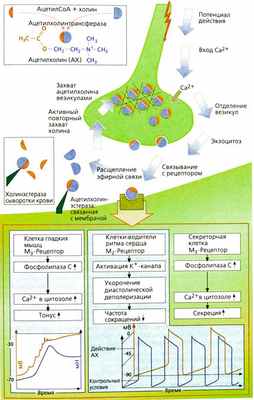
Функции ацетилхолина в холинергическом синапсе
Добавил пользователь Валентин П. Обновлено: 27.01.2026
Источник: «Наглядная фармакология».
Автор: X. Люльман. Пер. с нем. Изд.: М.: Мир, 2008 г.
Ацетилхолин (АХ) — медиатор в постганглионарных синапсах — накапливается в высокой концентрации в везикулах аксоплазмы нервного окончания. АХ образуется из холина и активированной уксусной кислоты (ацетилкофермент А) под действием фермента ацетилхолинтрансферазы. Высокополярный холин активно захватывается аксоплазмой. На мембране холинергического аксона и нервных окончаний имеется специальная транспортная система. Механизм высвобождения медиатора до конца не раскрыт. Везикулы закреплены в цитоскелете при помощи белка синапсина таким образом, что их концентрация около пресинаптической мембраны высокая, однако контакт с мембраной отсутствует. При возникновении возбуждения повышается концентрация Са2+ в аксоплазме, активируются протеинкиназы, и происходит фосфорилирование синапсина, приводящее к отсоединению везикул и связыванию их с пресинаптической мембраной. Затем содержимое везикул выбрасывается в синаптическую щель. Ацетилхолин мгновенно проходит сквозь синаптическую щель (молекула АХ имеет длину около 0,5 нм, а ширина щели составляет 30-40 нм). На постсинаптической мембране, т. е. мембране целевого органа, АХ взаимодействует с рецепторами. Эти рецепторы возбуждаются также алкалоидом мускарином и поэтому называются мускариновыми ацетилхолиновыми рецепторами (М-холинорецепторы). Никотин имитирует действие ацетилхолина на рецепторы ганглионарных синапсов и концевой пластинки. Никотин возбуждает холинорецепторы ганглионарных синапсов и концевой пластинки мотонейрона, поэтому этот тип рецепторов назван никотиновыми ацетилхолиновыми рецепторами (N-холинорецепторы).
В синаптической щели ацетилхолин быстро инактивируется специфической ацетилхолинэстеразой, находящейся в щели, а также менее специфической сывороточной холинэстеразой (бутирилхолинэстеразой), находящейся в сыворотке крови и интерстициальной жидкости.
По своему строению, способу передачи сигнала и сродству к различным лигандам М-холинорецепторы подразделяются на несколько типов. Рассмотрим M1, М2- и М3-рецепторы. M1-Рецепторы находятся на нервных клетках, например ганглиях, и их активация способствует переходу возбуждения с первого на второй нейрон. М2-Рецепторы расположены в сердце: открытие калиевых каналов приводит к замедлению диастолической деполяризации и уменьшению частоты сердечных сокращений. М3-Рецепторы играют роль в поддержании тонуса гладких мышц, например, кишечника и бронхов. Возбуждение этих рецепторов приводит к активации фосфолипазы С, деполяризации мембраны и повышению тонуса мышц. М3-Рецепторы расположены также в клетках желез, которые активируются посредством фосфолипазы С. В головном мозге имеются разные типы М-холинорецепторов, играющие роль во многих функциях: передаче возбуждения, памяти, обучаемости, болевой чувствительности, контроле активности ствола мозга. Активация М3-рецепторов в эндотелии сосудов может приводить к высвобождению оксида азота N0 и таким образом расширять сосуды.
Источник:
Клиническая фармакология по Гудману и Гилману том 1.
Редактор: профессор А.Г. Гилман Изд.: Практика, 2006 год.
Ацетилхолин (лат. Acetylcholinum) — медиатор нервной системы, биогенный амин, относящийся к веществам, образующимся в организме.
Ацетилхолину принадлежит важная роль как медиатору центральной нервной системы. Он участвует в передаче импульсов в разных отделах мозга, при этом малые концентрации облегчают, а большие — тормозят синаптическую передачу. Изменения в обмене ацетилхолина могут привести к нарушению функций мозга.
Ацетилхолин является посредником передачи нервного импульса к мышце. При недостатке ацетилхолина снижается сила сокращений мышц.
Окончания нервных волокон, для которых он служит медиатором, называются холинергическими, а рецепторы, взаимодействующие с ним, называют холинорецепторами. Холинорецепторы постганглионарных холинергических нервов (сердца, гладких мышц, желез) обозначают как м-холинорецепторы (мускариночувствительные), а расположенные в области ганглионарных синапсов и в соматических нервномышечных синапсах — как н-холинорецепторы (никотиночувствительнные). Такое деление связано с особенностями реакций, возникающих при взаимодействии ацетилхолина с этими биохимическими системами: мускариноподобных в первом случае и никотиноподобных — во втором; м- и н-холинорецепторы находятся также в разных отделах ЦНС.
При микроэлектродной регистрации электрических потенциалов постсинаптической мембраны нервно-мышечного синапса Фетт и Катц (Fatt and Katz, 1952) выявили спонтанные небольшие (0,1—3 мВ) деполяризующие потенциалы, возникающие случайным образом примерно 1 раз в секунду. Авторы назвали эти потенциалы миниатюрными потенциалами концевой пластинки. Их амплитуда была существенно ниже пороговой для развития потенциала действия. Они увеличивались под действием ингибитора АХЭ неостигмина и блокировались тубокурарином (конкурентным блокатором N-холинорецепторов); следовательно, они были обусловлены выделением ацетилхолина. В связи с этим было высказано предположение, что ацетилхолин выделяется из пресинаптических окончаний дробными постоянными порциями — квантами. Вскоре был обнаружен и морфологический субстрат квантов — синаптические пузырьки (De Robertis and Bennett, 1955). Когда в окончание аксона мотонейрона приходит потенциал действия, выделяется 100 и более квантов (пузырьков) ацетилхолина (Katz and Miledi, 1965). Закономерности хранения и выделения ацетилхолина, изученные на нервно-мышечном синапсе, применимы и к другим холинергическим синапсам с быстрой передачей.
Предполагается, что в каждом пузырьке содержится от 1000 до 50 000 молекул ацетилхолина, а в пресинаптическом окончании мотонейрона содержится 300 000 и более пузырьков. Кроме того, не исключено, что достаточно существенное количество ацетилхолина диффузно растворено в аксоплазме. Запись токов одиночных каналов постсинаптической мембраны нервно-мышечного синапса при постоянной аппликации ацетилхолина показала, что одна молекула этого медиатора вызывает потенциал порядка 3 х 10”7 В. Из этого следует, что даже минимальное (по расчетам) количество ацетилхолина в одном пузырьке — 1000 молекул — достаточно для того, чтобы вызвать миниатюрный потенциал концевой пластинки (Katz and Miledi, 1972).
Экзоцитоз ацетилхолина и других медиаторов из пресинаптических окончаний подавляется ботулотоксином и столбнячным токсином — ядами Clostridium botulinum и Clostridium tetani соответственно. Этими анаэробными спорообразующими организмами вырабатываются одни из самых сильных из известных токсинов (Shapiro et а. 1998). Токсины Clostridium, состоящие из связанных дисульфидными мостиками тяжелой и легкой цепей, соединяются с неизвестным пока рецептором на холинергическом окончании и затем посредством эндоцитоза переносятся в цитозоль. Легкая цепь представляет собой цинксодержащую эндопептидазу, которая после активации гидролизует компоненты ядра комплекса SNARE, участвующего в экзоцитозе. Различные типы ботулотоксина разрушают разные белки пресинаптической мембраны (синтаксин-1 и SNAP-25) и синаптических пузырьков (синаптобревин). Ботулотоксин А как лекарственное средство рассматривается в гл. 9 и 66.
Столбнячный токсин — это яд центрального действия: он ретроградно переносится по аксонам мотонейронов в тела этих нейронов в спинном мозге, далее переходит в связанные с мотонейронами тормозные нейроны и блокирует экзоцитоз медиатора из последних. Именно это и приводит к характерным для столбняка судорогам. Яд паука черная вдова — а-латротоксин — связывается с трансмембранными белками пресинаптических окончаний нейрексинами, вызывая массивный экзоцитоз синаптических пузырьков (Schiavo et al., 2000).
Функции ацетилхолина в холинергическом синапсе
Ткани и органы. Нервная ткань
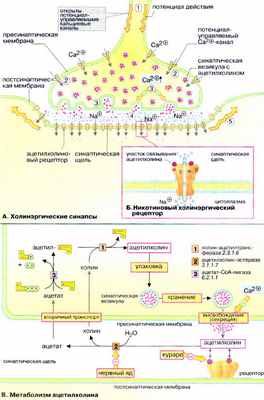
А. Холинэргические синапсы
Передача сигналов между нейронами и от нейронов к мышечным клеткам (так называемая нейронейрональная и нейромышечная трансдукция) происходит в нервных окончаниях (синапсах). С помощью сигнальных веществ, медиаторов. Синапсы образованы мембранами двух контактирующих клеток, пресинаптической и постсинаптической которые разделены узкой синаптической щелью. Медиатор выделяется в синаптическую щель за счет зкзоцитоза, диффундирует к рецепторам постсинаптической мембраны, связывается с ними и передает сигнал соседней клетке. Белки-рецепторы — это лиганд-активируемые ионные каналы (см. рис. 341) либо мембранные белки, которые управляют ионными каналами посредством G-белков (см. рис. 373).
Ацетилхолин — нейромедиатор моторной концевой пластинки. Ацетилхолиновые рецепторы (никотиновый и мускариновый) — это лиганд-активируемые ионные каналы, которые открываются для прохождения ионов Na + и К + . Никотиновые рецепторы (быстрые) локализованы главным образом в месте контакта аксонов со скелетными мышцами. Мускариновые рецепторы (медленные) локализованы в головном мозге, секреторных клетках, гладких и сердечных мышцах.
Процесс передачи сигнала включает следующие этапы. Потенциал действия достигает пресинаптической мембраны ( 1 ). Это вызывает открывание потенциал-управля емых Сa 2+ -каналов ( 2 ). Ионы Са 2+ проникают из внеклеточного пространства в клетку, их уровень в синапсе резко увеличивается, что инициирует процесс экзоцитоза. Синаптические везикулы выделяют содержимое (ацетилхолин) в синаптическую щель ( 3 ). Молекулы ацетилхолина диффундируют через синаптическую щель, связываются с постсинаптическими рецепторами и активируют их ( 4 ). Поток ионов Na + изменяет потенциал покоя постсинаптической мембраны нервной или мышечной клетки настолько, что открываются соседние потенциал-управляемые Na + каналы и возникает потенциал действия ( 5 , см. рис. 341).
Б. Никотиновый холинэргический рецептор
Наиболее детально изучен рецептор ацетилхолина, активируемый никотином. Это трансмембранный комплекс из пяти субъединиц (α 2 βδγ, 250-270 кДа), образующий лиганд-активируемый (хемовозбудимый) ионный канал, проницаемый для ионов Na + и К + . Участки связывания ацетилхолина локализованы на внеклеточной части α-субъединиц. При связывании лиганда в центре молекулы формируется трансмембранный канал, входное отверстие которого имеет форму воронки диаметром около 2 нм. Предполагается, что в формировании канала принимают участие все пять субъединиц. Канал открывается на короткое время для прохождения ионов Na + и К + . Считается, что открывание и закрывание канала происходит в результате аллостерических изменений в заряженных участках полипептидных цепей молекулы рецептора.
Рецептор может связывать различные лекарственные вещества: например, никотин действует как агонист ацетилхолина.
В. Метаболизм ацетилхолина
Ацетилхолин , уксуснокислый эфир холина, образуется в цитоплазме аксонов из ацетил-КоА и холина [ 1 ]. Нейромедиатор хранится в синаптических везикулах , в каждой везикуле содержится примерно 1000-10000 молекул ацетилхолина. После выделения из везикул ацетилхолин попадает в синаптическую щель. Избыток ацетилхолина расщепляется ацетилхолин-эстеразой [ 2 ]. Этот фермент имеет высокое число оборотов, что гарантирует быстрое удаление сигнального вещества. Продукты гидролиза, холин и уксусная кислота , активно захватываются пресинаптической частью синапса и используются для повторного синтеза ацетилхолина [ 3 ].
Соединения, блокирующие остаток серина в активном центре ацетилхолин-эстеразы [ 2 ], например токсин Е605, пролонгируют действие ацетилхолина и действуют как нейротоксины. Напротив, D-тубокурарин ( яд кураре, которым индейцы пропитывали наконечники стрел) является конкурентным ингибитором ацетилхолина при связывании с рецептором.
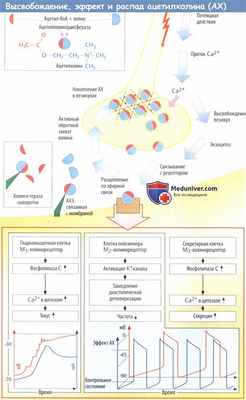
Функции ацетилхолина в холинергическом синапсе
Функции ацетилхолина в холинергическом синапсе
Ацетилхолин (АХ) служит медиатором постганглионарных синапсов окончаний парасимпатических нервов. Ацетилхолин (АХ) в высокой концентрации содержится в синаптических пузырьках (везикулах), плотно расположенных в аксоплазме пресинаптических окончаний. АХ образуется из холина и активированного ацетата (ацетилкоэнзима А); эту реакцию катализирует цитоплазматический фермент ацетилхолинтрансфераза.
Перенос высокополярного холина в аксоплазму осуществляет специфический холиновый переносчик, расположенный в мембранах окончаний холинергических аксонов и субпопуляции везикул. Во время непрерывной или интенсивной стимуляции этот переносчик обеспечивает стабильный синтез и высвобождение АХ. Образовавшийся АХ транспортируется в пузырьки везикулярным транспортером (переносчиком) АХ. Механизм высвобождения АХ до конца не изучен.
Заполненные ацетилхолина (АХ) везикулы прикрепляются к цитоскелету белком синапсином. Это обеспечивает группирование везикул вблизи пресинаптической мембраны, одновременно препятствуя их слиянию друг с другом. Предполагается, что во время активации мембраны нервного волокна ионы Са 2+ попадают в аксоплазму через потенциалзависимые каналы и активируют протеинкиназы, фосфорилирующие синапсин.
В итоге расположенные рядом с мембраной везикулы отделяются от цитоскелета и сливаются с пресинаптической мембраной. В результате слияния содержимое везикул попадает в синаптическую щель; одновременно с этим специфический холиновый переносчик встраивается в плазматическую мембрану. АХ быстро диффундирует через синаптическую щель (молекула АХ чуть больше 0,5 нм; размер синаптической щели составляет 20-30 нм). На постсинаптической мембране эффекторной клетки АХ связывается со своими рецепторами.
Эти рецепторы также активируются алкалоидом мускарином, поэтому их называют мускариновыми (М) холинорецепторами. Действие ацетилхолина (АХ) на ганглионарные рецепторы и рецепторы на двигательной концевой пластинке можно воспроизвести никотином; эти рецепторы называются никотиновыми (N)холинорецепторами
Выделившийся ацетилхолин (АХ) быстро подвергается гидролизу и инактивируется специфической ацетилхолинэстеразой (АХЭ), расположенной в пре- и постсинаптических мембранах (базальной пластинке двигательных концевых пластин), либо неспецифической сывороточной холинэстеразой (бутирилхолинэстеразой) — водорастворимым ферментом, который находится в сыворотке и интерстициальной жидкости.
В зависимости от молекулярной структуры, особенностей передачи сигнала и сродства к лигандам М-холинорецепторы можно разделить на пять подтипов. Здесь мы рассмотрим наиболее изученные подтипы М1, М3 и М3. М,-холинорецепторы расположены в нервных клетках, и прежде всего в клетках головного мозга. М2-холинорецепторы передают эффекты АХ на сердце: открытие К + -каналов замедляет диастолическую деполяризацию в клетках синусно-предсердного узла, уменьшая ЧСС.
М3-холинорецепторы отвечаютза регуляциютонуса гладкой мускулатуры. Например, в кишечнике и бронхахактивация этих рецепторов стимулирует фосфолипазу С, деполяризацию мембраны и повышает мышечный тонус. М3-холинорецепторы также находятся в железистом эпителии, в котором происходит активация фосфолипазы С и секреторной активности. В ЦНС представлены все подтипы М-холинорецепторов, где они отвечаютза различные процессы: от регуляции активности коры, процессов памяти и обучения, обработки болевой импульсации до двигательного контроля на уровне ствола мозга.
Ацетилхолин (АХ) расслабляет тонус мышц кровеносных сосудов. Действие это непрямое, оно обусловлено стимуляцией М3-холинорецепторов эндотелиальных клеток, которые в ответ высвобождают NO (оксид азота — эндотелиальный сосудорасширяющий фактор). Оксид азота диффундирует в гладкомышечные клетки, где вызывает расслабление тонуса, обусловленного их сокращением.
Учебное видео - строение синапса
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
ФУНКЦИИ ХОЛИНЕРГИЧЕСКИХ СИНАПСОВ
Холинергические синапсы локализованы в ЦНС (ацетилхолин регулирует моторику, пробуждение, память, обучение), а также в вегетативных ганглиях, мозговом слое надпочечников, каротидных клубочках, скелетных мышцах и внутренних органах, получающих постганглионарные парасимпатические волокна.
В скелетных мышцах синапсы занимают небольшую часть мембраны и изолированы друг от друга. В верхнем шейном ганглии около 100000 нейронов упакованы в объеме 2 — 3 мм 3 .
Ацетилхолин синтезируется в аксоплазме холинергических окончаний из ацетилкоэнзима А (митохондриального происхождения) и незаменимого аминоспирта холина при участии фермента холин-ацетилтрансферазы (холинацетилаза). Иммуноцитохимический метод определения этого фермента позволяет установить локализацию холинергических нейронов.
Ацетилхолин депонируется в синаптических пузырьках (везикулах) в связи с АТФ и нейропептидами (вазоактивный интестинальный пептид, нейропептид Y). Квантами выделяется при деполяризации пресинаптической мембраны и возбуждает холинорецепторы. В окончании двигательного нерва находится около 300 000 синаптических пузырьков, в каждом из них депонировано от 1000 до 50000 молекул ацетилхолина.
Весь ацетилхолин, находящийся в синаптической щели, подвергается гидролизу ферментом ацетилхолинэстеразой (истинная холинэстераза) с образованием холина и уксусной кислоты. Одна молекула медиатора инактивируется в течение 1 мс. Ацетилхолинэстераза локализована в аксонах, дендритах, перикарионе, на пресинаптической и постсинаптической мембранах.
Холин в 1000 — 10 000 раз менее активен по сравнению с ацетилхолином; 50 % его молекул подвергается нейрональному захвату и вновь участвует в синтезе ацетилхолина. Уксусная кислота окисляется в цикле трикарбоновых кислот.
Псевдохолинэстераза (бутирилхолинэстераза) крови, печени, нейроглии катализирует гидролиз эфиров растительного происхождения и лекарственных средств.
Холинорецепторы
Холинорецепторы представляют собой гликопротеины, состоящие из нескольких субъединиц. Большинство холинорецепторов являются резервными. На постсинаптической мембране в нервно-мышечном синапсе расположено до 100 млн холинорецепторов, из них не функционируют 40 — 99 %. В холинергическом синапсе на гладкой мышце находятся около 1,8 млн холинорецепторов, резервными являются 90 — 99%.
В 1914г. Генри Дейл установил, что эфиры холина могут оказывать как мускариноподобный, так и никотиноноподобный эффекты. В соответствии с химической чувствительностью холинорецепторы классифицируют на мускариночувствительные (М) и никотиночувствительные (Н) (табл. 20). Ацетилхолин имеет гибкую молекулу, способную в различных стереоконформациях возбуждать М- и Н-холинорецепторы.
М-холинорецепторы возбуждаются ядом мухомора мускарином и блокируются атропином. Они локализованы в нервной системе и внутренних органах, получающих парасимпатическую иннервацию (вызывают угнетение сердца, сокращение гладких мышц, повышают секреторную функцию экзокринных желез) (табл. 15 в лекции 9). М-холинорецепторы ассоциированы с G-белками и имеют 7 сегментов, пересекающих, как серпантин, клеточную мембрану.
Молекулярное клонирование позволило выделить пять типов М-холинорецепторов:
1. М1-холинорецепторы ЦНС (лимбическая система, базальные ганглии, ретикулярная формация) и вегетативных ганглиев;
2. М2-холинорецепторы сердца (снижают частоту сердечных сокращений, атриовентрикулярную проводимость и потребность миокарда в кислороде, ослабляют сокращения предсердий);
3. М3-холинорецепторы:
· гладких мышц (вызывают сужение зрачков, спазм аккомодации, бронхоспазм, спазм желчевыводящих путей, мочеточников, сокращение мочевого пузыря, матки, усиливают перистальтику кишечника, расслабляют сфинктеры);
· желез (вызывают слезотечение, потоотделение, обильное отделение жидкой, бедной белком слюны, бронхорею, секрецию кислого желудочного сока).
Таблица 20.Холинорецепторы
| Рецепторы | Агонисты | Антагонисты | Локализация | Функции | Эффекторный механизм |
| Мускариночувствительные | |||||
| м1 | Оксотреморин | Пиренцепин | ЦНС | Контроль психических и моторных функций, реакции пробуждения и обучения | Активация фосфолипазы С посредством Gq/11-белка |
| Вегетативные ганглии | Деполяризация (поздний постсинаптический потенциал) | ||||
| M2 | Метоктрамин | Сердце: синусный узел | Замедление спонтанной деполяризации, гиперполяризация | Ингибирование аденилатциклазы посредством Gi-белка, активация К + -каналов | |
| предсердия | Укорочение потенциала действия, уменьшение сократимости | ||||
| атриовентрикулярный узел | Уменьшение проводимости | ||||
| желудочки | Незначительное уменьшение сократимости | ||||
| М3 | Гексагидросила дифенидол | Гладкие мышцы | Сокращение | Аналогичен М1 | |
| Экзокринные железы | Повышение секреторной функции | ||||
| М4 | Тропикамид Химбацин | Альвеолы легких | — | Аналогичен М2 | |
| М5 | — | — | ЦНС (черная субстанция среднего мозга, гиппокамп) | — | Аналогичен М1 |
| Никотиночувствительные | |||||
| нH | Диметилфенил пиперазин Цитизин Эпибатидин | Арфонад | ЦНС | Аналогичны функциям М, | Открытие каналов для Na + , K + , Са 2+ |
| Вегетативные ганглии | Деполяризация и возбуждение постганглионарных нейронов | ||||
| Мозговой слой надпочечников | Секреция адреналина и норадреналина | ||||
| Каротидные клубочки | Рефлекторное тонизирование дыхательного центра | ||||
| Нм | Фенилтримети ламмоний | Тубокурарин-хлорид a-Бунгаротоксин | Скелетные мышцы | Деполяризация концевой пластинки, сокращение |
Внесинаптические М3-холинорецепторы находятся в эндотелии сосудов и регулируют образование сосудорасширяющего фактора — окиси азота (NО).
4. М4 - и М5-холинорецепторы имеют меньшее функциональное значение.
М1-, М3- и М5-холинорецепторы, активируя посредством Gq/11-белка фосфолипазу С клеточной мембраны, увеличивают синтез вторичных мессенджеров — диацилглицерола и инозитолтрифосфата. Диацилглицерол активирует протеинкиназу С, инозитолтрифосфат освобождает ионы кальция из эндоплазматического ретикулума,
М2- и М4-холинорецепторы при участии Gi- и G0-белков ингибируют аденилатциклазу (тормозят синтез цАМФ), блокируют кальциевые каналы, а также повышают проводимость калиевых каналов синусного узла.
Дополнительные эффекты М-холинорецепторов — мобилизация арахидоновой кислоты и активация гуанилатциклазы.
Н-холинорецепторы возбуждаются алкалоидом табака никотином в малых дозах, блокируются никотином в больших дозах.
Биохимическая идентификация и выделение Н-холинорецепторов стали возможны благодаря открытию их избирательного высокомолекулярного лиганда a-бунгаротоксина — яда тайваньской гадюки Bungarus multicintus и кобры Naja naja. Н-холинорецепторы находятся в ионных каналах, в течение миллисекунд они повышают проницаемость каналов для Na + , K + и Са 2+ (через один канал мембраны скелетной мышцы проходит 5 — 10 7 ионов натрия за 1 с).
Таблица 21.Классификация лекарственных средств, влияющих на холинерги-ческие синапсы (указаны основные препараты)
| Холиномиметики | |
| М, Н-холиномиметики | ацетилхолин-хлорид, карбахолин |
| М-холиномиметики | пилокарпин, ацеклидин |
| Н-холиномиметики (ганглиостимуляторы) | цитизин, лобелин |
| Средства, повышающие выделение ацетилхолина | |
| цисаприд | |
| Антихолинэстеразные средства | |
| Обратимые блокаторы | физостигмин, галантамин, амиридин, прозерин |
| Необратимые блокаторы | армин |
| Холиноблокаторы | |
| М-холиноблокаторы | атропин, скополамин, платифиллин, метацин, пиренцепин, ипратропия бромид |
| Н-холиноблокаторы (ганглиоблокаторы) | бензогексоний, пентамин, гигроний, арфонад, пахикарпин, пирилен |
| Миорелаксанты | |
| Антидеполяризующие | тубокурарин-хлорид, пипекурония бромид, атракурия бесилат, мелликтин |
| Деполяризующие | дитилин |
Н-холинорецепторы широко представлены в организме. Их классифицируют на Н-холинорецепторы нейронального (Нн) и мышечного (Нм) типов.
Нейрональные Нн-холинорецепторы представляют собой пентамеры и состоят из субъединиц a2 — a9, и β2 — β4(четыре трансмембранные петли). Локализация нейрональных Н-холинорецепторов следующая:
· кора больших полушарий, продолговатый мозг, клетки Реншоу спинного мозга, нейрогипофиз (повышают секрецию вазопрессина);
· вегетативные ганглии (участвуют в проведении импульсов с преганглионарных волокон на постганглионарные);
· мозговой слой надпочечников (повышают секрецию адреналина и норадреналина);
· каротидные клубочки (участвуют в рефлекторном тонизировании дыхательного центра).
Мышечные Нм-холинорецепторы вызывают сокращение скелетных мышц. Они представляют собой смесь мономера и димера. Мономер состоит из пяти субъединиц (a1 — a2, β, γ, ε, δ), окружающих ионные каналы. Для открытия ионных каналов необходимо связывание ацетилхолина с двумя a-субъединицами.
Пресинаптические М-холинорецепторы тормозят, пресинаптические Н-холинорецепторы стимулируют высвобождение ацетилхолина.
Холинергические синапсы: строение, функции
Холинергические синапсы представляют собой место, в котором происходит контакт двух нейронов или нейрона и эффекторной клетки, получающей сигнал. Синапс состоит из двух мембран – пресинаптической и постсинаптической, а также из синаптической щели. Передача нервного импульса осуществляется посредством медиатора, то есть вещества-передатчика. Происходит это в результате взаимодействия рецептора и медиатора на постсинаптической мембране. В этом заключаются основные функции холинергического синапса.
Медиатор и рецепторы
В парасимпатической НС медиатором является ацетилхолин, рецепторами – холинорецепторы двух типов: Н (никотин) и М (мускарин). М-холиномиметики, обладающие прямым типом действия, могут стимулировать рецепторы на мембране постсинаптического типа.
Синтез ацетилхолина осуществляется в цитоплазме нейронных холинергических окончаний. Он образуется из холина, а также ацетилкоэнзима-А, который имеет митохондриальное происхождение. Синтез происходит под действием цитоплазматического энзима холинацетилазы. В синаптических пузырьках происходит депонирование ацетилхолина. В каждом из таких пузырьков может находиться до нескольких тысяч ацетилхолиновых молекул. Нервный импульс провоцирует высвобождение молекул ацетилхолина в синаптическую щель. После этого он вступает во взаимодействие с холинорецепторами. Строение холинергического синапса уникально.
Строение
По данным, которые имеются у биохимиков, холинорецептор нервно-мышечного синапса может включать 5 белковых субъединиц, которые окружают ионный канал и проходят сквозь всю толщу мембраны, состоящей из липидов. Пара молекул ацетилхолина вступает во взаимодействие с парой α-субъединиц. Это приводит к тому, что открывается ионный канал и постсинаптическая мембрана деполяризуется.
Виды холинергических синапсов
Холинорецепторы по-разному локализованы и так же по-разному чувствительны к воздействию фармакологических веществ. В соответствии с этим различают:
- Маскариночувствительные холинорецепторы – так называемые М-холинорецепторы. Мускарин представляет собой алкалоид, присущий ряду ядовитых грибов, к примеру мухоморам.
- Никотиночувствительные холинорецепторы – так называемые Н-холинорецепторы. Никотин представляет собой алкалоид, содержащийся в листьях табака.
Их расположение
Первые располагаются в постсинаптической мембране клеток в составе эффекторных органов. Расположены они у окончаний постганглионарных парасимпатических волокон. Помимо этого они также есть в нейронных клетках вегетативных ганглиев и в коре головного мозга. Установлено, что М-холинорецепторы различной локализации гетерогенны, что обуславливает различную чувствительность холинергических синапсов к веществам фармакологической природы.

Виды в зависимости от расположения
Биохимики различают несколько видов М-холинорецепторов:
- Расположенные в вегетативных ганглиях и в ЦНС. Особенностью первых является то, что они локализованы вне синапсов – М1-холинорецепторы.
- Расположенные в сердце. Некоторые из них способствуют снижению высвобождения ацетилхолина – М2-холинорецепторы.
- Расположенные в гладких мышцах и в большей части эндокринных желез – М3-холинорецепторы.
- Расположенные в сердце, в стенках легочных альвеол, в ЦНС – М4-холинорецепторы.
- Расположенные в ЦНС, в радужной оболочке глаза, в слюнных железах, в мононуклеарных кровяных клетках – М5-холинорецепторы.
Воздействие на холинорецепторы
Большая часть эффектов, оказываемых известными фармакологическими веществами, влияющими на М-холинорецепторы, связана с взаимодействием этих веществ и постсинаптических М2- и М3-холинорецепторов.
Рассмотрим классификацию средств, стимулирующих холинергические синапсы, ниже.
Н-холинорецепторы располагаются в постсинаптической мембране нейронов ганглиев у окончаний каждого из преганглионарных волокон (в парасимпатических и симпатических ганглиях), в синокаротидной зоне, в мозговом слое надпочечников, в нейрогипофизе, в клетках Реншоу, в скелетных мышцах. Чувствительность различных Н-холинорецепторов неодинакова к веществам. Например, Н-холинорецепторы в структуре вегетативных ганглиев (рецепторы нейтрального типа) имеют значительные отличия от Н-холинорецепторов в скелетных мышцах (рецепторы мышечного типа). Именно такая их особенность позволяет избирательно блокировать ганглии специальными веществами. Например, курареподные вещества способны блокировать нервно-мышечную передачу.
Пресинаптические холинорецепторы и адренорецепторы участвуют в регуляции процесса высвобождения ацетилхолина в синапсах нейроэффекторной природы. Возбуждение этих рецепторов будет угнетать высвобождение ацетилхолина.
Ацетилхолин взаимодействует с Н-холинорецепторами и изменяет их конформацию, повышает уровень проницаемости постсинаптической мембраны. Ацетилхолин оказывает возбуждающий эффект на ионы натрия, которые проникают затем внутрь клетки, а это приводит к тому, что постсинаптическая мембрана деполяризуется. Изначально возникает локальный синаптический потенциал, который достигает определенной величины и начинает процесс генерации потенциала действия. После этого местное возбуждение, которое ограничено синаптической областью, начинает распространяться по всей клеточной мембране. Если происходит стимуляция М-холинорецептора, то при передаче сигнала значительную роль играют вторичные мессенджеры и G-белки.
Ацетилхолин действует в течение весьма короткого времени. Это обусловлено тем, что он стремительно гидролизуется под действием фермента ацетилхолинэстеразы. Холин, который образуется в процессе гидролиза ацетилхолина, в половине объема будет захвачен пресинаптическими окончаниями и транспортирован в цитоплазму клетки для последующего биосинтеза ацетилхолина.
Вещества, которые воздействуют на холинергические синапсы
Фармакологические и разнообразные химические вещества способны воздействовать на множество процессов, которые связаны с синаптической передачей:
- Процесс синтеза ацетилхолина.
- Процесс высвобождения медиатора. К примеру, карбахолин способен усиливать процесс выделения ацетилхолина, а ботулиновый токсин может препятствовать процессу высвобождения медиатора.
- Процесс взаимодействия между ацетилхолином и холинорецептором.
- Гидролиз ацетилхолина энзиматической природы.
- Процесс захвата холина, образованного в результате гидролиза ацетилхолина, пресинаптическими окончаниями. К примеру, гемихолиний способен угнетать нейроновый захват и транспортировку холина в цитоплазму клетки.
Классификация
Средства, стимулирующие холинергические синапсы, способны оказывать не только этот эффект, но и холиноблокирующий (угнетающий) эффект. В качестве основы для классификации подобных веществ биохимики используют направленность действия этих веществ на различные холинорецепторы. Если придерживаться такого принципа, то вещества, оказывающие влияние на холинорецепторы, можно классифицировать следующим образом:
- Вещества, которые оказывают влияние на М-холинорецепторы и Н-холинорецепторы: к холиномиметикам относятся ацетилхолин и карбахолин, а к холиноблокаторам – циклодол. характера. К ним относятся салицилат физостигмина, прозерин, гидробромид галантамина, армин.
- Вещества, которые влияют на холинергические синапсы. К холиномиметикам относятся гидрохлорид пилокарпина и ацеклидин, к холиноблокаторам – сульфат атропина, матацин, гидротартрат платифиллина, бромид ипратропия, гидробромид скопаламина.
Читайте также: