый месяц в среднем формируется около 30 фолликулов, потенциально способных преобразоваться в преовуляторные. Однако, физиология женщины так устроена, что в середине фолликулярной фазы цикла снижается уровень гормона ФСГ и, в связи с этим, дальнейшее развитие продолжает только один доминантный фолликул, а остальные подвергаются обратному развитию (этот процесс называется атрезией).
Этот процесс происходит каждый менструальный цикл — закладывается рост нескольких фолликулов, а овуляция происходит только одного фолликула. Это эволюционно сложившийся механизм и очень важен для понимания, что происходит в стимулированном цикле ВРТ.
В момент стимуляции постоянным количеством гормона ФСГ растут все примордиальные фолликулы, которые природа сформировала в данном менструальном цикле, и по сути мы «спасаем» от неминуемой гибели фолликулы, которым суждено погибнуть в данном цикле. Именно по этой причине легко можно объяснить, почему раньше времени не наступит истощение яичников после программ экстракорпорального оплодотворения.
Второй вопрос о программе ЭКО в спонтанном цикле без применения гормонов. Это низкоэффективные программы по многому ряду причин. Растёт один доминантный фолликул и вероятность получения зрелой качественной яйцеклетки мала. Либо происходит овуляция, и врач не успевает сделать пункцию фолликула, либо фолликул оказывается пустой без яйцеклетки, либо клетка не зрелая и не оплодотворяется, либо эмбриологам удаётся получить эмбрион, но беременность не наступает.
Подобные программы ВРТ имеет смысл проводить у пациенток, имеющих строгие противопоказания к применению гормонов (тяжёлые экстрагенитальные и онкологические заболевания, тромбозы в анамнезе и др.). В целом дороговизна ЭКО в спонтанном цикле не соответствует целесообразности процедуры, поэтому мы стараемся отказывать пациентам в проведении низкоэффективных программ.
В связи с этим возникает ещё один закономерный вопрос у пациенток. Экстракорпоральное оплодотворение с применением минимальной стимуляции яичников. В данных программах используются малые дозы ФСГ, на фоне которых созревают 3-4 фолликула. Такие стимуляции являются самыми актуальными во всех зарубежных клиниках, особенно в странах, где запрещено переносить более одного эмбриона. Данные программы ЭКО являются профилактикой тяжёлых угрожающих жизни осложнений, таких как синдром гиперстимуляции яичников, тромбозы, а также многоплодной беременности, тяжёлого течения беременности и рисков рождения недоношенных детей.
Однако необходимо иметь чёткое представление о том, какой когорте пациенток показана минимальная стимуляция. Если на приём приходит молодая женщина до 35 лет с хорошим фолликулярным запасом, нормальными гормонами крови, то ,конечно, целесообразно провести минимальную стимуляцию с получением малого количества ооцитов и минимальным риском осложнений.
Но, к сожалению, контингент возрастных женщин, готовящихся к программе ЭКО, растёт с каждым годом. На приём приходят всё чаще женщины старшей возрастной группы после 40 лет, с гормональными нарушениями, с низким фолликулярным ресурсом, после оперативных вмешательств на яичниках, после множества неудачных попыток ЭКО в других клиниках с получением 1-2-х ооцитов. Конечно, таким женщинам абсолютно не показаны мягкие схемы стимуляции, т.к. возможен бедный ответ яичников либо клеток может быть не получено вовсе.
В заключении следует добавить, что механизм преждевременного истощения яичников довольно сложный, генетически детерминированный, т.е. чаще носит наследственный характер. Причины возникновения разные, до конца не изучены. Могут быть и психоэмоциональные причины, связанные с серьёзным нервным потрясением. Могут быть аутоиммунные процессы в организме, негативно влияющие на ткань яичника. Ятрогенный характер истощения яичников — один из самых частых факторов, когда происходит потеря фолликулярного запаса яичников во время операций либо в случае химио- и лучевой терапии при злокачественных опухолях.
Но программы экстракорпорального оплодотворения даже с использованием высоких доз гормонов не вызывают синдрома преждевременного истощения яичников. И этот факт является камнем преткновения не только для пациентов, но врачей. Пациенты много лет откладывают проведение ВРТ ввиду страха перед гормональной стимуляцией. Однако, истощение яичников возникает совсем по другим причинам.
Читайте также: